


Introduction
Drought, a major consequence of climate change, affects weed growth, invasiveness, and management, posing a major threat to global crop and pasture production (Abbas et al. Reference Abbas, Farooq and Nadeem2025). Use of herbicides is the primary and most efficient method of weed control on a global scale. Glyphosate is the most widely used herbicide due to its broad-spectrum control and relatively low environmental toxicity (Nadeem et al. Reference Nadeem, Abbas, Tanveer, Maqbool, Zohaib and Shehzad2017). Water stress affects the absorption and translocation of glyphosate within the plant by influencing physiological processes, including leaf water status, stomatal conductance, and photosynthesis (Cowie et al. Reference Cowie, Byrne, Witkowski, Strathie, Goodall and Venter2020; Skelton et al. Reference Skelton, Ma and Riechers2016). Water stress also reduces herbicide efficacy by causing changes such as altered leaf orientation, stomatal closure, increased cuticle thickness, and dehydration, all of which limit herbicide absorption and translocation (Abbas et al. Reference Abbas, Farooq and Nadeem2025; Varanasi et al. Reference Varanasi, Prasad and Jugulam2016). Previous studies have revealed reduced efficacy of herbicides, including glyphosate, under water-stress conditions (Alizade et al. Reference Alizade, Keshtkar, Mokhtassi-Bidgoli, Sasanfar and Streibig2021; Bastiani et al. Reference Bastiani, Roma-Burgos, Langaro, Salas-Perez, Rouse, Fipke and Lamego2021; Zhou et al. Reference Zhou, Tao, Messersmith and Nalewaja2007). Enhancing herbicide performance under such conditions has become a significant challenge for modern weed scientists to protect agroecosystems while ensuring profitable crop production. Effective herbicide application is also crucial for delaying the development of herbicide resistance and for minimizing the unintended effects of hormesis on both crops and weeds (Abbas et al. Reference Abbas, Nadeem, Tanveer and Ahmad2016; Nadeem et al. Reference Nadeem, Abbas, Tanveer, Maqbool, Zohaib and Shehzad2017).
Leaf water content (LWC), a vital indicator of plant water status, is key to determining soil moisture stress (Zhou et al. Reference Zhou, Zhou, He, Zhou, Ji and Lv2021). Studies have revealed that LWC is the optimal indicator of soil moisture stress in standing crop conditions, where direct soil moisture assessment is difficult (Zhang and Zhou Reference Zhang and Zhou2019; Zhang et al. Reference Zhang, Xu, She, Hu, Zhu, Wang, Wu, You, Ke, Zhang and He2024). Advanced remote sensing and machine learning technologies, including LiDAR, hyperspectral imaging, microwave remote sensing, and terahertz time-domain spectroscopy, significantly improve LWC assessment, enabling real-time monitoring (Abbas et al. Reference Abbas, Farooq and Nadeem2025; Guo et al. Reference Guo, Feng, Yang and Yang2024; Zhang et al. Reference Zhang, Xu, She, Hu, Zhu, Wang, Wu, You, Ke, Zhang and He2024). Hence, real-time monitoring LWC of weed plants can help to estimate soil water stress and optimize herbicide application accordingly (Zhang et al. Reference Zhang, Xu, She, Hu, Zhu, Wang, Wu, You, Ke, Zhang and He2024).
LWC is a fundamental ecophysiological trait that influences the efficiency of various plant physiological processes (Abbas et al. Reference Abbas, Farooq and Nadeem2025). Reduced LWC under water-stress conditions decreases stomatal conductance and net photosynthesis (Xu et al. Reference Xu, Long, Feng, Zhu, Sai, Chirkova, Betts, Herrmann, Edwards, Okamoto, Hedrich and Gilliham2021). These alterations negatively affect the uptake and translocation of glyphosate by inhibiting its absorption at the leaf surface and its movement within the plant, which primarily relies on phloem transport driven by the plant’s water status (Abbas et al. Reference Abbas, Farooq and Nadeem2025; Skelton et al. Reference Skelton, Ma and Riechers2016). The potential of plants to maintain LWC determines their ability to tolerate drought and serves as a guide for the precise application of inputs, such as irrigation scheduling and nutrient management (Gu et al. Reference Gu, Qi, Burghate, Yuan, Jiao and Xu2020; McGrath and Lobell Reference McGrath and Lobell2013).
