


Non-technical Summary
Jellyfish and their polyp relatives belong to an ancient group of animals called cnidarians. Because their bodies are soft, they almost never fossilize, leaving gaps in our understanding of their origins. We describe a new fossil species, Paleocanna tentaculum, from rocks approximately 450 million years old (Upper Ordovician) in Québec, Canada. These remains are preserved as thin, carbon-rich impressions in limestone, alongside other typical shelly marine species. This cnidarian species lived inside upright tubes, alone or in clusters. Some tubes show faint striations near their bases. The polyp itself was long and narrow, with a ring of tentacles that extended above the rim of the tube. By comparing Paleocanna with other living and fossil genera, we found that it is more closely related to modern jellyfish groups such as box jellies, true jellyfish, and stalked jellyfish than it is to other extinct tube-dwelling relatives. This places our species nearer to the modern branch of the family tree than most other known fossil polyps. The exceptional preservation of these fossils makes this discovery one of the rare cases in which delicate soft-bodied organisms are found in Ordovician rocks.
Introduction
The Neoproterozoic era marks the origin of the cnidarian crown group, possibly in the Ediacaran or Cryogenian period (Van Iten et al., Reference Van Iten, Marques, Leme, Pacheco and Simoes2014). Polypoid cnidarians are predominantly known from their mineralized representatives, e.g., rugose and tabulate corals, which exhibit well-documented skeletal morphologies. In contrast, tubicolous cnidarians with soft-bodied preservation remain relatively rare despite their potential to shed light on the early evolution within the phylum.
Among the earliest representatives are Paraconularia ediacara Leme, Van Iten, and Simões, Reference Leme, Van Iten and Simões2022 from the Ediacaran Tamengo Formation of Brazil, considered the oldest documented conulariid and an example of early cnidarian skeletonization. Other carinachitids and hexangulaconulariids, e.g., Hexaconularia He and Yang, Reference He and Yang1986 and Carinachites Qian, Reference Qian1977 from South China, represent some of the earliest early Cambrian polypoid tubicolous cnidarians, interpreted as members of the stem group Medusozoa (Van Iten et al., Reference Van Iten, Zhu and Li2010; Song et al., Reference Song, Guo, Han, Van Iten and Peng2024). Auroralumina attenboroughii Dunn et al., Reference Dunn, Kenchington, Parry, Clark, Kendall and Wilby2022 from the Charnwood Forest, UK, is interpreted as a putative tubicolous stem-group medusozoan, with a bifurcating tubular structure and predatory behavior. Haootia quadriformis Liu et al., Reference Liu, Matthews, Menon, McIlroy and Brasier2014 from the Ediacaran Fermeuse Formation of Newfoundland was assigned as a total-group cnidarian and a crown-group staurozoan (McIlroy et al., Reference McIlroy, Pasinetti, Pérez-Pinedo, McKean, Dufour, Matthews, Menon, Nicholls and Taylor2024). These fossils from late Ediacaran strata support the idea that cnidarians diversified before the Cambrian.
Fossil records of cnidarians are rare and controversial for the Cambrian Period, most of which come from the Chengjiang biota (Liu et al., Reference Liu, Matthews, Menon, McIlroy and Brasier2014; Van Iten et al., Reference Van Iten, Marques, Leme, Pacheco and Simoes2014; Walde et al., Reference Walde, Weber, Erdtmann and Steiner2019; Dunn et al., Reference Dunn, Kenchington, Parry, Clark, Kendall and Wilby2022; Zhang et al., Reference Zhang, Parry, Vinther and Ma2022). Among these taxa, Archisaccophyllia kunmingensis Hou et al., Reference Hou, Stanley, Zhao and Ma2005 might instead have a closer affinity to phoronids, whereas Xianguangia sinica Chen and Erdtmann, Reference Chen, Erdtmann, Simonetta and Morris1991 could be a cnidarian or a stem-group ctenophore, implying a sea anemone-like ancestor for ctenophores (Ou et al., Reference Ou, Shu, Zhang, Han and Van Iten2022; Zhao et al., Reference Zhao, Vinther, Parry, Wei, Green, Pisani, Hou, Edgecombe and Cong2019). Conicula striata Luo et al., Reference Luo, Hu, Chen, Zhang and Tao1999 has variously been classified as a lophophorate (Luo et al., Reference Luo, Hu, Chen, Zhang and Tao1999; Zeng et al., Reference Zeng, Chen, Liu, Zhu, Zhao and Yang2022), an anthozoan (Hu, Reference Hu2005), and most recently as a stem-group medusozoan (Zhao et al., Reference Zhao, Parry, Vinther, Dunn, Li, Wei, Hou and Cong2023). The anthozoan hypothesis is based on a morphology that includes a polyp with an actinopharynx, characteristic of anthozoans, and enclosed within a conical, annulated periderm, suggesting that these traits were present in medusozoans and anthozoans. The Chengjiang hyolithid Burithes Missarzhevskii, Reference Missarzhevskii1969 is now reclassified as a stem-group medusozoan, Palaeoconotuba Qu, Li, and Ou, Reference Qu, Li and Ou2023. This reinterpretation was based on the presence of a conical theca and a funnel-like gastrovascular cavity. These two species in Palaeoconotuba and Conicula represent tubicolous forms, a morphological adaptation that appears to have been common in the Cambrian.
Other early Cambrian fossils provide evidence of cnidarian soft-bodied polyps and thecate species. Nailiana elegans Ou et al., Reference Ou, Shu, Zhang, Han and Van Iten2022, from the Chengjiang biota, is an early anthozoan. Olivooides Jian et al., Reference Jian, Guoxiang, Shin, Qiang and Sho2016, from the early Cambrian of China, represents a possible stem-group cubomedusa because it exhibits a conical peridermal theca and features characteristics of polyp and medusa stages. Gangtoucunia aspera Zhang et al., Reference Zhang, Parry, Vinther and Ma2022, from the Wulongqing Formation, represents a total-group medusozoan like a modern scyphozoan polyp. This benthic polyp has a long, tubular, annulated theca that is a mineralized tube, reinforcing the idea that cnidarian biomineralization evolved early in the group’s history.
