




Introduction
Extensive studies have demonstrated that the exploitation of and adaptation to marine environments long played an important role in the cultural and economic evolution of human societies since the late Middle Pleistocene (c. 190–130 ka, see reviews in Will et al. Reference Will2019; Rick Reference Rick2024). Although such maritime adaptations may be diverse, the regular and systematic use of marine foods is evident in this period through the widespread appearance of large shell deposits or middens in the archaeological record (Zhang & Hung Reference Zhang and Hung2016; Will et al. Reference Will2019). Adaptive strategies for island environments in the Indo-Pacific region have been the subject of recent research concerning prehistoric human movements and the colonisation of remote islands (e.g. Takamiya Reference Takamiya2006; Fujita et al. Reference Fujita2016; Summerhayes et al. Reference Summerhayes, Bedford and Spriggs2019; Kerby et al. Reference Kerby2022; Amano et al. Reference Amano2024; Faylona & Pawlik Reference Faylona and Pawlik2025). These studies, focusing on the analysis of lithics and faunal and shell assemblages, present evidence of maritime adaptations reflecting regional diversity, resource availability and local ecological conditions. Yet, the overall dietary importance of marine resources is difficult to estimate and reconstructions of palaeoenvironmental and palaeoecological conditions during phases of human activity are incomplete.
Analyses of stable carbon (δ13C), nitrogen (δ15N) and oxygen (δ18O) isotopes, which vary in predictable ways between the different trophic levels of food webs and between different foraging environments (i.e. terrestrial and marine ecosystems), have already been conducted to explore human and animal diets and ecological conditions on islands from South and East Asia to Near and Remote Oceania, providing direct insights into the long-term adaptive flexibility of humans in maritime environments (Choy et al. Reference Choy2012; Kinaston et al. Reference Kinaston2014; Kusaka et al. Reference Kusaka2015; Commendador et al. Reference Commendador2019; Stone et al. Reference Stone2019; Roberts et al. Reference Roberts2020; Roberts et al. Reference Roberts2022a & Reference Robertsb). Analysis of these isotopes in incremental dentine samples has also been used to elucidate the early lives of islanders and changes to their diets across their lifespan (e.g. Montgomery et al. Reference Montgomery2013; Stantis et al. Reference Stantis2021).
The coastal and inland settings of the south-east China coast, a key region in discussions of Austronesian origins and dispersals (Bellwood Reference Bellwood1984; Yang et al. Reference Yang2020), have been the focus of a growing research interest in local prehistoric marine adaptations and subsistence strategies (Zhang & Hung Reference Zhang and Hung2016; He et al. Reference He2023; Zhou et al. Reference Zhou2025). A recent study integrating radiocarbon dating and the analysis of cultural deposits and microfossils at the Jingtoushan site (Figure 1) traces Neolithic human marine adaptation to 8000 cal BP on the mainland coast (He et al. Reference He2023), yet lacks direct evidence from human remains. Well-preserved human remains dating from the Neolithic (c. 10 000–4000 BP) are scarce in this region, but a few studies do show the importance of marine foods in the diets of coastal humans. To date, the earliest radiocarbon dates and isotopic data from humans in this region are provided by two human skeletons found on Liangdao Island (Chen Reference Chen2013; Chiu et al. Reference Chiu2015), dating to 8300–7500 cal BP and reflecting a heavy reliance on marine foods. Isotopic studies of several later sites (c. 7000–6000 BP) on the mainland coast provide evidence of a diverse marine and terrestrial diet (Zhang et al. Reference Zhang2003; Hu et al. Reference Hu2010; Minagawa et al. Reference Minagawa2011; Dong Reference Dong2016). However, direct evidence for the diet of early Neolithic humans on islands and an assessment of their adaptive strategies concerning ecological diversity remain to be further investigated.