Influence of water stress on glyphosate efficacy has been studied for some weed species; however, comparative responses of different weed morphologies such as Santa Maria feverfew (Parthenium hysterophorus L.) (C₃ broadleaf) and giant foxtail (Setaria faberi Herrm.) (C₄ grass) remain limited. Parthenium hysterophorus is a cross-pollinated, troublesome, and climate-resilient invasive weed with significant negative effects on agriculture, environment, and human health globally (Bajwa et al. 2017; Mao et al. Reference Mao, Bajwa and Adkins2021). Its adaptability to tolerate a broad range of temperatures and drought stress, strong allelopathic potential, and rapid reproductive cycle make it particularly difficult to control (Abbas et al. Reference Abbas, Nadeem, Burgos, Matloob, Kamran, Munir, Kalyar and Farooq2024; Cowie et al. Reference Cowie, Byrne, Witkowski, Strathie, Goodall and Venter2020; Sharma et al. Reference Sharma, Rathee, Ahmad, Siddiqui, Alamri, Kaur, Kohli, Singh and Batish2024). Variation in its LWC across different elevations has been reported, underscoring its physiological adaptability (Sharma et al. Reference Sharma, Rathee, Ahmad, Siddiqui, Alamri, Kaur, Kohli, Singh and Batish2024).
Setaria faberi is an annual, self-pollinated invasive weed native to eastern Asia (Nurse et al. Reference Nurse, Darbyshire, Bertin and DiTommaso2009). It has become widely established in temperate, tropical, and subtropical regions, where it causes severe crop yield losses (Nurse et al. Reference Nurse, Darbyshire, Bertin and DiTommaso2009; Papapanagiotou et al. Reference Papapanagiotou, Alvanou, Giantsis, Vasilakoglou and Eleftherohorinos2025). Its rapid evolution of competitive ability and the development of resistance to multiple herbicide modes of action make its control increasingly challenging (Ethridge et al. Reference Ethridge, Chandra, Everman, Jordan, Locke, Owen and Leon2023; Papapanagiotou et al. Reference Papapanagiotou, Alvanou, Giantsis, Vasilakoglou and Eleftherohorinos2025). Understanding how water stress–induced changes in LWC affect glyphosate efficacy across diverse weed types could be a milestone in optimizing site-specific herbicide dose through remote estimation of LWC. Remotely monitoring LWC can help schedule irrigation before herbicide application or optimize site-specific herbicide doses to overcome the detrimental effect of drought stress on glyphosate efficacy (Abbas et al. Reference Abbas, Farooq and Nadeem2025; Gu et al. Reference Gu, Qi, Burghate, Yuan, Jiao and Xu2020). The objective of this study was to evaluate the effect of drought on LWC of two weed species representing different morphologies and to assess the impact of LWC on glyphosate efficacy.
Materials and Methods
Greenhouse Site, Design, and Experimental Setup
The experiments were conducted under greenhouse conditions, with a temperature range of 24–37 C, relative humidity between 50% and 65%, and a photoperiod of 13 to 14 h. A completely randomized design with a factorial arrangement was used, incorporating two treatment factors: moisture levels (well watered at 100% WHC, moderate at 75% WHC, and severe at 50% WHC) and glyphosate doses (0, 180, 360, 540, 720 and 900 g ae ha⁻¹). Here, 720 g ha⁻¹ represents the recommended field rate. To minimize positional effects, pots were rotated after 3 d. The study was conducted in 2025 and was repeated during the same year.
The soil used in this study was a sandy clay loam, collected from the 0- to 12-cm soil depth at the College of Agriculture, University of Sargodha (UOS), Sargodha, Pakistan (32.134°N, 72.686°E). Soil samples were randomly collected from various locations in the field and combined into a composite sample. The soil was air-dried at 70 C for 12 d until completely dry, then sieved to ≤2 mm to remove large-size aggregates, and subsequently used to fill plastic pots. The soil had the following physicochemical properties: sand, 58%; silt, 28%; clay, 14%; pH 7.7; EC, 2.48 dS m−1; total nitrogen, 60 mg kg−1; available phosphorus, 8.35 mg kg−1; total organic carbon, 7 g kg−1; bulk density, 1.58 Mg m−3; and 56% water-holding capacity (WHC). To simulate different water-stress conditions in a controlled environment, soils were maintained at 100% WHC (well watered), 75% WHC (moderate drought stress), and 50% WHC (severe drought stress) (Bajwa et al. 2017; Farooq et al. Reference Farooq, Koirala, Loftus, Zhang, Zamanian, Banfield and Dippold2025). Soil moisture levels were adjusted daily based on gravimetric water content to maintain the target water-stress condition throughout the experimental period.