In the context of previously described stem-group medusozoans, this study presents the first detailed description of an Ordovician, soft-bodied, tubicolous-polyp medusozoan. This discovery is particularly significant given the scarcity of nonbiomineralized organisms in the Ordovician and underscores the crucial role of exceptional soft-tissue preservation in reconstructing early cnidarian evolution.
Locality and geological setting
The fossil materials, preserved along the upper surfaces of shaly limestone beds, were collected from a small quarry located at Saint-Joachim, ~ 43 km northeast of Québec City, Québec, Canada, considered herein within the Upper (Grondines Member) Neuville Formation of the Trenton Group (Harper and Pickerill, Reference Harper and Pickerill1996). The Trenton Group of Québec represents a transgressive continental (Laurentian), platformal margin succession of Katian age, deposited on a carbonate ramp that primarily developed in warm-water tropical and later in temperate conditions (Lavoie, Reference Lavoie1995; Desrochers et al., Reference Desrochers, Jisuo, Keith, Servais, Harper, Lefebvre and Pervical2023). The Upper Neuville Formation type locality near the town of Neuville, ~ 80 km southwest of Saint-Joachim, underlies the Utica Shale and consists of 98 m of beds of dark gray limestones up to 25 cm thick, separated by shale interbeds up to 15 cm thick (Clark and Globensky, Reference Clark and Globensky1973).
The associated biota includes rugose corals, conulariids, bryozoans, brachiopods, gastropods, cephalopods, trilobites, crinoids, carpoids, cystoids, and trace fossils.
Materials and methods
All material observed is curated in the Musée de Paléontologie et de l’Évolution in Montréal, Québec. Approximately 135 specimens preserved on 15 slabs were studied. Thirty-nine specimens were sufficiently complete and visible to be measured and photographed. Photos were taken using (1) soft-white halogen lightbulbs; (2) cross-polarized light with an optical filter enhancing contrast to reveal digestive features; and (3) submerged in water or ethanol to expose details not discernible through other methods. The specimens, being flat in relief, required no preparation before photography.
Elemental mapping (EDS) was conducted on Paleocanna tentaculum n. gen. n. sp., specimen MPEP713.3 using an Environmental Scanning Electron Microscope (ESEM) with a Field Emission Gun (FEG) Quanta 200 model at the University of Windsor Great Lakes Institute for Environmental Research, Ontario, Canada. The EDS mapping and point-elemental analyses were conducted in a low vacuum pressure chamber of 70 Pa. The EDS detector employed was an energy dispersive X-ray spectroscopic (EDAX) Octane Plus SDD detector, operated using TEAM software (https://www.coherent.com.au/content/media/Edax/TEAM%20EDS%20For%20TEM.pdf).
A phylogenetic analysis was performed using 69 taxa, including ctenophore species, and 236 discrete morphological traits. The morphological character traits used were modified from Zhao et al. (Reference Zhao, Parry, Vinther, Dunn, Li, Wei, Hou and Cong2023), especially those concerning the external periderm (see Data Availability Statement). Analysis was run using PAUP 4.0b10 (Swofford, Reference Swofford2002) under the assumptions of parsimony using the heuristic and branch-and-bound search algorithm by bootstrapping using 1,000 replicates. Characters were treated as unordered and with equal weight, unknown states were scored as ‘?’. Changes of individual characters along branches were tracked using Mesquite 3.51 (Maddison and Maddison, Reference Maddison and Maddison2023).
Repository and institutional abbreviation
All specimens are deposited in the Musée de Paléontologie et de l’Évolution (MPEP) collections.
Systematic paleontology
Phylum Cnidaria Verrill, Reference Verrill1865
Subphylum Medusozoa Petersen, Reference Petersen, Larwood and Rosen1979
Genus Paleocanna new genus
Type species
Paleocanna tentaculum n. gen. n. sp., by monotypy.
Diagnosis
As for the type species.
Etymology
Paleocanna: palaios, meaning old or ancient, plus canna, meaning flute or pipe.
Remarks
As for the type species.