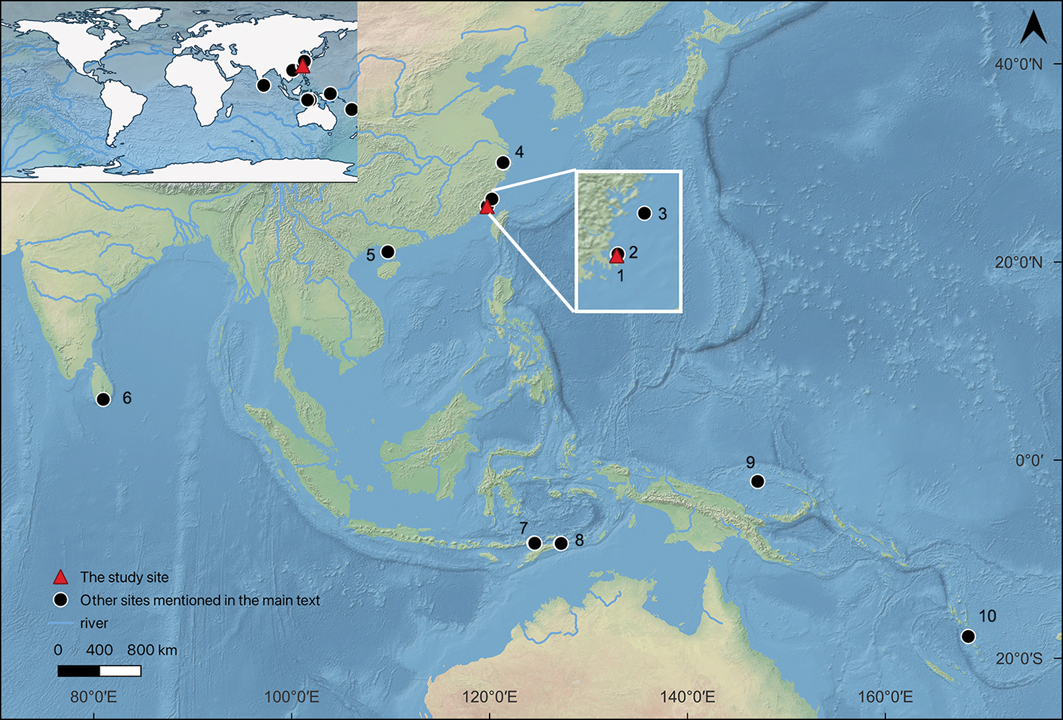
Map showing the location of the studied site (red triangle) and other sites mentioned in the text (black dots): 1) Xiying; 2) Keqiutou, Donghuaqiu and Guishan; 3) Liangdao; 4) Jingtoushan, Tianluoshan and Hemudu; 5) Liyudun; 6) Mini-athiliya; 7) Makpan and Tron Bon Lei; 8) Matja Kuru 1 and 2, Asitau Kuru and Lene Hara; 9) Pamwak; 10) Teouma (figure by authors).

In this article, we undertake an analysis of human and animal remains recently excavated from the Xiying site on Haitan Island (Figure 1), possibly the earliest human remains yet found on the island. Stable isotope analysis (carbon, nitrogen and oxygen) of bone and enamel from terrestrial and marine animals helps reconstruct the foraging environments and ecological conditions on Early Neolithic Haitan Island, and provides a faunal baseline for human dietary determinations from bulk and incremental (dentine) stable isotope analyses. Seven human bone samples also provide new radiocarbon dates for activity on the island. Comparison of our new dating and isotopic data with existing data from the south-east coast of China and the islands of South Asia, Southeast Asia and Oceania provides new insights into the timing and nature of maritime adaptations of Early Neolithic islanders on the south China coast, contributing to global discussions on human adaptive flexibility in marine environments.
Archaeological context
The Xiying site is located in the eastern part of Xiying Natural Village, Suping Town, Fujian Province, China (25°36′13.2″N, 119°44′25.4″E), at an elevation of about 10m with the northern coastal mountains of Haitan Island to the north-west (Figure 1). The site was discovered in 1992, and preliminary surveys in 2022 revealed a preserved area of approximately 1ha. From 2022 to 2023, the School of History and Cultural Heritage, Xiamen University, conducted two excavations in the southern part of the site: Areas I and II, separated by a modern pathway (Figure 2). The excavated features included one shell deposit, two houses, 15 postholes, three burials, seven ash pits and three ash ditches. Pottery (including fu cauldrons with round bottoms (圜底釜), guan jars (罐) and bo bowls (钵)), stone tools (hammers, pitted pebbles, choppers, scrapers, adzes and spindle whorls) and bone artefacts were unearthed (IA CASS et al. 2024). The main cultural layers date to approximately 7400–6400 BP, representing the earliest Neolithic remains discovered on Haitan Island to date (IA CASS et al. 2024).
a) Aerial view of excavations at the Xiying site in 2023 (from IA CASS et al. 2024). b–d) Human burials discovered during the 2023 excavation season (figure by authors).

