


Non-technical Summary
Fossil discoveries from the Riversleigh World Heritage Area in northwestern Queensland, Australia, have uncovered three previously unknown species of small, insect-eating marsupials that lived about 18 million years ago. Named Phantasmodon travouilloni, Phantasmodon minuferox, and Keeunidae gen. and sp. indet., these animals ranged from shrew- to mouse-sized. Their teeth share distinctive features with older Australian species and with Djarthia murgonensis, the continent’s oldest known marsupial, recovered from 55-million-year-old deposits in Murgon, southern Queensland. Comparative and evolutionary analyses indicate that these fossils belong to a newly recognized branch of the marsupial family tree, the Keeunamorphia, one of the earliest offshoots of Australidelphia: a group that includes all living Australian marsupials. The Keeunamorphia lineage appears to have originated in Gondwana more than 50 million years ago and persisted in Australia for over 35 million years before disappearing in the Middle Miocene. Some features also link keeunamorphians to early South American marsupials, suggesting ancient connections between continents that once made up Gondwana. These fossils reveal that Australia was once home to an ancient marsupial lineage that connects the continent’s earliest forms with the later-evolving carnivorous and omnivorous groups, including dasyuromorphians and bandicoots, shedding new light on the deep origins and diversification of Australia’s marsupial fauna.
Introduction
Tribosphenic molars represent the likely plesiomorphic dental condition for the mammalian clade Theria, which comprises Eutheria (placentals and their stem relatives) and Metatheria (marsupials and their stem relatives) (Szalay, Reference Szalay and Archer1982; Rich et al., Reference Rich, Vickers-Rich, Constantine, Flannery, Kool and van Klaveren1997; Luo et al., Reference Luo, Cifelli and Kielan-Jaworowska2001; Kielan-Jaworowska et al., Reference Kielan-Jaworowska, Cifelli and Luo2004; Voss and Jansa, Reference Voss and Jansa2009; Beck et al., Reference Beck, Voss and Jansa2022; Beck, Reference Beck, Cáceres and Dickman2023). In Australia, the only modern/extant marsupials retaining a relatively unmodified tribosphenic dentition are dasyurids (Archer, Reference Archer1976a; Beck et al., Reference Beck, Voss and Jansa2022). However, the fossils emerging from sites such as the Riversleigh World Heritage Area reveal a far more complex history of tribosphenic metatherians in Australia (Archer and Hand, Reference Archer, Hand, Archer and Clayton1984; Long et al., Reference Long, Archer, Flannery and Hand2002; Black et al., Reference Black, Archer, Hand, Godthelp and Talent2012).
The Oligo-Miocene deposits of the Riversleigh World Heritage Area contain the richest assemblage of fossil mammals known in Australia and are the source of most extinct tribosphenic marsupial species described to date on that continent. Prior to this study, all of the fossil tribosphenic marsupials from Riversleigh have been attributed to the orders Dasyuromorphia, Peramelemorphia, or Notoryctemorphia (Archer, Reference Archer1999; Long et al., Reference Long, Archer, Flannery and Hand2002; Archer et al., Reference Archer, Arena, Bassarova, Beck and Brewer2006; Black et al., Reference Black, Archer, Hand, Godthelp and Talent2012). These marsupials occupy a wide variety of dietary niches, including medium-sized to large (1–20 kg) faunivores (Muirhead and Archer, Reference Muirhead and Archer1990; Muirhead, Reference Muirhead1992, Reference Muirhead1997; Muirhead and Gillespie, Reference Muirhead and Gillespie1995; Wroe, Reference Wroe1996, Reference Wroe2001a; Muirhead and Wroe, Reference Muirhead and Wroe1998; Wroe and Musser, Reference Wroe and Musser2001; Archer et al., Reference Archer, Christmas, Hand, Black and Creaser2016a; Rovinsky et al., Reference Rovinsky, Evans and Adams2019; Churchill et al., Reference Churchill, Archer and Hand2024), small (0.05–1 kg) insectivores (Wroe, Reference Wroe1997a, Reference Wroe1998, Reference Wroe2001b; Arena et al., Reference Arena, Archer, Godthelp, Hand and Hocknull2011; Archer et al., Reference Archer, Hand, Black, Beck, Arena, Wilson, Kealy and Hung2016b; Churchill et al., Reference Churchill, Archer, Hand, Myers, Gillespie and Beck2023, Reference Churchill, Archer and Hand2025a, Reference Churchill, Archer and Handb), and small- to medium-sized (0.05–2 kg) omnivores (Muirhead and Filan, Reference Muirhead and Filan1995; Travouillon et al., Reference Travouillon, Gurovich, Beck and Muirhead2010, Reference Travouillon, Gurovich, Archer, Hand and Muirhead2013, Reference Travouillon, Hand, Archer and Black2014, Reference Travouillon, Archer, Hand and Muirhead2015; Gurovich et al., Reference Gurovich, Travouillon, Beck, Muirhead and Archer2014).