Paleocanna tentaculum new species
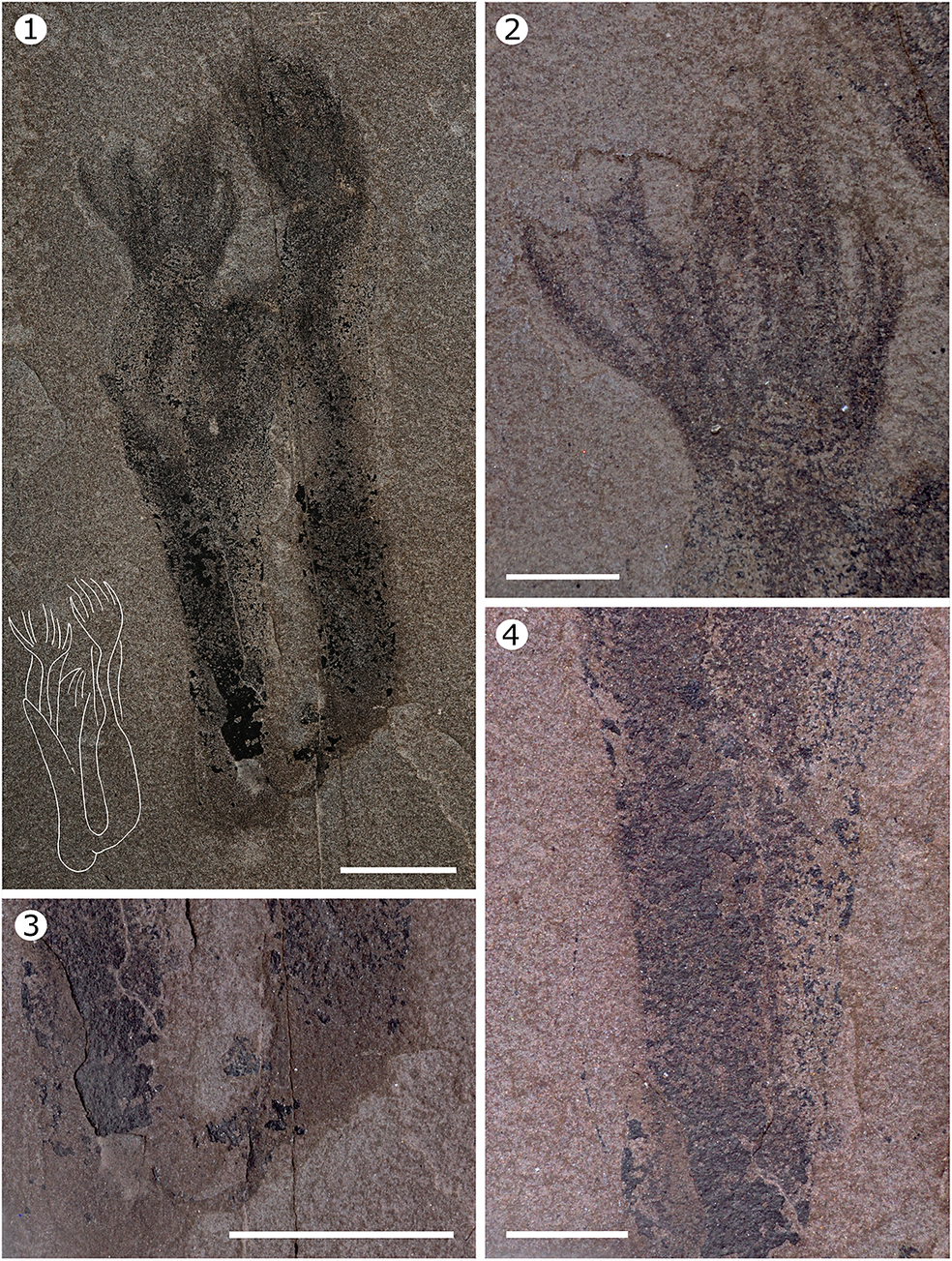
Paleocanna tentaculum n. gen. n. sp., holotype, MPEP713.4. (1) General view of the organism taken under cross-polarized light; accompanying line drawing illustrates the outline of the tube and tentacles. (2) Detail of the left set of tentacles. (3) Detail of the basal part of the tube. (4) Detail of the tube. Scale bars = 1 cm (1, 3), 0.5 cm (2, 4).

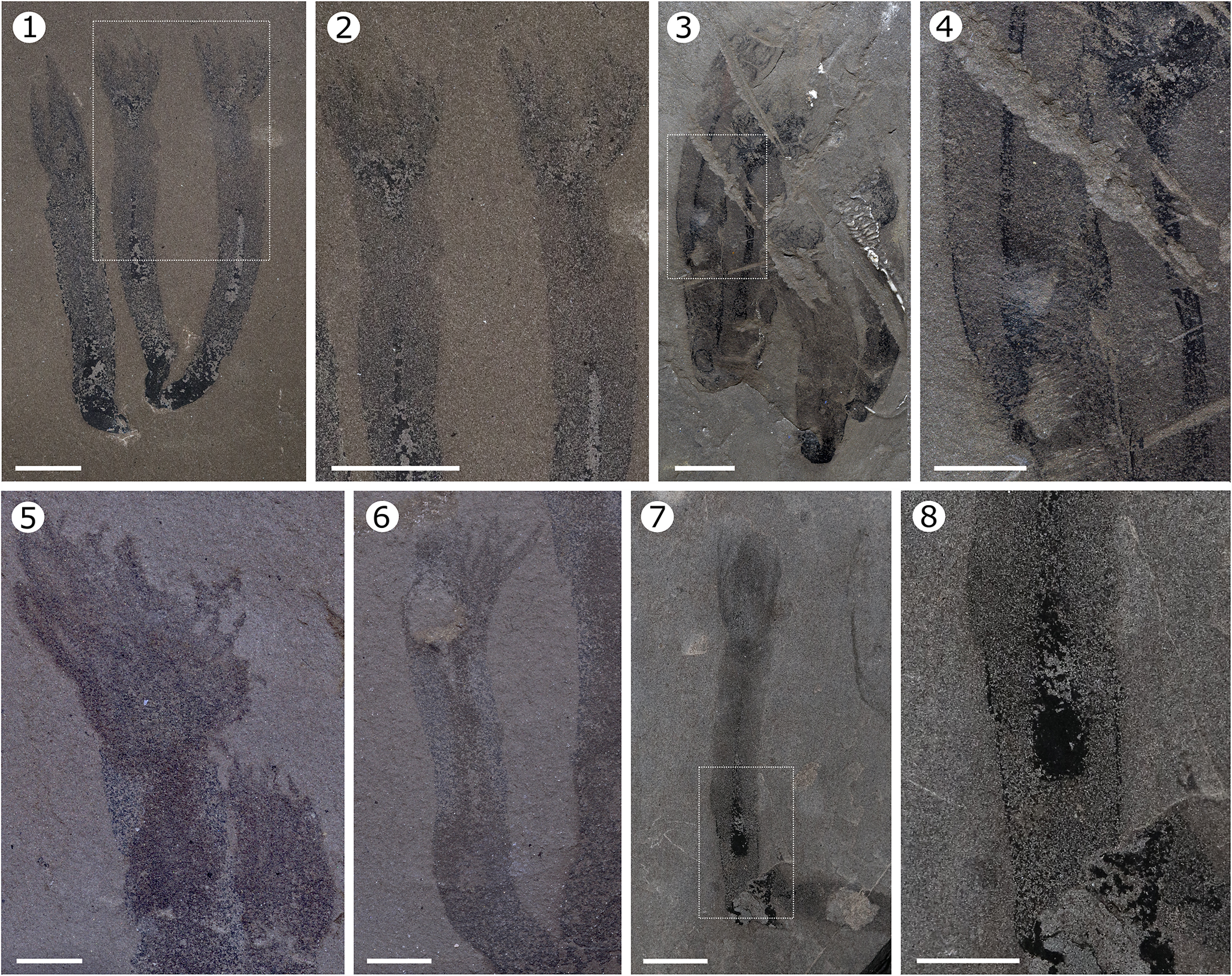
Internal and external anatomy of Paleoacanna tentaculum n. gen. n. sp. (1) MPEP713.6 observed under polarized light showing details of the tentacles and gastrovascular cavity. (2) Detail of the squared region in (1), showing the continuity of the tentacles with the coelenteron. (3) MPEP1524.1 observed immersed in ethanol resembling the morphology of a hydrozoan, illustrating the tentacles, the digestive tract, and the rounded terminus of the periderm. (4) Detail of the squared region in (3), showing the gut and the periderm in greater detail. (5) MPEP713.11 observed under ethanol exhibiting an array of several tentacles. (6) MPEP713.11 showing detail of the tentacles and gut; the tentacles appear finger-like, whereas in other samples, they present a more feather-like form. (7) MPEP713.1 observed under polarized light displayig a sharper tube base, a rarely noted feature that likely helped anchor the organism to the substratum or bury it in sediment. (8) Detail of the squared region in (7), highlighting a central gut, indicating that, as in the other specimens, the gastrovascular cavity does not extend to the distal portion of the tube; a fragment of periderm is preserved at the base. Scale bars = 1 cm (1–4), 0.5 cm (5–8).

Paleocanna tentaculum n. gen. n. sp., living in clusters or individually. (1) MPEP713.6, containing clustered and individual specimens oriented in the same direction, suggesting rapid burial; attached individuals indicate a facultative semicolonial existence. (2) MPEP1525.1 showing a cluster of > 15 individuals; the boxed region (enlarged at upper right) highlighting part of the tubes that exhibit striations. (3) MPEP713.7 showing two clusters of two and four individuals; the former showing some wrinkle-like striations at the base. (4) MPEP713.8 showing one and two individuals, preserved oriented in the same direction. Scale bars = 5 cm (1), 1 cm (2–4).

Periderm annulations of Paleocanna tentaculum n. gen. n. sp. Specimens MPEP713.7 (1, 3) and MPEP713.15 (2, 4) exhibiting different striations that occur parallel to each other in the most distal regions. These markings range from pronounced, wrinkle-like ridges to finer, straight lineations. Scale bars = 0.5 cm.

Type specimens