During the 2022 excavation, human bone fragments (including parts of the cranium, mandible, humerus and femur) and teeth (incisors, canines and molars) were recovered from the western part of the shell deposit in Area I and from a ditch that intruded into the shell deposit, representing at least four individuals. In 2023, three burials (M1, M2 and M3; Figure 2b–d) were discovered along the western edge of the shell deposit in Area I, displaying consistent orientations, body placement (extended supine) and dimensions. Osteological examination of all human remains were conducted by Dr Qun Zhang at the Department of Archaeology, Wuhan University. The occupants of M1 and M3 were most likely males, estimated to have been 28–32 years and an adult with indeterminate age, respectively; the age of the individual in M2 was about 30 years but the sex could not be determined based on skeletal morphology.
A large shell deposit (designated BK), marine and terrestrial animal bones and a small quantity of charred plant seeds were found at the site. More than 10 shell species were identified in BK, including oysters, mud clams, hard clams and the trumpet shell whelk (IA CASS et al. 2024). Marine and terrestrial animals, including deer, water buffalo, small carnivorous mammals, birds, dolphins, freshwater and sea turtles, and fish, were identified by Dr Chong Yu at the School of Sociology and Anthropology, Sun Yat-sen University. Radiocarbon dating of shell and animal bone samples yielded results of 7400–6500 cal BP (IA CASS et al. 2024). One immature rice grain, one fragment of charred rice and one rice spikelet were found in BK (IA CASS et al. 2024) but were not submitted for radiocarbon dating due to the small number of plant remains recovered.
Materials and methods
A total of 38 animal bone samples, including small-sized deer (n = 3), medium-sized deer (n = 13), water buffalo (n = 3), freshwater turtle (n = 7), sea turtle (n = 5), mullet (n = 4) and marine fish (unidentified species, n = 3), were taken for carbon and nitrogen stable isotope analyses to provide a faunal isotopic baseline for the site. Deer (Cervidae), as herbivores, tolerate a wide variety of habitats; their isotopic composition directly reflects environmental influences via the plants consumed and is consequently used as an indicator in palaeoenvironmental and palaeoecological studies (Stevens et al. Reference Stevens2014, Reference Stevens2025). Thus, relatively well-preserved tooth enamel samples from deer (four small-sized deer and 10 medium-sized deer) were also sampled for carbon and oxygen stable isotope analyses, to reconstruct the environmental and ecological conditions at the site.
Seven human bone samples, including the four individuals identified in 2022 and the three individuals from burials discovered in 2023, were collected for carbon and nitrogen stable isotope analysis and for radiocarbon dating at Beta Analytic (Miami, USA) and at the Radiocarbon Laboratory of Peking University. Three human teeth (two canines and one incisor, covering approximately 0.5–14.5 years of age), each from a different individual, were sampled for sequential carbon and nitrogen stable isotope analysis (dentine collagen), and carbon and oxygen stable isotope analysis (enamel), to understand the dietary composition of Xiying individuals across life courses. Detailed information about all materials and the methods used to analyse them is included in the online supplementary material (OSM). Seven human bones, 20 tooth dentine serial sections and 23 animal bones yielded well-preserved collagen that meet the collagen quality criteria for carbon and nitrogen stable isotope analysis (for detail, see the OSM) and were included for further discussion. Tooth enamel is more resistant to diagenetic alteration and is widely used as a suitable candidate for stable isotope analysis (Kohn & Cerling Reference Kohn and Cerling2002). The enamel samples we selected for pretreatment were all relatively well preserved and the isotopic results from all samples were included for further analysis.