However, other Australian Oligo-Miocene deposits include tribosphenic taxa that do not comfortably fit within any currently recognized marsupial orders. Keeuna woodburnei and Ankotarinja tirarensis were described by Archer (Reference Archer1976b) based on partial dental material known from the late Oligocene deposits of the Etadunna Formation in South Australia. At the time, Archer (Reference Archer1976b) described these taxa as “structural ancestors” to dasyurids based on their resemblance to modern New Guinean dasyurids (e.g., species of Neophascogale and Phascolosorex). However, Archer (Reference Archer1976b) also noted resemblances, especially in the more plesiomorphic A. tirarensis, to unspecialized tribosphenic metatherians from the early Paleogene Itaboraí Fauna of Brazil (e.g., species of Coona, Marmosopsis, Mirandatherium, Monodelphis, Derorhynchus, Ischyrodidelphis, Didelphopsis, and Minusculodelphis), raising the possibility that they may also share a specific relationship with South American metatherians (Marshall, Reference Marshall and Archer1987; Goin, Reference Goin, Jones, Dickman and Archer2003; Carneiro and Oliveira Reference Carneiro, Oliveira, Cáceres and Dickman2023; Bergqvist et al., Reference Bergqvist, Carneiro, Zanesco, Otávio Resende Castro and Oliveira2024).
Later studies supported assignment of K. woodburnei and A. tirarensis to the superordinal marsupial clade Australidelphia, which includes the South American order Microbiotheria and the four extant Australian marsupial orders (Wroe, Reference Wroe1997b; Wroe et al., Reference Wroe, Ebach, Ahyong, Muizon and Muirhead2000; Beck et al., Reference Beck, Voss and Jansa2022; Beck, Reference Beck, Cáceres and Dickman2023). The assignment of K. woodburnei and A. tirarensis to Australidelphia was based on the close resemblance (especially the presence of a central cusp on the upper molars) between those taxa and Djarthia murgonensis Godthelp et al., Reference Godthelp, Wroe and Archer1999, a plesiomorphic australidelphian from the early Eocene Tingamarra Local Fauna of northeastern Queensland (Godthelp et al., Reference Godthelp, Wroe and Archer1999; Beck et al., Reference Beck, Godthelp, Weisbecker, Archer and Hand2008b). However, the precise relationships of K. woodburnei and A. tirarensis to other australidelphians remains uncertain (Godthelp et al., Reference Godthelp, Wroe and Archer1999; Kealy and Beck, Reference Kealy and Beck2017; Churchill et al., Reference Churchill, Archer, Hand, Myers, Gillespie and Beck2023, Reference Churchill, Archer and Hand2024, Reference Churchill, Archer and Hand2025a, Reference Churchill, Archer and Handb).
The earliest record of tribosphenic metatherians in Australia is from the early Eocene (ca. 54.6 Ma) deposits of the Tingamarra Local Fauna at Murgon in southern Queensland (Godthelp et al., Reference Godthelp, Archer, Cifelli, Hand and Gilkeson1992). Djarthia murgonensis, which was described based on upper dentitions retaining P3 and M1–4 and lower dentitions retaining p2–3 and m1–4, is the most complete and well-studied mammalian taxon from Murgon (Godthelp et al., Reference Godthelp, Wroe and Archer1999; Beck et al., Reference Beck, Godthelp, Weisbecker, Archer and Hand2008b). Since its description, D. murgonensis has been widely considered the most plesiomorphic known member of the australidelphian marsupial radiation (Godthelp et al., Reference Godthelp, Wroe and Archer1999; Beck, Reference Beck2008, Reference Beck2012; Beck et al., Reference Beck, Godthelp, Weisbecker, Archer and Hand2008b). The shared presence of a ‘central cusp’ on the ectoloph of the upper molars of D. murgonensis, Keeuna woodburnei, and Ankotarinja tirarensis to the exclusion of other australidelphians led to these taxa being recovered within a clade that is sister to the rest of the australidelphian radiation in several later phylogenetic analyses (Kealy and Beck, Reference Kealy and Beck2017; Churchill et al., Reference Churchill, Archer, Hand, Myers, Gillespie and Beck2023, Reference Churchill, Archer and Hand2024, Reference Churchill, Archer and Hand2025a, Reference Churchill, Archer and Handb). However, until now, this clade has remained unnamed and has not been the focus of detailed study.