Holotype: MPEP713.4. Paratypes: MPEP713.1–713.3, 713.5–713.15. Approximately 135 individuals are distributed on these 15 rock slabs.
Diagnosis
Cylindrical tubicolous periderm with fine, parallel, transverse annulations; polyp with anterior tentacle crown extending from aperture.
Occurrence
Upper Neuville Formation of the Saint Lawrence Lowlands of Québec, Canada (Clark, Reference Clark1959).
Description
Paleocanna tentaculum n. gen. n. sp. is a long, uniform, tubicolous organism. Individuals have a mean length of 36.9 mm (ranging from 24.9–51.0 mm) and a mean width of 6.0 mm (N = 39). Some tubes are attached in groups at shared bases, whereas others grew as isolated individuals (Fig. 3). The outer surfaces of the tubes have no sand, broken shells, or other debris. The basal ends are dark black, and some specimens show some regular transverse striations at the base and along the organism (Fig. 4). These annulations appear to be continuous and arranged in parallel. No longitudinal grooves or midlines were observed. The base of the tubes is usually rounded, rarely sharp, and there is no clear evidence of a holdfast. The apical aperture of the tube, from which the animal emerged, ends horizontally and rarely in a sharp angle.
The soft body of the animal is elongated with a rounded aboral end. In all 135 fossils, the body extends from the aperture of the tube. Body septa were not found. Some individuals show a distinct gastrovascular cavity or coelenteron in the center of the tube that terminates blindly (Fig. 2.1, 2.3, 2.7). The midgut is ~ 1.388 mm (+ 4 mm) wide, and the posterior gut is 2.309 mm (+ 3 mm) wide. The tentacles are numerous, ~ 12 in the best-preserved specimens (Fig. 2), without clear evidence of their distribution in a single or multiple coronae. Three specimens show subtle details of the tentacles, which appear as finger-like to feathery structures and represent ~ 20–25% of the total body length (Fig. 2.2, 2.5, 2.6). No individual circlet of tentacles was found in the open, relaxed, feeding position. Instead, the circlet of tentacles is tightly packed, intertwined, and extending anteriorly from the body. No mouth was observed but it was presumed to be situated at the center of the circlet of tentacles. An interpretive drawing of Paleocanna tentaculum n. gen. n. sp. as a living animal is provided (Fig. 5).
Interpretive drawings of Paleocanna tentaculum n. gen. n. sp. as living organisms. (1) Oral view showing a circle of tentacles. (2) Lateral perspective of the tube, highlighting the tube aperture and annular striations. (3) Depiction of individuals living in single tubes, as well as clusters of two or three tubes attached together.