Results
Radiocarbon dating
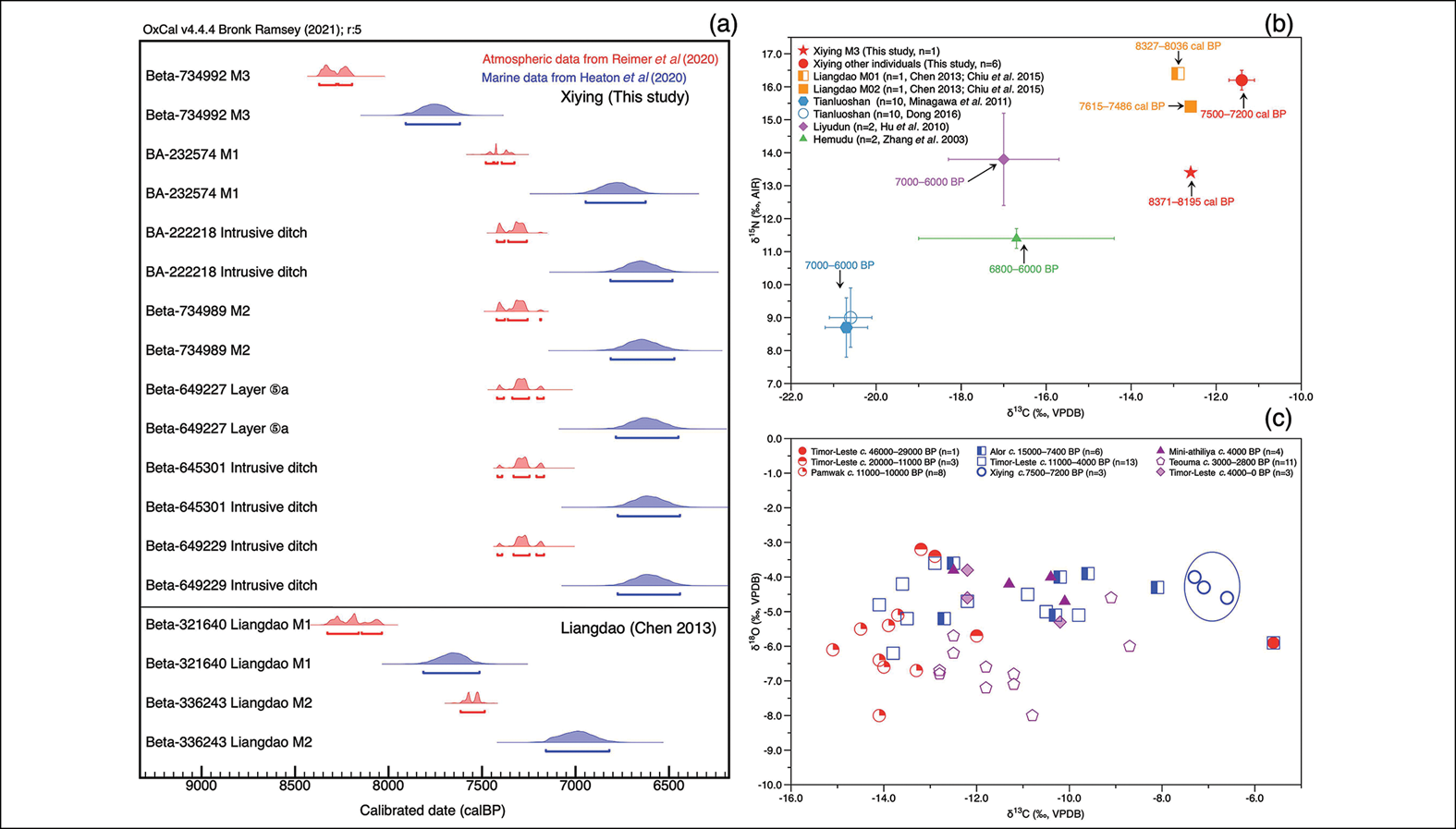
Radiocarbon dates for the seven human bone samples are presented in full in Table S5. Calibrating dates from coastal human skeletal remains is challenging due to the spatially and temporally variable marine reservoir effect and dietary 14C offsets caused by mixed marine and terrestrial food consumption (Stuiver & Braziunas Reference Stuiver and Braziunas1993; Petchey et al. Reference Petchey2014). Since no local accurate marine reservoir correction (ΔR) is currently available for the study area (Zhang et al. Reference Zhang2025), we calibrated all radiocarbon dates twice, once using the IntCal20 curve (Reimer et al. Reference Reimer2020) and once using the Marine20 curve (Heaton et al. Reference Heaton2020). Calibration using the IntCal20 curve may yield calibrated ages that are older than the true ages (average 400 years for global average surface water marine reservoir effect), while calibration using the Marine20 curve without a local accurate ΔR value and percentage estimates of the marine dietary component may result in younger ages (Stuiver & Braziunas Reference Stuiver and Braziunas1993; Reimer et al. Reference Reimer2013; Zhang et al. Reference Zhang2025). The true dates probably lie somewhere in between the results of the two calibrations. If regional ΔR values become available in the future, it will be necessary to recalibrate the existing data.
The calibrated dates for most Xiying individuals range from c. 7500–7200 cal BP at 95.4% confidence using the IntCal20 calibration curve and c. 7000–6500 cal BP at 95.4% confidence using the Marine20 calibration curve. The individual from M3 yielded an outlier date of 7480±30 BP (8371–8195 cal BP at 95.4% confidence using IntCal20; 7908–7619 cal BP at 95.4% confidence using Marine20). The main date range is consistent with the chronology of the archaeological remains from the site, corresponding to the Early Neolithic period.