The Itaboraí Formation in Brazil is a limestone deposit previously estimated to be 53–50 million years old (Woodburne et al., Reference Woodburne, Goin, Raigemborn, Heizler, Gelfo and Oliveira2014; Bergqvist et al., Reference Bergqvist, Carneiro, Zanesco, Otávio Resende Castro and Oliveira2024), and known for its diverse fossil record, including multiple lineages of metatherians (Marshall, Reference Marshall and Archer1987; Beck, Reference Beck2017b; Carneiro and Oliveira Reference Carneiro, Oliveira, Cáceres and Dickman2023; Carneiro et al., Reference Carneiro, Bampi, Rangel, Guimaraes and Oliveira2024a, Reference Carneiro, Goin, Bampi, Silva, Rangel, Guimaraes and Areasb, Reference Carneiro, Oliveira, Silva, Bampi and Areasc, Reference Carneiro, Zanesco, Bergqvist, Goin, Guedes, Agnes and Oliveirad). However, a recent study revisiting the age of the Itaboraí Basin suggests different ages of the fauna-bearing infilled fissures, the oldest (MCT-2) being late Paleocene to earliest Eocene, and the youngest (MCT-1) being early Eocene in age (Bergqvist et al., Reference Bergqvist, Carneiro, Zanesco, Otávio Resende Castro and Oliveira2024; Carneiro et al., Reference Carneiro, Bergqvist, Castro and Zanesco2026). Of the fauna described from the Itaboraí basin (MCT-1 and MCT-2), Carolopaulacoutoia itaboraiensis Paula Couto, Reference Paula Couto1970, Itaboraidelphys camposi Marshall and Muizon, Reference Marshall and Muizon1984, and species of Didelphopsis, which have been referred to the family Sternbergiidae, have been proposed as close relatives of Djarthia murgonensis, Keeuna woodburnei, and Ankotarinja tirarensis by some authors (Oliveira, Reference Oliveira1998; Goin, Reference Goin, Jones, Dickman and Archer2003). This is based on the shared possession of a central cusp on the upper molars, a featured only present among metatherians in these taxa and some herpetotheriids such as the putative herpetotheriid Maastrichtidelphys meurismeti Martin et al., Reference Martin, Case, Jagt, Schulp and Mulder2005, and some unequivocal herpetotheriids such as species of Peratherium (Archer, Reference Archer1976b; Oliveira, Reference Oliveira1998; Williamson et al., Reference Williamson, Brusatte, Carr, Weil and Standhardt2012, Reference Williamson, Brusatte and Wilson2014; Carneiro and Oliveira Reference Carneiro, Oliveira, Cáceres and Dickman2023). However, more recent studies have not supported a clade relationship between D. murgonensis, K. woodburnei, A. tirarensis and South American sternbergiids (Oliveira and Goin, Reference Oliveira and Goin2011; Carneiro et al., Reference Carneiro, Goin, Bampi, Silva, Rangel, Guimaraes and Areas2024b).
This study describes three new tribosphenic metatherian species from Riversleigh’s Early Miocene deposits that share a combination of derived and plesiomorphic dental features, including a central cusp on the ectoloph of the upper molars, with Keeuna woodburnei, Ankotarinja tirarensis, and Djarthia murgonensis. The clade comprising these six species cannot be referred to any of the currently recognized Australian orders, including Dasyuromorphia, although they are dentally similar to relatively plesiomorphic members of that order. The phylogenetic and morphometric results of this study lead us to the conclusion that establishment of a new marsupial order is appropriate to accommodate these taxa.
Methods
Materials
All specimens referred to in this study have been registered with Queensland Museum catalogue numbers, the institution where they will be permanently housed. While under study, they are currently held in the vertebrate paleontological research collections of the University of New South Wales, Sydney.
Data collection
All specimens were measured using a WILD MMS235 measuring apparatus attached to a Leica WILD M3B microscope. Molar measurements were recorded following Churchill et al. (Reference Churchill, Archer and Hand2025a) and include maximum length, anterior length, posterior length, anterior width, and posterior width for upper and lower molars, with distance from stB (stylar cusp B) to the paracone, distance from stD (stylar cusp D) to the metacone, length of the postmetacrista, and maximum width of the talon also recorded for the upper molars. All specimens were temporarily coated in magnesium oxide powder to improve the visibility of crown morphology for photographic purposes.