Etymology
Species tentaculum, for tentacle.
Taphonomy
When multiple individuals are present, they are consistently oriented in the same direction, suggesting limited transport and rapid burial (Fig. 3.1, 3.3). Fossils were confined to the upper portions of the beds and did not appear in underlying or overlying strata, supporting rapid deposition events and minimal postburial disturbance. Although some specimens are well preserved (Figs. 1, 2.1, 2.3), most show only faint outlines. This faint preservation of certain structures, e.g., tentacles, could indicate limited decay prior to burial; however, the lack of disarticulation or fragmentation suggests minimal bioturbation or transport after death. This interpretation is supported by the associated taxa, notably echinoderms, which are often preserved as fully articulated specimens. Preservation likely occurred under anoxic to dysoxic conditions, inhibiting decay or scavenging.
Elemental analysis revealed that the darkest areas of the fossils are preserved primarily as carbon films, suggesting retention of original organic material. Other parts are slightly enriched in calcium (6%) and magnesium (5%), which could indicate localized mineralization, possibly reflecting differences in tissue composition or diagenetic overprinting. The surrounding matrix was composed primarily of aluminosilicates with dominant elements including silicon (26%) and aluminum (13%), consistent with a fine-grained siliciclastic sedimentary environment (Fig. 6). No sections of the fossil beds were made, but in situ observations were used to assess fossil occurrence and stratigraphic context.
Elemental analysis of specimen MPEP 713.3 of Paleocanna tentaculum n. gen. n. sp. Entire specimen at left, with boxed region indicatng the location of the mapped section. The figure includes a photograph of the mapped region and corresponding element maps. Elemental composition percentages are as follows: aluminum (13%), carbon (6%), calcium (6%), iron (1%), potassium (3%), magnesium (5%), sodium (2%), oxygen (32%), phosphorus (2%), sulfur (3%), and silicon (26%). These values indicate that organisms are mostly carbon, slightly enriched in calcium or magnesium, whereas the matrix is rich in aluminum and potassium. Scale bars = 1 mm.

Remarks
Paleocanna tentaculum n. gen. n. sp. is the first soft-bodied cnidarian polyp to be formally described from the Ordovician of North America, distinct from conulariids, anthozoan corals, and Sphenothallus Hall, Reference Hall1847.
Phylogenetic results
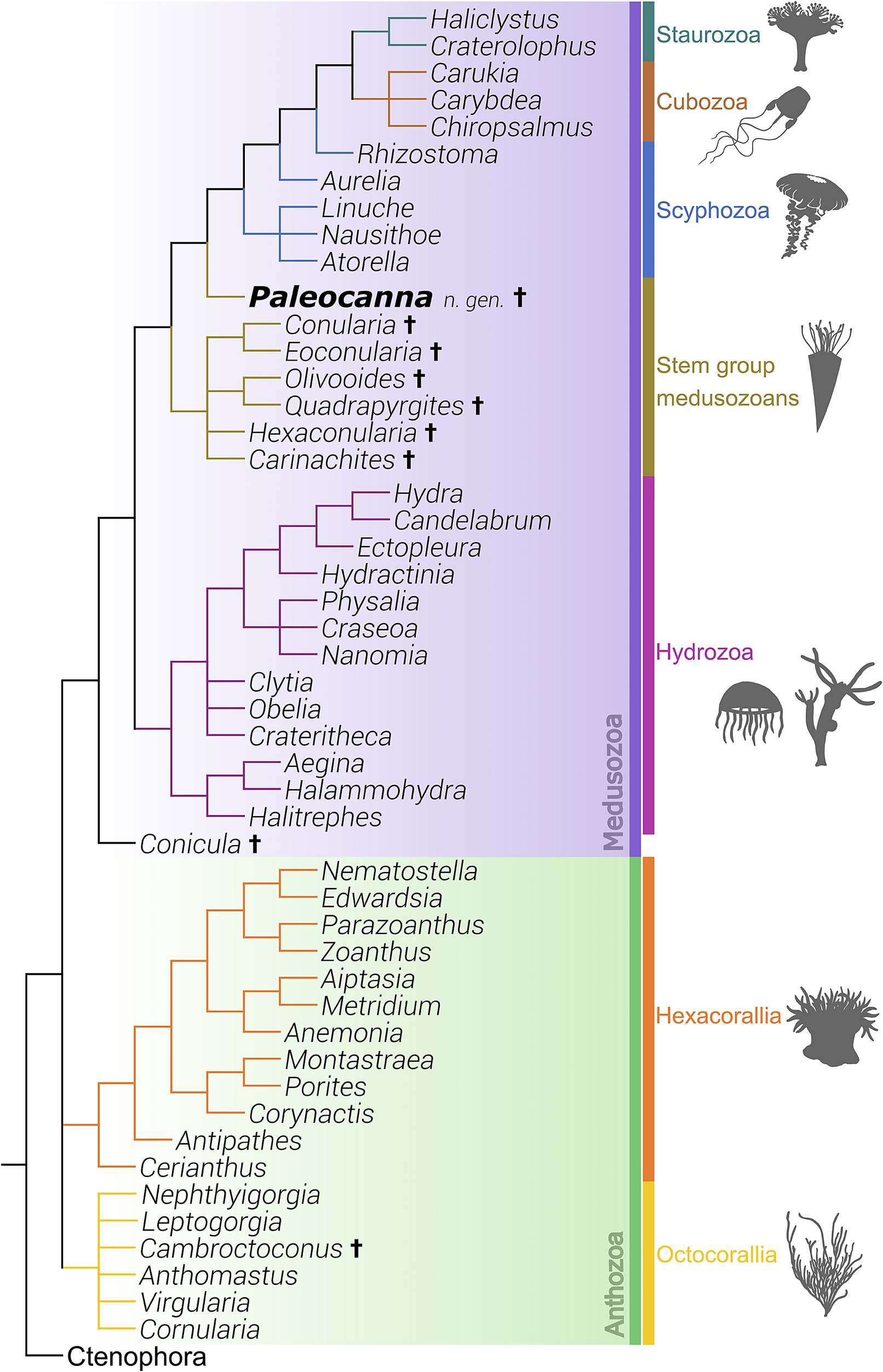
The most recent phylogenies of cnidarians place the Anthozoa (Hexacorallia and Octocorallia) as sister group to Medusozoa (Staurozoa, Scyphozoa, Cubozoa, and Hydrozoa) (Van Iten et al., Reference Van Iten, Marques, Leme, Pacheco and Simoes2014; Forero-Mejia, Reference Forero-Mejia, Molodtsova, Ostman, Bavestrello and Rouse2020). Paleocanna n. gen. placed among other cnidarian taxa, within the medusozoan lineage (Fig. 7). Most fossil cnidarian polyps occupy positions as sister groups to either the total-group Medusozoa stem or the anthozoan clade (Dunn et al., Reference Dunn, Kenchington, Parry, Clark, Kendall and Wilby2022; Ou et al., Reference Ou, Shu, Zhang, Han and Van Iten2022; Zhang et al., Reference Zhang, Parry, Vinther and Ma2022; Qu et al., Reference Qu, Li and Ou2023; Zhao et al., Reference Zhao, Parry, Vinther, Dunn, Li, Wei, Hou and Cong2023), whereas Paleocanna n. gen. is resolved as a basal crown Medusozoan, nested nearer to the clade containing cubozoans, scyphozoans, and staurozoans.