Isotopic data from bone and dentine collagen
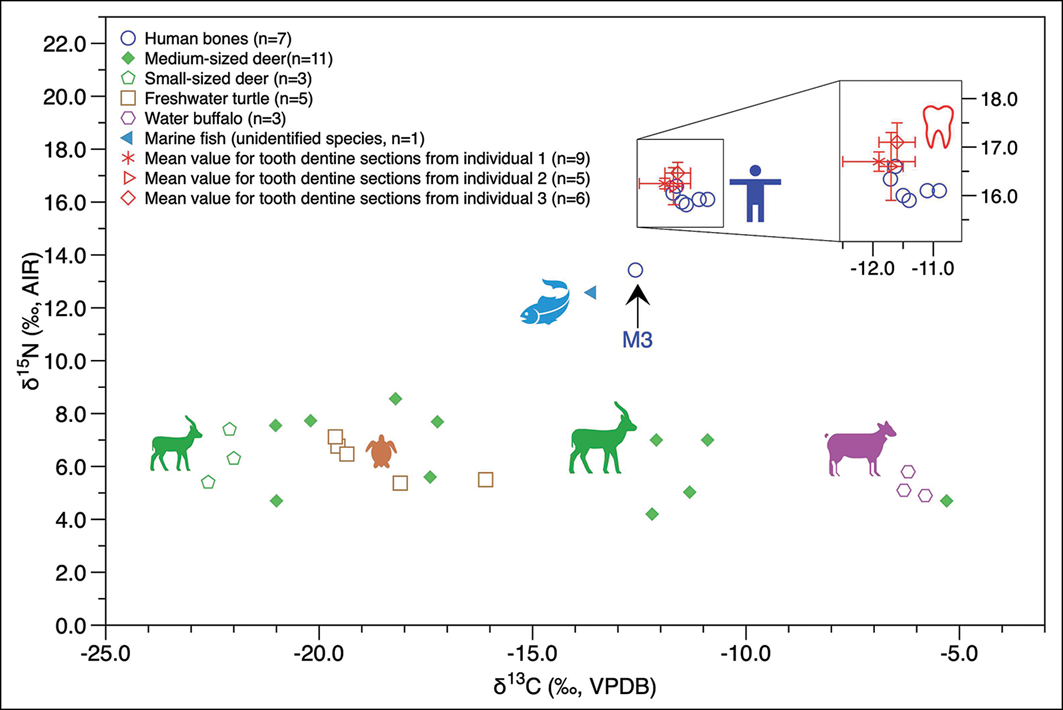
Figure 3 presents the scatterplot of isotope data for humans (including bones and dentine serial sections) and animals from Xiying. The δ13C values for terrestrial herbivores (small-sized deer, medium-sized deer, water buffalo; n = 17) range from −22.6‰ to −5.3‰ (mean = −14.8±6.4‰), and the δ15N values range from 4.2‰ to 8.6‰ (mean = 6.2±1.3‰). The δ13C values for freshwater turtles (n = 5) range from −19.6‰ to −16.1‰ (mean = −18.6±1.5‰), and the δ15N values range from 5.4‰ to 7.1‰ (mean = 6.3±0.8‰). The one marine fish (unidentified species) provides a δ13C value of −13.6‰ and a δ15N value of 12.6‰. These results indicate a wide range of diets for terrestrial herbivores, suggesting an environment covered by a mix of C3 and C4 vegetation, and the complexity of aquatic ecosystems.
Human and animal δ13C and δ15N values from Xiying. The arrow indicates the data from burial M3 (figure by authors).

The δ13C values of human bones range from −12.6‰ to −10.9‰ (mean = −11.5±0.5‰, n = 7), and the δ15N values range from 13.4‰ to 16.6‰ (mean = 15.8±1.1‰, n = 7), suggesting a diet high in animal protein. The δ13Cdentine values of incremental dentine samples range from −13.0‰ to −11.2‰ with a mean value of −11.8±0.5‰; the δ15Ndentine values range from 16.0‰ to 17.7‰ with a mean value of 16.8±0.5‰ (n = 20). Except for the individual from M3, the δ13C (mean = −11.4±0.3‰, n = 6) and δ15N (mean = 16.2±0.3‰, n = 6) values of adult bone collagen are close to the values of tooth dentine sections.