The dentitions of the new taxa were compared to those of other tribosphenic marsupials including representative dasyuromorphians, peramelemorphians, didelphimorphians, microbiotherians, and notoryctemorphians because these are the clades that have modern taxa that share the most similarities with the new taxa described here. The new taxa were also compared to fossil tribosphenic metatherians from Paleogene deposits in North and South America and Europe. All specimens that have been used as part of the morphological comparisons are listed in Supplementary File 1.
Body-mass estimates
The body masses of the new taxa were estimated using the molar regressions of Myers (Reference Myers2001), based on the ‘dasyuromorphian-only’ dataset. The limitations noted by Rovinsky et al. (Reference Rovinsky, Evans, Martin and Adams2020) in using molar regressions to estimate body mass for large metatherians do not apply to these relatively diminutive taxa.
Phylogenetic analyses
Phylogenetic relationships were investigated using maximum parsimony (MP) analyses of morphological data, and Bayesian total-evidence analyses of combined morphological and molecular datasets. The morphological dataset comprised 219 craniodental and postcranial characters (58 ordered; 161 unordered) modified from previous studies of dasyuromorphian and marsupial relationships (Wroe et al., Reference Wroe, Ebach, Ahyong, Muizon and Muirhead2000; Kealy and Beck, Reference Kealy and Beck2017; Rovinsky et al., Reference Rovinsky, Evans and Adams2019; Churchill et al., Reference Churchill, Archer, Hand, Myers, Gillespie and Beck2023, Reference Churchill, Archer and Hand2024, Reference Churchill, Archer and Hand2025a, Reference Churchill, Archer and Handb). Eight additional craniodental characters were added based on newly identified features suitable for phylogenetic scoring (see Character Analysis section in Supplementary File 3). The primary matrix included 67 taxa representing dasyuromorphians and other marsupials, with three Paleocene Tiupampa taxa (Mayulestes ferox Muizon, Reference Muizon1994, Pucadelphys andinus Marshall and Muizon, Reference Marshall and Muizon1988, and Andinodelphys cochabambensis) designated as outgroups. The new taxon Phantasmodon travouilloni n. gen. n. sp. was included and scored for 64% of craniodental characters. A second version of the matrix (71 taxa) additionally incorporated early Eocene sternbergiids Carolopaulacoutoia itaboraiensis, Itaboraidelphys camposi, Didelphopsis spp., and the herpetotheriid Peratherium constans Teilhard de Chardin, Reference Teilhard de Chardin1927, to test alternative placements of several Australian Paleogene taxa.
Maximum parsimony analyses were conducted in PAUP* using heuristic searches with tree-bisection–reconnection branch swapping and a molecular scaffold constraining relationships among extant taxa. Clade support was assessed using bootstrap analyses with 250 replicates. Bayesian tip-and-node dated total-evidence analyses were performed in MrBayes using the morphological dataset combined with a 16.4 kb (kilobases) nuclear and mitochondrial sequence alignment. Morphological data were analyzed under a single Mk model and molecular data under a single GTR+I+G model, with divergence times estimated using separate independent clock models for the morphological and molecular partitions, and fossil tip and node calibrations. Analyses were run for 100 million generations and summarized using a 50% majority-rule consensus tree. Full methodological details are provided in Supplementary File 2.
Anatomical abbreviations
a, anterior; ac, anterior cingulid; acm, anterior cingulum; ar, ascending ramus; b, buccal; cc, central cusp; co, cristid obliqua; end, entoconid; hd, hypoconid; hld, hypoconulid; l, lingual; m, molar; me, metacone; mecl, metaconule; mecd, metacristid; med, metaconid; mef, mental foramen; mf, masseteric fossa; ms, mandibular symphysis; mst, metastylar; p, premolar; pa, paracone; pacd, paracristid; pad, paraconid; pc, posterior cingulid; phcd, posthypocristid; precd, preentocristid; prd, protoconid; prpc, preparacrista; prmc, premetacrista; pspc, postparacrista; psmc, postmetacrista; stA, stylar cusp A; stB, stylar cusp B; stC, stylar cusp C; stD, stylar cusp D; stE, stylar cusp E. This study follows the morphological terminology of Archer (Reference Archer1976a) for the molar dentition and the thegotic terminology of Every (Reference Every1970) and the cheektooth serial homology of Flower (Reference Flower1867) (see Beck et al., Reference Beck, Voss and Jansa2022: table 12 for alternative homologies). Stylar cusp homology follows Clemens (Reference Clemens1966).
Repositories and institutional abbreviations
AR, University of New South Wales paleontological collections; QM F, Queensland Museum paleontological collections.