Seeds of P. hysterophorus and S. faberi were collected from plants growing naturally along the roadside near the College of Agriculture, UOS, Sargodha, Pakistan (32.133°N, 72.687°E). The plants had not been exposed to herbicides before seed collection. The seeds were first kept in the shade for 2 d and then sun-dried for 1 d, after which they were packed in brown paper bags and stored at room temperature in a well-ventilated storage area (with relative humidity below 20%) until use (for approximately 6 mo). Seeds were sown in 12 by 12 cm square pots having a 25-cm depth. Initially, four seeds were sown per pot and later thinned to retain one healthy and vigorous seedling per pot.
Glyphosate was sprayed at 28 d after emergence, when seedlings had reached the 5- to 6-leaf stage. Plants in the control treatment were not sprayed and were allowed to grow until the end of the experiment, 49 d after emergence. Water-stress conditions were checked, and water was applied to maintain each drought level before glyphosate application. Glyphosate doses, as per the treatment plan, were applied using a spray chamber fit with a TeeJet® nozzle (Kissan Ghar, Pakistan) calibrated to deliver 108 L ha−1 of spray volume at 200 kPa. Control pots were sprayed with water only. Pots were not irrigated for 24 h after spraying. Drought treatments were maintained continuously before and after spraying.
Data Collection and Analysis
LWC was determined on the day of spraying. One youngest fully expanded leaf was sampled from each plant. The LWC values were calculated using the following formula (Garnier and Laurent 1994):
 $${\rm{LWC}} = {{{{\rm{LFW}} - {\rm{LDW}}}}\over{{{\rm{LFW}}}}}\; \times 100$$
$${\rm{LWC}} = {{{{\rm{LFW}} - {\rm{LDW}}}}\over{{{\rm{LFW}}}}}\; \times 100$$
where LFW represents the leaf fresh weight measured immediately after removal from plant, and LDW represents the leaf dry weight after oven drying at 70 C until constant weight was achieved.
Stomatal conductance (mol m−2 s−1) was measured on intact leaves with a portable porometer on the day of spraying. Three readings were recorded from each plant and averaged.
The shikimic acid concentration was measured at 7 d after spraying via high-performance liquid chromatography (Cardinali et al. Reference Cardinali, Dias, Mueller, Abercrombi, Stewart, Tornisielo and Christoffoleti2015). Values are expressed as grams of shikimate per kilogram of dry matter (g kg−1). A higher shikimic acid concentration indicates greater glyphosate efficacy.
Weed mortality percentage and biomass reduction were assessed to study treatment effects on glyphosate efficacy. Mortality percentage of weeds was determined at 21 d after glyphosate application. The shoots were harvested at the soil surface at 21 d after glyphosate application, oven-dried at 70 C to a constant weight, and recorded as shoot dry weight (SDW).
Biomass reduction was calculated as follows:
 $${\rm{\% \;Biomass\;reduction}} = {{{{\rm{SDW\;treated}}}}\over{{{\rm{SDW\;control}}}}}{\rm{\;}} \times 100$$
$${\rm{\% \;Biomass\;reduction}} = {{{{\rm{SDW\;treated}}}}\over{{{\rm{SDW\;control}}}}}{\rm{\;}} \times 100$$
Statistical Analysis
The values shown in the tables represent the means of four replicates ± standard errors of the means (mean ± SEM). Data were tested for normality (Shapiro-Wilk test, P > 0.05) and homogeneity of variance (Levene’s test, P > 0.05) before ANOVA and met the assumptions of normality. Data from experiments conducted under a completely randomized design with a factorial arrangement were analyzed separately for each species. For each measured trait, a two-way ANOVA with moisture levels and glyphosate doses as factors was performed using the computer package Statistix v. 8.1 (Analytical Software, Tallahassee, FL, USA) (Steel et al. Reference Steel, Torrie and Dickey1997). Mean values were separated using Tukey’s honest significant difference (HSD) test at α = 0.05. Species sensitivity was assessed by directly comparing the mean values for each trait under identical treatments, as generated from the parallel ANOVAs.