Phylogeny of Cnidaria, with Paleocanna tentaculum n. gen. n. sp. Strict consensus tree generated using 69 ingroup taxa and 236 discrete hard and soft-part morphological characters, polarized using species of the phylum Ctenophora as the sole outgroup. Paleocanna tentaculum n. gen. n. sp. is resolved as an early branching member of the crown group Medusozoa, more closely related to the clade comprising Cubozoa, Scyphozoa, and Staurozoa, than to other fossil medusozoans, e.g., conulariids. Animal silhouettes from PhyloPic (http://phylopic.org/). Taxa not otherwise mentioned in the text are Aegina Eschscholtz, Reference Eschscholtz1829; Aiptasia Gosse, Reference Gosse1858; Anemonia Risso, Reference Risso1827; Anthomastus Verrill, Reference Verrill1878; Antipathes Pallas, Reference Pallas1766; Atorella Vanhöffen, Reference Vanhöffen1902; Aurelia Lamarck, Reference Lamarck1816; Candelabrum de Blainville, Reference De Blainville and Levrault1830; Carukia Southcott, Reference Southcott1967; Carybdea Peron and Lesueur, Reference Peron and Lesueur1810; Cerianthus Delle Chiaje, Reference Delle Chiaje1841; Chiropsalmus Agassiz, Reference Agassiz1862; Clytia Lamouroux, Reference Lamouroux1812; Corynactis Allman, Reference Allman1846; Craseoa Pugh and Harbison, Reference Pugh and Harbison1987; Crateritheca Stechow, Reference Stechow1921; Craterolophus James-Clark, Reference James-Clark1863; Ectopleura Agassiz, Reference Agassiz1862; Edwardsia Quatrefages, Reference Quatrefages1842; Eoconularia Sinclair, Reference Sinclair1943; Halammohydra Remane, Reference Remane1927; Haliclystus James-Clark, Reference James-Clark1863; Halitrephes Bigelow, Reference Bigelow1909; Hydra Linnaeus, Reference Linnaeus1758; Hydractinia Van Beneden, Reference van Beneden1844; Leptogorgia Milne Edwards, Reference Milne Edwards1857; Linuche Eschscholtz, Reference Eschscholtz1829; Metridium de Blainville, Reference De Blainville and Levrault1824; Montastraea de Blainville, Reference De Blainville and Levrault1830, Nanomia Agassiz, Reference Agassiz1865; Nausithoe Kölliker, Reference Kölliker1853; Nematostella Stephenson, Reference Stephenson1935; Nephthyigorgia Kükkenthal, Reference Kükenthal, Michaelsen and Hartmeyer1910; Obelia Peron and Lesueur, Reference Peron and Lesueur1810; Parazoanthus Haddon and Shackleton, Reference Haddon and Shackleton1891; Physalia Lamarck, Reference Lamarck1801; Porites Link, Reference Link1807; Quadrapyrgites Li et al., Reference Li, Hua, Zhang, Zhang, Jin and Liu2007; Rhizostoma Cuvier, Reference Cuvier1800; Virgularia Lamarck, Reference Lamarck1816; and Zoanthus Lamarck, Reference Lamarck1801.

Paleocanna tentaculum n. gen. n. sp. was placed in the phylogeny based on external morphological characters, including a tubular, organic periderm that extended along the entire polyp, and bore widespread striae-like annulations, Ecologically, it is inferred to have been solitary to colonial, with a microphagous feeding strategy. Characters associated with the preservation of soft tissues, e.g., tentacles, set Paleocanna n. gen. apart from comparable extinct polypoid taxa that are typically known only from their mineralized periderm structures.
Symmetry is a key morphological character in early medusozoan evolution. In Paleocanna n. gen., the preserved structures suggest some sort of radial symmetry, consistent with a polypoid cnidarian body plan. However, the lack of clear longitudinal ridges or lamellae differentiates it from taxa like Conularia Sowerby, Reference Sowerby1821 and the hexangulaconulariids, which typically exhibit well-defined tetraradial or hexaradial symmetry with fan-shaped geometries (Song et al., Reference Song, Guo, Han, Van Iten and Peng2024). Similarly, taxa like Carinachites and Olivooides show more derived symmetry with pronounced apical structures and often exhibit quadriradial to pentaradial symmetry (Jian et al., Reference Jian, Guoxiang, Shin, Qiang and Sho2016; Song et al., Reference Song, Guo, Han, Van Iten and Peng2024).