Isotopic data from enamel
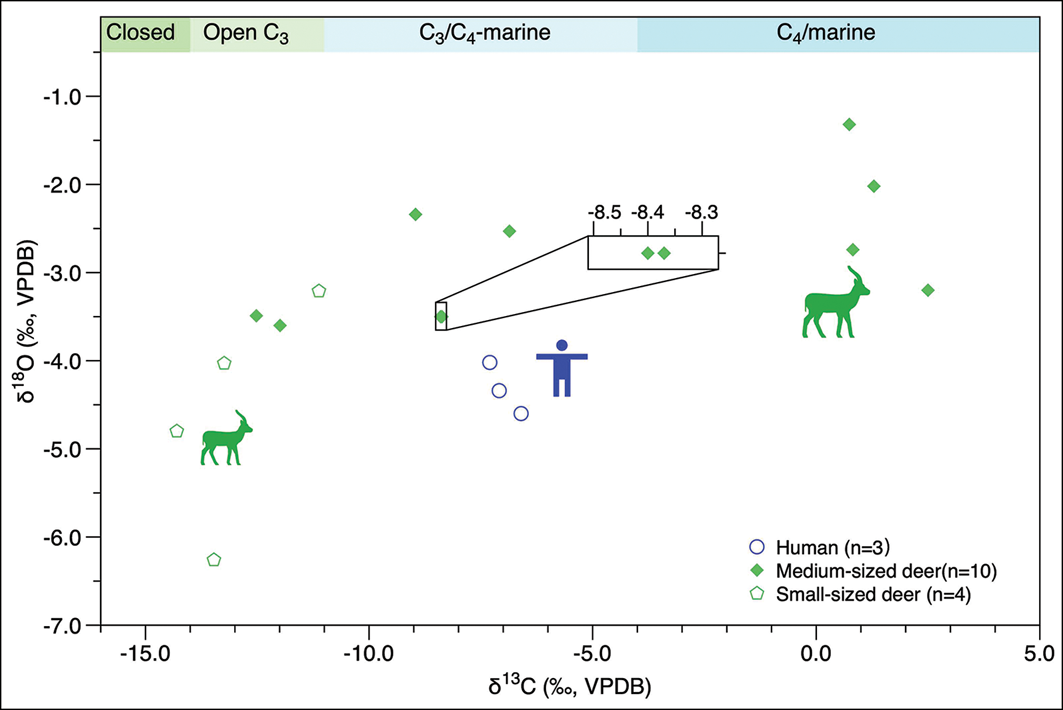
Figure 4 presents the scatterplot of δ13C and δ18O values from human and animal dental enamel bioapatite. The small-sized deer (n = 4) display an average δ13Cenamel value of −13.0±1.3‰ (range: −14.3‰ to −11.1‰) and an average δ18Oenamel value of −4.6±1.3‰ (range: −6.3‰ to −3.2‰), suggesting they inhabited a mosaic of closed forest conditions and open grassland environments, probably drinking surface water and browsing on lower-canopy leaves or plants with lower δ13C and δ18O values.
Human and animal δ13C and δ18O values from Xiying. Approximate cut-offs of δ13C values for diets relying completely on closed forest (–14‰), open C3 settings (–11‰) and C4/marine resources (–4‰∼0‰) are indicated based on previous studies conducted in Southeast Asia, Oceania and Japan (Kusaka et al. Reference Kusaka2015; Roberts et al. Reference Roberts2017, Reference Roberts2020) (figure by authors).

The medium-sized deer (n = 10) display an average δ13Cenamel value of −5.2±5.9‰ (range: −12.5‰ to 2.5‰) and an average δ18Oenamel value of −2.8±0.8‰ (range: −3.6‰ to −1.3‰). The relatively higher δ13C and δ18O values of medium-sized deer are characteristic of feeding and drinking in more open settings.
The δ13Cenamel values for humans range from −7.3‰ to −6.6‰ with a mean value of −7.0±0.4‰, and the δ18Oenamel values ranged from −4.6‰ to −4.0‰ with a mean value of −4.3±0.3‰ (n = 3), indicating a reliance on a mixture of open C3 and C4/marine resources.
Discussion
Foraging environments and ecological conditions on Early Neolithic Haitan Island
The feeding and drinking habits and ecological niches of deer, indicated by the broad range of δ13Cenamel, δ18Oenamel and δ13Ccollagen values, suggest a large spectrum of habitats and dietary resources on the island, from closed forests to open C3 (e.g. wetland plants) and C4 (e.g. grasses in sandy soils) or marine environments (Stevens et al. Reference Stevens2025). The much higher δ13Cenamel values of four medium-sized deer and δ13Ccollagen values of water buffalo (Figures 3 & 4) probably reflect an increased reliance on marine resources rather than C4 plants in open environments, as previous studies in the north-west of Europe have shown that terrestrial mammals (e.g. deer, hares, sheep) in coastal areas will naturally consume seaweed, resulting in δ13C values around 8–14‰ higher than animals relying on terrestrial vegetation (Conradt Reference Conradt2000; Balasse et al. Reference Balasse2009, Reference Balasse2019). Although a larger and more diverse animal dataset is needed for a more comprehensive interpretation, our faunal stable isotope data nevertheless present an insight into the diverse foraging environments and ecological conditions present on Early Neolithic Haitan Island, encompassing closed forests, open C3 vegetation, C4 resources and marine environments.