Dose–response curves were generated and analyzed using the drc package in R (v. 4.5.1) (Ritz et al. Reference Ritz, Baty, Streibig and Gerhard2015). A four-parameter log-logistic model was fit to the growth response data at each moisture level (50%, 75%, and 100% WHC). The model equation used was:
 $$f\left( x \right) = c + \left( {d - c} \right)/(1 + \exp \left( {b\left( {\log \left( x \right) - \log \left( e \right)} \right)} \right))$$
$$f\left( x \right) = c + \left( {d - c} \right)/(1 + \exp \left( {b\left( {\log \left( x \right) - \log \left( e \right)} \right)} \right))$$
where f( x ) is the biomass reduction at dose x , d is the upper limit (response at 0 dose), c is the lower limit (response at infinite dose), b is the slope of the curve around the inflection point, e is the effective dose at which the response is halfway. GR50, the dose causing 50% biomass reduction, was derived from model parameters. The relationship of LWC with shikimic acid concentration and mortality % was analyzed using Pearson’s correlation. The correlation coefficient (r) and its statistical significance (P-value) were calculated. Analyses were conducted using R (v. 4.5.1). As there was no significant effect of experimental run (P > 0.05), data from both runs were pooled for all subsequent analyses and graphical presentation.
Results and Discussion
LWC
Water stress caused a significant reduction in the LWC of both P. hysterophorus and S. faberi (Table 1 ). The reduction in LWC was more pronounced under severe drought stress conditions for both weed species. Under well-watered conditions, higher LWC values (ranging from 91% to 95%) were recorded in the broadleaf weed (P. hysterophorus) compared with the narrow-leaf weed (S. faberi), which showed LWC values ranging from 84% to 88%. However, under drought conditions, a greater reduction in LWC was observed in P. hysterophorus, while a comparatively smaller reduction was recorded for S. faberi. The average LWC of P. hysterophorus under well-watered, moderate drought, and severe drought conditions was 93%, 83%, and 68%, respectively. For S. faberi, the average LWC under the same conditions was 86%, 80%, and 66%, respectively. Moderate and severe drought stress caused 10% and 26% reduction in the LWC of P. hysterophorus, respectively. In contrast, the same conditions caused 6% and 23% reduction in the LWC of S. faberi, respectively. Under low soil moisture conditions, water uptake is reduced at the root surface, which ultimately affects leaf water relations (Abbas et al. Reference Abbas, Farooq and Nadeem2025). The ability of plants to maintain LWC varies by species; plants with greater stress tolerance maintain higher LWC under water-deficit conditions and continue physiological processes effectively (Abbas et al. Reference Abbas, Farooq and Nadeem2025; Guerfel et al. Reference Guerfel, Baccouri, Boujnah, Chaïbi and Zarrouk2009). In contrast, sensitive species cannot maintain their LWC under stress, which negatively impacts physiological processes such as photosynthesis, transpiration, and stomatal conductance. The effect of water stress on LWC and physiological processes is well documented for various crop species (Egilla et al. 2005). Weeds like P. hysterophorus also exhibit reductions in leaf water potential and relative water content under water-stress conditions (Cowie et al. Reference Cowie, Byrne, Witkowski, Strathie, Goodall and Venter2020). The differential ability to maintain LWC observed between P. hysterophorus and S. faberi is attributed to their distinct morphologies. Narrow-leaved plants demonstrate greater drought tolerance by maintaining higher LWC under water-stress conditions (Farooq et al. Reference Farooq, Wahid, Kobayashi, Fujita and Basra2009).
Effects of water-stress levels on the leaf water content (LWC) of Parthenium hysterophorus and Setaria faberi a .

a Water-stress levels: well-watered, 100% water holding capacity (WHC); moderate drought, 75% WHC; and severe drought, 50% WHC. Values are presented as mean ± SE (n = 4). Within a column for each species, means with different letters are significantly different according to Tukey’s honest significant difference (HSD) test (P ≤ 0.05).
Stomatal Conductance
Water stress caused a significant reduction in the stomatal conductance of both P. hysterophorus and S. faberi (Table 2 ). The reduction was more pronounced under severe drought stress conditions for both weed species. Under well-watered conditions, higher stomatal conductance values (ranging from 0.35 to 0.41 mol m−2 s−1) were recorded in P. hysterophorus compared with S. faberi, which showed values ranging from 0.32 to 0.38 mol m−2 s−1. The average stomatal conductance of P. hysterophorus under well-watered, moderate drought, and severe drought conditions was 0.38, 0.22, and 0.07 mol m−2 s−1, respectively. For S. faberi, the average stomatal conductance under the same conditions was 0.35, 0.21, and 0.06 mol m−2 s−1, respectively. Moderate and severe drought stress caused 42% and 82% reduction in the stomatal conductance of P. hysterophorus, respectively. In contrast, the same conditions caused 40% and 83% reduction in the stomatal conductance of S. faberi, respectively. As a defensive strategy to survive under drought by maintaining high LWC, plants close their stomata to reduce water losses through transpiration (Guerfel et al. Reference Guerfel, Baccouri, Boujnah, Chaïbi and Zarrouk2009). Increase in abscisic acid concentration and hydraulic changes due to reduced LWC caused reduction in stomatal conductance (Xu et al. Reference Xu, Long, Feng, Zhu, Sai, Chirkova, Betts, Herrmann, Edwards, Okamoto, Hedrich and Gilliham2021). Cowie et al. (Reference Cowie, Byrne, Witkowski, Strathie, Goodall and Venter2020), reported deceased stomatal conductance in P. hysterophorus under water-stress conditions.