Paleocanna n. gen. nested outside the clade that contains other extinct taxa; this suggests that the genus is part of a lineage of medusozoans, possibly related to conulariids or similar extinct forms. The new medusozoan reflects both morphological and taphonomic diversity within the group. The presence of other extinct medusozoans implies that Paleocanna n. gen. represents a lineage that was relatively diverse in the past.
Discussion
Paleocanna tentaculum n. gen. n. sp. is an Upper Ordovician tubicolous polyp medusozoan from the Saint-Joachim Quarry, Neuville Formation, Québec. The tubes are interpreted as epibenthic, likely oriented perpendicular to the seafloor, sometimes found attached to one another, and possibly buried in or attached to the substratum (Fig. 3). The tubular periderm that extends along the polyp exhibits widespread, continuous, fine annulations. The occurrence of attached individuals suggests a facultative semicolonial mode of life with clusters of up to 15 specimens (Fig. 3.2). Based on its morphology, a polypoid phase is inferred, with the polyp possessing a stalk or peduncle.
Although the tentacles cannot be observed in detail, they differ from those of previously reported cnidarian species in being fine and numerous. These contrast with tentacles of Gangtoucunia Luo et al., Reference Luo, Hu, Chen, Zhang and Tao1999 and Paleoconotuba, which are short and thick (Qu et al., Reference Qu, Li and Ou2023; Zhang et al., Reference Zhang, Parry, Vinther and Ma2022), and those of Nailiana Ou et al., Reference Ou, Shu, Zhang, Han and Van Iten2022 and Conicula Luo et al., Reference Luo, Hu, Chen, Zhang and Tao1999 that are smooth, finger-like, and longer compared to those of other fossil species (Ou et al., Reference Ou, Shu, Zhang, Han and Van Iten2022; Zhao et al., Reference Zhao, Parry, Vinther, Dunn, Li, Wei, Hou and Cong2023). The number of tentacles also varies among genera from at least five in Gangtoucunia, at least six in Conicula, from three to five in Palaeoconotuba, and eight in Nailiana. Although the phylogenetic position of Xianguangia sinica is uncertain, its fine tentacles suggest that it was a filter feeder (Ou et al., Reference Ou, Han, Zhang, Shu, Sun and Mayer2017). Following this same logic, the fine tentacles of Paleocanna tentaculum n. gen. n. sp. could have been used to filter food, by capturing food by gravitational deposition or by tentacle motion.
Due to the nature of soft-bodied preservation and the lack of part-counterpart material, it is not possible to completely rule out the possibility that the apparent termination of the gut is an artifact of preservation. However, the digestive structure appears consistently truncated at its posterior end across multiple specimens, with no evidence of continuation, disruption, or displacement (Fig. 2.1, 2.7). The preservation of surrounding tissues also suggests that the truncation is not the result of damage or incomplete exposure. Based on its repeated expression and anatomical context, we interpret this morphology as a blind gut.
Tubiculous cnidarians have existed at least since the Ediacaran, with genera such as Cloudina Germs, Reference Germs1972; Corumbella Hahn et al., Reference Hahn, Hahn, Leonardos, Pflug and Walde1982; Haootia Liu et al., Reference Liu, Matthews, Menon, McIlroy and Brasier2014; and Paraconularia Sinclair, Reference Sinclair1940 (Babcock et al., Reference Babcock, Grunow, Sadowski and Leslie2005; Vinn and Zatón, Reference Vinn and Zatoń2012; Liu et al., Reference Liu, Matthews, Menon, McIlroy and Brasier2014; Leme et al., Reference Leme, Van Iten and Simões2022). Auroralumina attenboroughii, also from the Ediacaran, appears to have been a transitional species, because it occupies a morphospace closer to Cambrian tubicolous genera, e.g., Cambroctoconus Park et al., Reference Park, Woo, Lee, Lee, Lee, Han, Chough and Choi2011, Carinachites, and Olivooides (Dunn et al., Reference Dunn, Kenchington, Parry, Clark, Kendall and Wilby2022). These taxa exhibit characteristics of both anthozoans and medusozoans (Dunn et al., Reference Dunn, Kenchington, Parry, Clark, Kendall and Wilby2022). The early Cambrian Gangtoucunia aspera provides evidence for the origin of biomineralization in cnidarians (Zhang et al., Reference Zhang, Parry, Vinther and Ma2022). Tubular fossils diversified into the Cambrian, suggesting adaptations to different ecological niches.
Paleocanna n. gen. differs from most of these species in having a tubular periderm rather than a conical one, as well as in having finely striated annulations instead of the sharper rib- or crest-like structures. This tubular body is preserved as a carbonaceous film, as confirmed by elemental analysis, and the annulations along its length suggest some degree of structural integrity. However, there is no indication of a rigid or biomineralized wall; the tube appears to have been originally organic. Some of these thecate species also exhibit longitudinal ridges and midlines characteristic of biomineralized conulariids, a trait not observed in Paleocanna tentaculum n. gen. n. sp. This distinction is remarkable given that most stem-group medusozoans are known from the Cambrian, whereas Ordovician assemblages are predominantly composed of conulariids, which were both diverse and widespread during this period. Paleocanna n. gen. more closely resembles Cambrian forms and appears to co-occur with conulariids, possibly representing a different phylogenetic branch. At least one species of conulariid is known from the locality, based on fragmentary but recognizable specimens. These conulariids do not exhibit evidence of soft-tissue preservation.