A homogeneous marine-based human diet in a broad range of habitats
With the associated faunal data providing a local baseline, we can use the δ13Ccollagen (mean = –11.5±0.5‰) and δ15Ncollagen (mean = 15.8±1.1‰) values of the human bones to infer that the individuals from Xiying occupied a very high trophic level (higher than that of marine fish, see Figure 3), and probably consumed a significant proportion of marine foods. Comparison of δ13C values from tooth enamel and bone collagen (Δ13Cenamel–collagen) offers another means of evaluating trophic levels (Lee-Thorp et al. Reference Lee-Thorp1989; Clementz et al. Reference Clementz2009). Due to variation in the lipid content of different diets, Δ13Cenamel–collagen is expected to be about 7.6‰ for herbivores and 4.8‰ for carnivores in terrestrial ecosystems, and approximately 7.8‰ for herbivores and 3.6‰ for carnivores in marine ecosystems (Clementz et al. Reference Clementz2009; Kusaka et al. Reference Kusaka2015). An average Δ13Cenamel–collagen value of approximately 4.5‰ is observed for the Xiying humans. In conjunction with the high δ15Ncollagen values, this suggests a diet incorporating higher trophic level foods, primarily from marine ecosystems.
Additionally, a narrow dietary range is observed for most individuals. Excluding the individual from M3, the standard deviations for human δ13C, δ15N and δ18O values are close to 0.3, consistent with the low isotopic variability typically associated with populations consuming a homogenous diet (standard deviation = 0.3; Lovell et al. Reference Lovell1986). Data from incremental dentine sections (see Figure 3) further suggest a homogenous marine-based diet from childhood through adulthood among the Xiying population; slightly higher early-life δ15N values can potentially be attributed to breastfeeding effects (Fuller et al. Reference Fuller2006). However, due to poor preservation complete isotopic profiles, representing diet from infancy through childhood to adolescence, could not be obtained from incremental dentine sections; acquisition of better-preserved teeth and more age-constrained incremental dentine samples would enable a more detailed discussion of individual life histories.
Overall, despite the availability of varied habitats, indicated by the faunal isotopic data, our results indicate that the individuals from Xiying specialised in the exploitation of high-trophic-level resources, consuming a marine-based diet across their lifespan, with some supplementary contributions from open C3 and C4 resources (see Figure 4).
Direct evidence of early maritime adaptation on the south-east China coast
To elucidate the heavy reliance of the Xiying individuals on marine resources within the broader context of human adaptive strategies, we compared our results with published findings on Early to Middle Neolithic humans from coastal China (Figure 5a & b). The early date of the individual from M3 at Xiying (8371–8195 cal BP at 95.4% confidence using IntCal20; 7908–7619 cal BP at 95.4% confidence using Marine20) may be comparable to that of Liangdao M1 (8327–8036 cal BP at 95.4% confidence using IntCal20; 7814–7514 cal BP at 95.4% confidence using Marine20; Chen Reference Chen2013). However, this is currently our only dating evidence prior to 8000 BP. A more comprehensive discussion of early human maritime adaptation around 8000 BP will only be possible if additional contemporaneous remains are recovered. The individual from M3 also delivered a δ15N value that was 2–3‰ lower than the other individuals from Xiying and the individuals found at Liangdao (Figure 5b). This disparity could be due to changes in the availability of resources, although verification from faunal baselines using earlier animal bones is needed, or could reflect the consumption of a greater proportion of marine foods from lower trophic levels. With this one exception, humans from the island sites of Xiying and Liangdao all had marine-based diets, displaying high δ13C and δ15N values compared to humans from the Hemudu, Tianluoshan and Liyudun sites in the coastal region of the mainland. More diverse diets are suggested for these latter sites, including both high and low trophic level foods from marine and terrestrial ecosystems. The specialised marine-based diets of Xiying and Liangdao humans may be an adaptive strategy to the island environment.
a) Calibrated radiocarbon dates for human bones from Xiying and Liangdao on the south-east China coast. b) Scatter plot of δ13C and δ15N mean values of human bone collagen from the Early and Middle Neolithic sites on the south-east China coast. c) Scatter plot of δ13C and δ18O values of human tooth enamel from Late Pleistocene to Late Holocene Neolithic sites on islands in South Asia, Southeast Asia and Oceania (the blue oval marks the data from this study, other data from Roberts et al. Reference Roberts2020, Reference Roberts2022a, Reference Roberts2022b). Locations of the mentioned sites are illustrated in Figure 1, raw data are listed in the OSM (figure by authors).