Effects of water-stress levels on the stomatal conductance of Parthenium hysterophorus and Setaria faberi a .

a Water-stress levels: well-watered, 100% water holding capacity (WHC); moderate drought, 75% WHC; and severe drought, 50% WHC. Values are presented as mean ± SE (n = 4). Within a column for each species, means with different letters are significantly different according to Tukey’s honest significant difference (HSD) test (P ≤ 0.05).
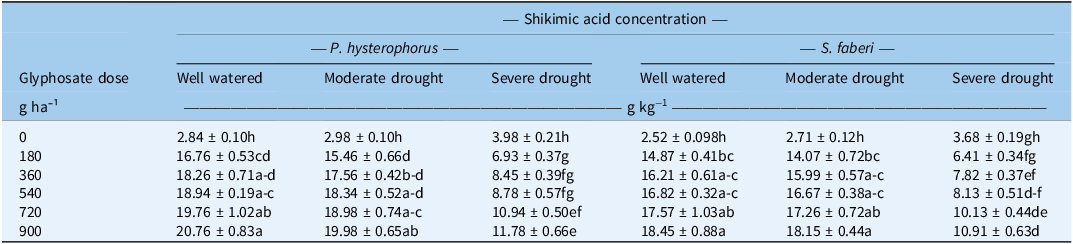
Shikimic Acid Concentration
To estimate glyphosate efficacy, shikimic acid concentration was measured as a proxy for 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) enzyme inhibition. The data revealed that both water-stress levels and glyphosate doses significantly affected the shikimic acid concentration in the leaves of both P. hysterophorus and S. faberi (Table 3 ). The highest shikimic acid concentration was observed when glyphosate was applied at the 900 g ha−1 dose under well-watered conditions for both weed species. Water stress reduced this concentration, with the lowest concentrations found under severe drought stress. For P. hysterophorus, moderate drought stress caused a relatively small reduction in shikimic acid concentration, ranging from 4% to 8% across the five glyphosate doses. The reduction was far more pronounced under severe drought, which caused a massive decrease ranging from 43% to 59%. Similarly, for S. faberi, moderate drought stress resulted in a minor reduction (0.9% to 5.4%), while severe drought caused a substantial reduction ranging from 41% to 57%. Notably, the shikimic acid concentration at the 180 g ha−1 glyphosate dose under well-watered conditions was higher than that at the 900 g ha−1 dose under severe drought stress for both species. Hence, severe drought stress significantly inhibited glyphosate efficacy, as measured by the reduced concentration of shikimic acid, across all tested doses. In contrast, moderate drought had little to no effect. A strong positive correlation was observed between LWC (at the time of glyphosate spray) and shikimic acid concentration (at 7 d after glyphosate spray) in both P. hysterophorus and S. faberi (Pearson’s r = +0.846 and 0.828, P = 7.05 × 10−5 and 1.39 × 10−4) (Figure 1). This relationship was consistent across all glyphosate doses tested (180 to 900 g ha−1). The decreased glyphosate efficacy under water-stress conditions is likely due to reduced foliar uptake and translocation, linked to lower LWC and stomatal conductance (Bastiani et al. Reference Bastiani, Roma-Burgos, Langaro, Salas-Perez, Rouse, Fipke and Lamego2021). Studies revealed that LWC impairs key physiological processes in plants (Abbas et al. Reference Abbas, Farooq and Nadeem2025). Specifically, the decreased stomatal conductance inhibits the foliar absorption of glyphosate at leaf surface (Cowie et al. Reference Cowie, Byrne, Witkowski, Strathie, Goodall and Venter2020; Skelton et al. Reference Skelton, Ma and Riechers2016). These outcomes are supported by previous studies that revealed reduced glyphosate absorption under water-stress conditions (Zhou et al. Reference Zhou, Tao, Messersmith and Nalewaja2007). Reduced glyphosate efficacy under water stress (at 50% WHC) compared with well-watered conditions (100% WHC) has been confirmed in South African lovegrass (Eragrostis plana Nees) (Bastiani et al. Reference Bastiani, Roma-Burgos, Langaro, Salas-Perez, Rouse, Fipke and Lamego2021).