Systematic paleontology
Class Mammalia Linnaeus, Reference Linnaeus1758
Subclass Theria Parker and Haswell, Reference Parker and Haswell1897
Metatheria Huxley, Reference Huxley1880
Marsupialiformes Rowe, Reference Rowe1988, Vullo et al., Reference Vullo, Gheerbrant, Muizon and Néraudeau2009, sensu Beck, Reference Beck and Ebach2017a
Marsupialia Illiger, Reference Illiger1811, sensu Beck and Lee, Reference Beck and Lee2014
Order Keeunamorphia new order
Included species
Keeuna woodburnei Archer, Reference Archer1976b, Ankotarinja tirarensis Archer, Reference Archer1976b, Djarthia murgonensis Godthelp et al., Reference Godthelp, Wroe and Archer1999, Phantasmodon travouilloni n. gen. n. sp., P. minuferox n. gen. n. sp., and Keeunidae n. fam. gen. and sp. indet. (this study).
Phylogenetic definition
The most inclusive clade including Keeuna woodburnei, but excluding Didelphis marsupialis Linnaeus, Reference Linnaeus1758, Caenolestes fuliginosus (Tomes, Reference Tomes1863), Dasyurus viverrinus (Shaw, Reference Shaw1800), Perameles nasuta Geoffroy, Reference Geoffroy Saint-Hilaire1804, Notoryctes typhlops Stirling, Reference Stirling1889, Phalanger orientalis (Pallas, Reference Pallas1766), and Dromiciops gliroides Thomas, Reference Thomas1894.
Diagnosis
All members of Keeunamorphia n. ord., including the taxa described here, possess the following combination of dental features to the exclusion of all other metatherians: upper molars and lower molars tribosphenic in overall morphology; central cusp present on the stylar shelf situated slightly buccal to the centrocrista and lingual to stC and stD on all upper molars; narrow anterior cingulum present on the anterior face of all upper molars between stA and preprotocrista; preparacristae oriented perpendicular to the long axis of the tooth on all molars; preparacristae contact the anterior or lingual base of stB on all molars; metacone higher crowned than paracone on M1–3; centrocrista strongly V-shaped on all upper molars; ectolophs narrow as a result of buccal intrusion by the centrocrista; metaconules large relative to presumed plesiomorphic condition in tribosphenic metatherians on all upper molars; metacone subequal in height to paracone on M4; posterior cingulid present on m1–3; talonid wider than trigonid on m1–3; cristid obliqua terminates at the base of the trigonid buccal to the metacristid carnassial notch on m1–4; entoconid large and approaching almost subequal in height to paraconid on m1–3; preentocristid well developed on m1–3; hypoconulid reduced on all molars; hypoconulid much lower crowned than the entoconid on all molars; three cusps on m4 talonid; entoconid large on m4; posthypocristid present on m4.
Keeunamorphians differ from dasyuromorphians in the following combination of dental features: presence of a central cusp on all upper molars; ectolophs narrow on M1–3; internal angle of protocone obtuse on M1–3; metastylar corner of tooth not elongated on M2–3; metaconules large (rather than reduced or absent) on all upper molars; metacone subequal in height to paracone on M4 (rather than highly reduced or absent); talonid wider than trigonid on m3; entoconids large on all molars.
Keeunamorphians differ from peramelemorphians in the following combination of dental features: presence of a central cusp; preparacrista contacts stB (not stA) on all molars; centrocrista complete and V-shaped on M1–3 (except in Galadi grandis Travouillon et al., Reference Travouillon, Gurovich, Beck and Muirhead2010, and species of Yarala, which have complete and V-shaped centrocristae); preparacrista on M1 oriented perpendicular to the long axis of the tooth; stC present on M1–3; posterior cingulids present on m1–3; cristid obliqua terminates at the base of the trigonid buccal to the metacristid carnassial notch (rather than lingual); trigonids markedly higher crowned than talonids (except in species of Yarala, which share this condition); hypoconulid notch present on m2–4; three cusps on the m4 talonid.
Keeunamorphians differ from South American sternbergiids (which also possess a central cusp on the upper molars: Carolopaulacoutoia itaboraiensis, Itaboraidelphys camposi, Didelphopsis spp.) in the following dental features: preparacrista contacts the lingual base of stB on all molars; M2–3 not wider than long; stD larger than stB on M1–2; metaconule large and appressed to the lingual base of the metacone on M4; m4 talonid more transversely reduced.