The annulations observed in the periderm of Paleocanna n. gen. bear some resemblance to those of Sphenothallus, which is also known to be preserved as carbonaceous compressions (Muscente and Xiao, Reference Muscente and Xiao2015). However, several key differences suggest a distinct identity. Sphenothallus is typically preserved as a phosphatic structure with longitudinal thickenings and transverse walls (Muscente and Xiao, Reference Muscente and Xiao2015; Van Iten et al., Reference Van Iten, Gašparič, Hitij, Kolar-Jurkovšek and Jurkovšek2023), whereas Paleocanna n. gen. appears to be a full thin-walled tube and lacks any indication of phosphatization. Morphologically, Paleocanna n. gen. lacks the pronounced tapering and pointed apex usually seen in Sphenothallus. The latter is also most often found as isolated individuals, whereas Paleocanna n. gen. commonly occurs in clusters, potentially indicating a different life mode.
The growth of Paleocanna tentaculum n. gen. n. sp. was allometric, exhibiting minimal variation in width as the lengths increased, with a ± 36.9 mm length and ± 6.0mm width (N = 39). Interpretation of this morphometric data requires caution due to the soft-bodied nature of cnidarians and the potential alterations in actual biological measurements caused by the fossilization process. Despite these challenges, allometric growth characterizes the development of most animals, and has been present since the Cambrian in trilobites, for example (Du et al., Reference Du, Peng, Wang, Wang and Wang2019; Holmes et al., Reference Holmes, Paterson and García-Bellido2021).
Paleocanna tentaculum n. gen. n. sp. was buried in a low-energy, anoxic, muddy bottom environment favorable to the preservation of solid organic matter and labile animal soft tissues. Elemental analysis of MPEP713.3 revealed pure carbon although a portion of the material appeared to be marginally enriched in calcium or magnesium, whereas the matrix itself exhibited higher concentrations of aluminum and potassium. None of these elements can be original, except for carbon. This mineral composition bears some resemblance to Burgess Shale-type (BST) fossilization. The siliciclastic deposits that preserve BST fossils have high clay-to-organic ratios, which is one way to facilitate exceptional preservation by slowing the rate of decay (Anderson et al., Reference Anderson, Tosca, Gaines, Mongiardino Koch and Briggs2018). The preservation of these cnidarian fossils is attributed to the transportation of the organisms within a moving sediment cloud and subsequent burial with all hollows and spaces filled with fine-grained clays (Gaines et al., Reference Gaines, Hammarlund, Hou, Qi, Gabbott, Zhao, Peng and Canfield2012).
The Saint Joachim quarry fossils and shales exhibit the characteristics of a BST locality, including their occurrence in fine-grained sediment, the quality of preservation, high organic content, a low-energy depositional setting, and likely anoxic conditions. Comparable soft-bodied cnidarians have been described from the Burgess Shale, including a medusa with remarkable preservation of internal anatomy, e.g., gonads (Moon et al., Reference Moon, Caron and Moysiuk2023). Although the Saint Joachim fossils are more limited in anatomical detail, they nonetheless preserve key morphological features including annulated tubes and tentacle impressions.
Understanding the preservation of soft-bodied organisms like Paleocanna tentaculum n. gen. n. sp. provides a valuable contrast to most Ordovician deposits, which typically feature only mineralized organisms, especially given the rarity of soft-tissue preservation during this period (Moysiuk et al., Reference Moysiuk, Izquierdo-López, Kampouris and Caron2022). This rare type of preservation is especially relevant in a group that represents some of the earliest metazoans to biomineralize, and in a locality where the associated biota includes typical hard-bodied species like bryozoans, brachiopods, cephalopods, trilobites, and echinoderms (Clark and Globensky, Reference Clark and Globensky1973; Harland and Pickerill, Reference Harland and Pickerill1982).
Conclusions
Paleocanna tentaculum n. gen. n. sp. represents a remarkable discovery from the Upper Ordovician Neuville Formation in Québec. As a tubicolous medusozoan that is more closely related to the living medusozoan clade containing Staurozoa, Cubozoa, Scyphozoa, than to the other fossil genera, this species shares several morphological and ecological similarities with both modern anthozoans and medusozoans, including an annulated periderm and a digestive tract, the structure of the tentacles, and an epibenthic lifestyle. The fossil evidence suggests semicolonial behavior with their tubes being attached to surfaces or buried in the sediment. This discovery provides a window onto the complex ecosystems of the Ordovician Period, offering an important comparison to both modern cnidarians and other fossilized medusozoans from earlier periods.
The preservation of soft tissues in Paleocanna tentaculum n. gen. n. sp. was facilitated by burial under environmental conditions of very low physical energy and sediment pore-water anoxia. This rare type of preservation is significant because it challenges the typical Ordovician fossil record, which predominantly features organisms with mineralized hard parts. Elemental analysis suggests that organic carbon was preserved, making this an important site for understanding the biology and fossilization of ancient soft-bodied organisms. The insights provided by Paleocanna tentaculum n. gen. n. sp. help fill gaps in our understanding of early anthozoan evolution, and how these ancient soft-bodied organisms adapted to and thrived in marine environments.
Acknowledgments
Financial support was provided by NSERC Discovery grant 2023-05647 to CBC and a FRQNT Ph.D. bursary to GRG. We thank J. Iellamo who collected most of the specimens, understood their scientific value, and made them available for study. We also want to thank M. Meacher who collected and donated specimen MPEP1525.1, as well as J.-B. Caron for his valuable assistance with the elemental analysis. Thanks to our reviewers for their comments and suggestions that improved our manuscript.
Competing interests
The authors declare no competing interests.
Data availability statement
The character matrix is available through the Morphobank repository (http://doi.org/10.7934/P6124).





 Open access
Open access