Effects of water-stress levels and glyphosate doses on the shikimic acid concentration in Parthenium hysterophorus and Setaria faberi a .
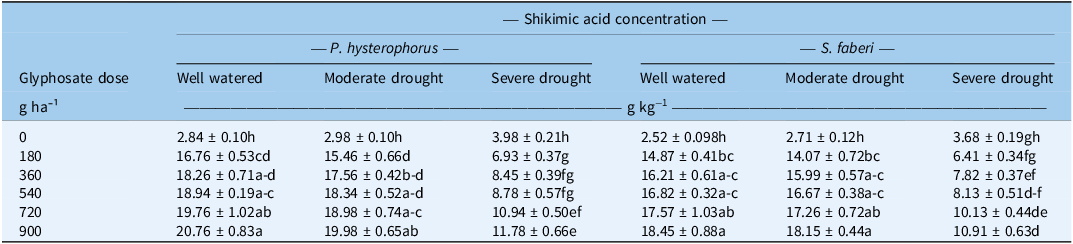
a Water-stress levels: well-watered, 100% water holding capacity (WHC); moderate drought, 75% WHC; and severe drought, 50% WHC. Values are presented as mean ± SE (n = 4). Within a column for each species, means with different letters are significantly different according to Tukey’s honest significant difference (HSD) test (P ≤ 0.05).
Correlation between leaf water content (LWC) and shikimic acid concentration in Parthenium hysterophorus (left) and Setaria faberi (right) across five herbicide doses. Each point present means of observation at each glyphosate dose across three moisture levels. Regression lines show a significant positive correlation (r = 0.846 and 0.828, P = 7.05 × 10−5 and 1.39 × 10−4).

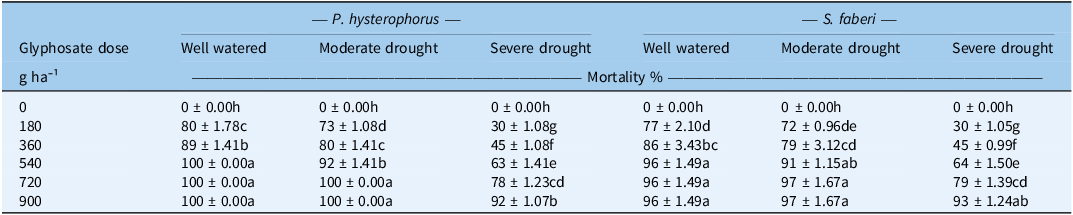
Mortality
The mortality percentage was significantly affected by water-stress levels and glyphosate doses for both weed species, P. hysterophorus and S. faberi (Table 4). The highest mortality (100%) for P. hysterophorus was observed at the 720 and 900 g ha−1 glyphosate doses under both well-watered and moderate drought stress conditions. In contrast, S. faberi showed 96% to 97% mortality at the same doses (720 and 900 g ha−1) under well-watered and moderate drought stress conditions. Water stress reduced the mortality percentage, with the lowest mortality found under severe drought stress for both species. For P. hysterophorus, moderate drought stress caused a relatively small reduction in mortality (0% to 10%). The reduction was far more pronounced under severe drought, which caused a substantial decrease ranging from 8% to 63%, depending on the glyphosate dose. Similarly, for S. faberi, moderate drought stress resulted in a minor reduction (1% to 8%), while severe drought caused a more significant reduction in mortality, ranging from 3% to 61%. For the 720 g ha−1 glyphosate dose, severe drought stress caused 22% and 18% reduction in the mortality of P. hysterophorus and S. faberi, respectively. Notably, the mortality at the 360 g ha−1 glyphosate dose under well-watered conditions was statistically similar to the mortality caused by the 900 g ha−1 dose under severe drought stress for both species. Hence, severe drought stress significantly reduced mortality across all tested doses. In contrast, moderate drought had little to no effect. A strong positive correlation was observed between LWC (at the time of glyphosate spray) and mortality % (at 21 d after glyphosate spray) in both P. hysterophorus and S. faberi (Pearson’s r = +0.59 and 0.53, P = 0.02 and 0.043) (Figure 2). This relationship was consistent across all glyphosate doses tested (180 to 900 g ha−1). The reduction in mortality can be explained by reduced shikimic acid concentration under water-stress conditions. This is attributed to decreased LWC and stomatal conductance, which inhibited glyphosate uptake. A strong positive correlation between LWC and mortality rate has been confirmed in the present study. This finding is consistent with previous studies that reported reduced glyphosate efficacy under water stress (Bastiani et al. Reference Bastiani, Roma-Burgos, Langaro, Salas-Perez, Rouse, Fipke and Lamego2021; Skelton et al. Reference Skelton, Ma and Riechers2016; Zhou et al. Reference Zhou, Tao, Messersmith and Nalewaja2007). Similarly, glyphosate efficacy for controlling P. hysterophorus was reduced under stress conditions imposed by elevated carbon dioxide concentration (Bajwa et al. Reference Bajwa, Wang, Chauhan and Adkins2019).