Keeunamorphians differ from didelphids in the following combination of dental features: presence of central cusp on all upper molars; preparacrista contacts posterolingual base of stB, rather than between stB and stA; absence of postmetacrista on M4; metaconule large and appressed to the lingual base of the metacone on M4; posterior cingulid present on m1–3 (except in Philander pebas Voss, Díaz-Nieto, and Jansa, Reference Voss, Díaz-Nieto and Jansa2018, and the ‘Rapposa’ group of Marmosa, which also exhibit this condition; Voss et al., Reference Voss, Díaz-Nieto and Jansa2018, Reference Voss, Giarla, Díaz-Nieto and Jansa2020); m4 talonid reduced relative to the trigonid; protoconids markedly taller than metaconids and paraconids on all molars.
Keeunamorphians differ from the plesiomorphic South American metatherians from Tiupampa (clade Pucadelphyoidea; see Muizon and Ladevèze, Reference Muizon and Ladevèze2022: fig. 14) Andinodelphys cochabambensis, Pucadelphys andinus, and Incadelphys antiquus Marshall and Muizon, Reference Marshall and Muizon1988, in the following combination of dental features: central cusp present on all upper molars; upper molars not markedly wider than long; preparacrista contacts the lingual base, rather than the anterior base, of stB; metacone markedly larger than paracone on M3 (approximately subequal in size on M3 in pucadelphyoids); metacone subequal in height to paracone on M4; metaconules large and appressed to lingual base of metacone on M4; talons broader on all molars with an internal angle of ~90–110°, rather than ~60°.
Publication LSID
urn:lsid:zoobank.org:pub:A2699457-5B2C-4F0C-B8B6-DD41401C3A17
Keeunamorphia new order
Family incertae sedis
Included species
Ankotarinja tirarensis Archer, Reference Archer1976b, and Djarthia murgonensis Godthelp et al., Reference Godthelp, Wroe and Archer1999.
Remarks
Djarthia murgonensis and Ankotarinja tirarensis differ from other keeunamorphians in their very small body size (~10–30 g) and the presence of transversely oriented metacristids on all lower molars. In other members of the order, the metacristid is more oblique (~15–30°) relative to the long axis of the tooth. Although these similarities suggest a potentially distinctive lineage, phylogenetic analyses did not recover D. murgonensis and A. tirarensis as sister taxa: parsimony analyses placed them as paraphyletic (Figs. 5 and 6), whereas Bayesian analyses recovered them in separate parts of the tree (Figs. 7 and 8). Given the limited material and small number of scorable characters, we consider erection of a distinct family to be premature, and both taxa are here regarded as Keeunamorphia n. ord. incertae sedis.
Family Keeunidae new family
Type genus
Keeuna Archer, Reference Archer1976b.
Diagnosis
Members of Keeunidae n. fam. including Phantasmodon travouilloni n. gen. n. sp., P. minuferox n. gen. n. sp., Keeunidae n. fam. gen. and sp. indet., and Keeuna woodburnei differ from other keeunamorphians in the following combination of dental features: stylar cups D large on M3; metacristids oblique on all lower molars.
Type species
Phantasmodon travouilloni n. gen. n. sp.
Diagnosis
Species of Phantasmodon n. gen. differ from other keeunamorphians in possessing obliquely oriented metacristids on m1–2. Generic differences of the remaining lower molar and upper molar dentition between species of Phantasmodon n. gen. as represented by P. travouilloni n. gen. n. sp. and other keeunamorphians are listed in the species diagnosis of P. travouilloni.
Etymology
The genus name Phantasmodon is derived from phantasma (Greek), meaning phantom/ghost and odon/odontos (Greek) meaning tooth: phantom tooth.
Remarks
Species of Phantasmodon n. gen. differ sufficiently from Keeuna, Ankotarinja , and Djarthia to warrant placement in a distinct genus. The genus is characterized by a distinctive reorganization of both the stylar shelf and the shearing crests of the upper and lower molars. Most of these features are currently documented only in Phantasmodon travouilloni n. gen. n. sp., the only species known from a complete upper and lower molar dentition and therefore inferred to represent the generic condition pending discovery of additional material for P. minuferox n. gen. n. sp., which is presently known only from m1–2. In P. travouilloni, species of Phantasmodon n. gen. are distinguished from other keeunamorphians by the combination of a larger internal angle of the M3 centrocrista, a consistently higher-crowned stD relative to stB, the polymorphic presence of a short posterior cingulum on M1–3, oblique metacristids and posthypocristids, and a central cusp that is divided into two smaller cusps. This suite of characters differentiates Phantasmodon n. gen. from Djarthia murgonensis, Keeuna woodburnei, and Ankotarinja tirarensis, and indicates a departure from the more transverse shearing arrangement and simpler central cusp morphology that characterize other members of Keeunamorphia n. ord.