The distinctive environments of islands, often featuring small land areas, presented new challenges to prehistoric humans seeking new ways to reach new places (Mijares & Kaifu Reference Mijares and Kaifu2022). To assess potential adaptive strategies, we summarised available human enamel δ13C and δ18O values from Late Pleistocene to Late Holocene sites on the islands in South Asia, Southeast Asia and Oceania (Figure 5c). This revealed that early humans (before 10 000 BP) from Timor Island and Pamwak on Manus Island, and later humans (after 10 000 BP) from Alor Island, Timor Island, Teouma on Vanuatu island and Mini-athiliya on Sri Lanka, relied to varying degrees on C4/marine, closed forest and open C3 habitats. These data demonstrate the adaptive flexibility of prehistoric humans when meeting diverse island habitats. Yet, even within this dataset, the δ13C and δ18O values of Xiying individuals are among those showing the strongest dependence on marine resources.
The bones found at the Xiying site are the earliest human remains discovered on Haitan Island to date. Combining estimates for the main cultural remains (c. 7400–6400 BP) with the new dates presented here, we infer that human occupation on the island possibly dates no earlier than 7500 cal BP. Faunal isotopic data indicate a broad range of ecological resources on Haitan Island at this time and the identification of rice phytoliths and macroremains suggests that rice may have been cultivated on the island by 7000 BP (IA CASS et al. 2024; Zhou et al. Reference Zhou2025). However, our isotopic data show that the individuals buried at Xiying incorporated substantial amounts of high-trophic-level marine resources into their diets, and did not rely heavily on terrestrial resources like some populations on tropical or Oceania islands.
These dietary choices potentially reflect the foraging strategy of hunter-gatherers (Bettinger Reference Bettinger2009). The ocean provides a stable and continuous source of high-protein, high-return foods, including high-trophic-level fish such as the dolphins found at Xiying, which would be preferentially consumed over lower-protein, lower-return foods. In such a situation, groups can adapt to and sustain themselves within island environments. The technology of this adaptation, including stone tools, boats and fishing gear, awaits further discoveries for a deep discussion. Nevertheless, marine provision as a foundation for cultural development since around 7500 cal BP is supported by later Keqiutou, Donghuaqiu and Guishan sites (from c. 6500–3200 BP) on Haitan Island (IA CASS et al. 2024).
Conclusion
Investigating how prehistoric humans adapted to and sustained themselves in island environments expands our understanding of human relationships with aquatic habitats and the importance of marine resources in human cultural and economic evolution. Analysis of stable isotopes from the bones and teeth of terrestrial and marine animals at the Xiying site indicates diverse foraging environments and ecological conditions on Early Neolithic Haitan Island (including closed forests, open C3 vegetation, C4 resources and marine environments). Using this faunal baseline, the lifelong, specialised exploitation of high-trophic-level resources by the individuals buried at Xiying is revealed in their isotopic signatures. Embedding these data within the wider regional context provides evidence of early maritime adaptation on the south-east China coast, c. 7500 cal BP, and new insights into the dietary flexibility of prehistoric humans.
Acknowledgements
Special thanks to Dr Tianyang Gu for her assistance with sample preparation work. We also thank the editors and anonymous reviewers for their extremely helpful advice.
Funding statement
This work was supported by National Social Science Foundation of China (grant number 22CKG023).
Online supplementary material (OSM)
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2026.10335 and select the supplementary materials tab.
Author contributions: CRediT categories
Bing Yi: Conceptualization-Lead, Data curation-Lead, Formal analysis-Lead, Funding acquisition-Lead, Investigation-Equal, Methodology-Lead, Project administration-Lead, Resources-Equal, Software-Lead, Supervision-Lead, Validation-Lead, Visualization-Lead, Writing - original draft-Lead, Writing - review & editing-Lead. Qun Zhang: Data curation-Equal, Methodology-Equal. Chong Yu: Data curation-Equal, Methodology-Supporting. Yun Guo: Investigation-Equal, Resources-Equal. Weiyao Han: Resources-Equal. Yaowu Hu: Data curation-Equal, Formal analysis-Equal. Wenjie Zhang: Project administration-Lead, Resources-Lead.




 Open access
Open access