Effects of water-stress levels and glyphosate doses on the mortality % of Parthenium hysterophorus and Setaria faberi a .

a Water-stress levels: well-watered, 100% water holding capacity (WHC); moderate drought, 75% WHC; and severe drought, 50% WHC. Values are presented as mean ± SE (n = 4). Within a column for each species, means with different letters are significantly different according to Tukey’s honest significant difference (HSD) test (P ≤ 0.05).
Positive correlation between leaf water content (LWC) and mortality % in Parthenium hysterophorus (left) and Setaria faberi (right) across five herbicide doses. Each point present means of observation at each glyphosate dose across three moisture levels. Regression lines show a significant positive correlation (r = 0.59 and 0.529, P = 2.0 × 10⁻² and 4.3 × 10⁻²).

Biomass Reduction
The reduction in dry biomass was significantly affected by the different levels of drought stress and glyphosate doses for both weed species, P. hysterophorus and S. faberi (Table 5 ). The highest biomass reduction (84% to 91%) for P. hysterophorus was observed at the 720 and 900 g ha−1 glyphosate doses under both well-watered and moderate drought stress conditions. In contrast, S. faberi showed 78% to 89% biomass reduction at the same doses (720 and 900 g ha−1) under well-watered and moderate drought stress conditions. Water stress reduced the biomass reduction, with the lowest biomass reduction found under severe drought stress for both species. For P. hysterophorus, moderate drought stress caused a relatively small reduction in biomass (4% to 8%). The reduction was far more pronounced under severe drought, which caused a substantial decrease ranging from 29% to 58%, depending on the glyphosate dose. Similarly, for S. faberi, under moderate drought stress, 3% to 7% less biomass reduction occurred compared with well-watered conditions, while a 26% to 57% lower reduction in biomass occurred under severe drought conditions. Notably, the biomass reduction at the 180 g ha−1 glyphosate dose (62%) under well-watered conditions was statistically similar to the biomass reduction (65%) caused by the 900 g ha−1 dose under severe drought stress for P. hysterophorus. Similarly, the biomass reduction at the 360 g ha−1 glyphosate dose (68%) under well-watered conditions was statistically similar to the biomass reduction (66%) caused by the 900 g ha−1 dose under severe drought stress for S. faberi. Hence, severe drought stress caused significant inhibition of biomass reduction across all tested doses. The lower biomass reduction observed under water-stress conditions can be explained by reduced shikimic acid concentration, which reflected decreased inhibition of EPSPS enzyme, a key catalyst in the shikimate pathway. Consequently, less EPSPS inhibition allowed the pathway to remain active, enabling the weeds to sustain growth. Reduced efficacy of glyphosate under water-stress conditions is supported by the work of Zhou et al. (Reference Zhou, Tao, Messersmith and Nalewaja2007) and Skelton et al. (Reference Skelton, Ma and Riechers2016). Similarly, glyphosate application at 50% WHC showed less efficacy to reduce biomass of E. plana compared with its application at 100% WHC (Bastiani et al. Reference Bastiani, Roma-Burgos, Langaro, Salas-Perez, Rouse, Fipke and Lamego2021).
Effects of water-stress levels and glyphosate doses on the biomass reduction of Parthenium hysterophorus and Setaria faberi a .

a Water-stress levels: well watered, 100% water holding capacity (WHC); moderate drought, 75% WHC; and severe drought, 50% WHC. Values are presented as mean ± SE (n = 4). Within a column for each species, means with different letters are significantly different according to Tukey’s honest significant difference (HSD) test (P ≤ 0.05).