Phantasmodon travouilloni new species
Figures 1–3, 4.1–4.7, 9.1–9.3, 10.1
Phantasmodon travouilloni n. gen. n. sp. holotype right maxilla retaining M3 and M4 (QM F61741). Specimen shown in (1, 2) occlusal view as a stereo pair with labeled cusp morphology; (3) oblique buccal view; (4) buccal view; (5) oblique lingual view; (6) lingual view. Abbreviations: acm, anterior cingulum; cc, central cusp; mcl, metaconule; me, metacone; mst, metastylar; pa, paracone; pr, protocone; prmc, premetacrista; prpc, preparacrista; prprc, preprotocrista; psmc, postmetacrista; pspc, postparacrista; psprc, postprotocrista; stA, stylar cusp A; stB, stylar cusp B; stC, stylar cusp C; stD, stylar cusp D; stE, stylar cusp E. Scale bar represents 2 mm.

Figure 1. Long description
Panel 1 (top left) displays an occlusal view of the right maxilla with M3 and M4, annotated with dental features: anterior cingulum (acm), central cusp (cc), metaconule (mcl), metacone (me), metastylar (mst), paracone (pa), protocone (pr), premetacrista (prmc), preparacrista (prpc), preprotocrista (prprc), postmetacrista (psmc), postparacrista (pspc), postprotocrista (psprc), and stylar cusps A to E (stA, stB, stC, stD, stE). Panel 2 (top right) is a stereo pair occlusal view without labels. Panel 3 (middle left) shows an oblique buccal view, highlighting the lateral aspect of the teeth. Panel 4 (middle right) presents a direct buccal view. Panel 5 (bottom left) is an oblique lingual view, and panel 6 (bottom right) is a direct lingual view. The scale bar at the bottom left represents 2 millimeters.
Phantasmodon travouilloni n. gen. n. sp. paratype isolated upper molar specimens. (1) Right M1 (AR22170); (2–6) left M1s (AR22926, AR22173, AR22927, AR22171, and AR22172); (7) right M2 (AR22187); (8) left M2 (QM F30872); (9–11) right M3s (AR22200, AR22928, and AR22188); (12) left M3 (AR22929). All specimens shown as an occlusal stereo pair (top), in buccal view (bottom left), and in lingual view (bottom right). Scale bar represents 2 mm.

Figure 2. Long description
From top left to bottom right, the grid contains twelve numbered molar specimens. The first six columns in the top two rows are labeled M1, showing right M1 (1) and left M1s (2 to 6). The third row contains right M2 (7) and left M2 (8). The last two rows show right M3s (9 to 11) and left M3 (12). Each specimen is presented in three views: the uppermost image is the occlusal (chewing surface), the lower left is the buccal (cheek side), and the lower right is the lingual (tongue side). The scale bar below specimen 5 indicates 2 millimeters. The teeth display variation in cusp shape, crown height, and root structure, with some showing wear or discoloration. All specimens are isolated and not attached to jaw fragments.
Phantasmodon travouilloni n. gen. n. sp. paratype left dentary (QM F61742). (1) specimen shown in occlusal view as a stereo pair; (2) dentition shown in occlusal view as a stereo pair with labeled cusp morphology; (3) specimen shown in lingual view; (4) specimen shown in buccal view; (5) dentition shown in lingual view; (6) dentition shown in buccal view. Abbreviations: ac, anterior cingulid; ap, angular process; ar, ascending ramus; co, cristid obliqua; end, entoconid; hd, hypoconid; hld, hypoconulid; maf, mandibular foramen; mecd, metacristid; med, metaconid; mef, mental foramen; mf, masseteric fossa; pacd, paracristid; pad, paraconid; pc, posterior cingulid; phcd, posthypocristid; prd, protoconid; precd, preentocristid. Scale bar on the left represents 5 mm. Scale bar on the right represents 2 mm.

Figure 3. Long description
Panel 1 at the top left displays a stereo pair of the left dentary in occlusal view, showing the full tooth row and jaw outline. Panel 2 at the top right presents a stereo pair of the dentition in occlusal view, with black lines labeling ac (anterior cingulid), pacd (paracristid), prd (protoconid), co (cristid obliqua), hd (hypoconid), pc (posterior cingulid), phcd (posthypocristid), pad (paraconid), mecd (metacristid), med (metaconid), precd (preentocristid), end (entoconid), and hld (hypoconulid). The left scale bar is 5 mm, the right is 2 mm. Panel 3 at the middle left shows the specimen in lingual view, with ap (angular process) and maf (mandibular foramen) labeled. Panel 4 at the middle right shows the specimen in buccal view, with ar (ascending ramus), mf (masseteric fossa), and mef (mental foramen) labeled. Panel 5 at the bottom left displays the dentition in lingual view, and panel 6 at the bottom right shows the dentition in buccal view. All panels highlight the spatial arrangement and morphology of the jaw and teeth.