The dose–response curves of glyphosate efficacy in reducing biomass of P. hysterophorus and S. faberi were significantly influenced by different levels of drought stress (Figures 3 and 4). Increasing drought stress led to higher GR50, indicating that water stress reduced glyphosate efficacy. For P. hysterophorus, the GR50 values were 115, 128 and 498 g ha−1 at 100%, 75%, and 50% WHC, respectively. Similarly, for S. faberi, the GR50 values were 124, 135 and 481 g ha−1 at the same drought stress level. Results showed a clear inverse relationship between glyphosate efficacy and drought stress. An approximately 4-fold higher GR50 (481 to 498 g ha−1) was required under severe drought (50% WHC) compared with GR50 (115 to 124 g ha−1) under well-watered (100% WHC) conditions.
Effect of water-stress levels (well watered, 100% water holding capacity [WHC]; moderate drought, 75% WHC; and severe drought, 50% WHC); and glyphosate doses on dry biomass reduction of Parthenium hysterophorus, expressed as percentage of means (n = 4).

Effect of water-stress levels (well watered, 100% water holding capacity [WHC]; moderate drought, 75% WHC; and severe drought, 50% WHC) and glyphosate doses on dry biomass reduction of Setaria faberi, expressed as percentage of means (n = 4).

Effective weed control under changing climate conditions demands novel, climate-responsive strategies to ensure the precise application of herbicides (Abbas et al. Reference Abbas, Farooq and Nadeem2025). The results of the present study revealed that increasing water stress significantly reduced LWC and stomatal conductance in two weeds exhibiting different morphologies: broadleaf (P. hysterophorus) and narrow-leaf (S. faberi). The broadleaf (P. hysterophorus) showed comparatively more reduction in LWC and stomatal conductance compared with the narrow-leaf (S. faberi). These physiological changes caused reduction in shikimic acid concentration, showing strong correlation with the change in LWC. Consequently, the reduced efficacy of glyphosate (less mortality and biomass reduction) in correlation to reduction in LWC was observed. Our data show that LWC and stomatal conductance were both reduced under water stress. The correlation of LWC with shikimic acid concentration and glyphosate efficacy suggest that LWC can serve as a valuable, integrative biomarker for predicting herbicide efficacy. Results suggest that maintaining LWC above approximately 75% is critical for ensuring effective glyphosate efficacy. Glyphosate applications when LWC falls below 70% may result in unacceptable levels of weed control at recommended field rates. To the best of our knowledge, the potential of using LWC as a predictive biomarker for herbicide efficacy under water stress has not been explored. However, LWC is already established as a strong indicator of available soil water content in crop production (Zhou et al. Reference Zhou, Zhou, He, Zhou, Ji and Lv2021) and intensively used for irrigation scheduling, nutrient application, and other inputs (Abbas et al. Reference Abbas, Farooq and Nadeem2025; Gu et al. Reference Gu, Qi, Burghate, Yuan, Jiao and Xu2020). In addition, weed species exhibited different magnitudes of LWC reduction under drought; however, a nearly identical proportional loss in glyphosate efficacy for both weed species indicates that the physiological water status itself (LWC), rather than morphological distinction, is the primary factor of herbicide performance under drought. This highlights the potential importance of LWC as predictive biomarker for herbicide efficacy across diverse weed types.
Using LWC as indicator of water stress and herbicide efficacy can significantly help to improve weed management under water-stress conditions, as measuring soil water content in a standing crop is practically difficult and less efficient. Advanced remote sensing and machine learning techniques can remotely estimate real-time LWC (Abbas et al. Reference Abbas, Farooq and Nadeem2025; Guo et al. Reference Guo, Feng, Yang and Yang2024). The outcome of the present study can be used to estimate soil water stress and anticipate glyphosate efficacy by remotely measuring LWC of weeds before glyphosate application. Growers can schedule irrigation before herbicide application and/or optimize real-time herbicide doses based on LWC information for effective weed control. This study proposes LWC as a vital physiological trait to guide herbicide use that has been overlooked in previous weed management studies. Validating the relationship between LWC and herbicide efficacy across other weed species and herbicides is suggested in future studies to develop precision weed management protocols capable of overcoming the challenges of climate change. Furthermore, future research should investigate how quickly LWC recovers and glyphosate susceptibility is restored following irrigation of drought-stressed weeds. This information will be vital for optimizing the timing of both irrigation and subsequent herbicide application in a precision weed management system.
Acknowledgments
The authors would like to thank Faisal Rashid, PhD agronomy student at the College of Agriculture, University of Sargodha, Sargodha, Pakistan, for his dedicated work measuring LWC and dry biomass.
Funding statement
This research received no specific grant from any funding agency or the commercial or not-for-profit sectors.
Competing interests
No competing interests have been declared by the authors.








 Open access
Open access