Phantasmodon travouilloni n. gen. n. sp. paratype isolated lower molar specimens (1–7). (1, 2) Left m1s (AR22182 and AR22208); (3) right dentary retaining m2 with alveoli for m3 and m4 (AR22201); (4) right m2 (AR22193); (5) left m2 (AR22209). (6, 7) left m3s (AR22207 and AR22181); (8) Keeunidae n. fam. gen. and sp. indet. right m1 (QM F61740); (9) Phantasmodon minuferox n. gen. n. sp. holotype left m1 (QM F61739) and (10) paratype left m2 (AR22195). All specimens shown as an occlusal stereo pair (top), in buccal view (middle), and in lingual view (bottom). Scale bar represents 4 mm.

Figure 4. Long description
Ten panels are arranged in a grid, each labeled 1 to 10. Each panel displays a fossil lower molar specimen in three views: the top row shows the occlusal (chewing surface), the middle row shows the buccal (cheek side), and the bottom row shows the lingual (tongue side). Panels 1 and 2 show left m1s (AR22182 and AR22208); panel 3 shows a right dentary with m2 and alveoli for m3 and m4 (AR22201); panel 4 shows right m2 (AR22193); panel 5 shows left m2 (AR22209); panels 6 and 7 show left m3s (AR22207 and AR22181); panel 8 shows Keeunidae indeterminate right m1 (Q M F61740); panel 9 shows Phantasmodon minuferox holotype left m1 (Q M F61739); panel 10 shows paratype left m2 (AR22195). A black scale bar below panel 3 indicates 4 millimeters. The teeth vary in cusp pattern and wear, with some embedded in jaw fragments and others isolated.
50% majority rule consensus of 279,180 most parsimonious trees (length = 1120; CI excluding autapomorphies = 0.2607; HI excluding autapomorphies = 0.739; RI = 0.524; RC = 0.137) resulting from maximum parsimony analysis of the 218-character morphological matrix, with relationships among modern taxa constrained to match molecular studies (Mitchell et al., Reference Mitchell, Pratt, Watson, Gibb, Llamas, Kasper, Edson, Hopwood, Male and Armstrong2014; Westerman et al., Reference Westerman, Krajewski, Kear, Meehan, Meredith, Emerling and Springer2015). Node values represent 1000 replicate bootstrap values > 50%. Taxa marked with italicized daggers (†) represent extinct species. Clades representing recognized families of Dasyuromorphia are colored: Thylacinidae in red, Myrmecobiidae in yellow; Dasyuridae in blue; Malleodectidae in pink. The clade representing the new metatherian order Keeunamorphia n. ord. is colored in green. Taxon label of new taxon shown in bold. Taxa listed on Figure 5 not otherwise noted in the text: Antechinomys laniger (Gould, Reference Gould1856); Antechinus flavipes (Waterhouse, Reference Waterhouse1838); Caluromys philander (Linnaeus, Reference Linnaeus1758); Dasycercus cristicauda (Krefft, Reference Krefft1867); Dasykaluta rosamondae (Ride, Reference Ride1964); Dasyuroides achilpatna Archer, Reference Archer and Archer1982; Dasyuroides byrnei Spencer, Reference Spencer1896; Dasyurus albopunctatus Schlegel, Reference Schlegel1880; Dasyurus dunmalli Bartholomai, Reference Bartholomai1971; Dasyurus hallucatus Gould, Reference Gould1842; Dasyurus maculatus (Kerr, Reference Kerr1792); Dasyurus spartacus Van Dyck, Reference Van Dyck1988; Dasyurus viverrinus (Shaw, Reference Shaw1800); Echymipera kalubu (Fischer, Reference Fischer1829); Malleodectes mirabilis Arena et al., Reference Arena, Archer, Godthelp, Hand and Hocknull2011; Malleodectes wentworthi Churchill et al., Reference Churchill, Archer, Hand, Myers, Gillespie and Beck2023; Marmosa murina (Linnaeus, Reference Linnaeus1758); Murexechinus melanurus (Thomas, Reference Thomas and Loria1899); Murexia longicaudata (Schlegel, Reference Schlegel1866); Myoictis melas (Müller, Reference Müller, Temminck and van der Hoek1840); Neophascogale lorentzii (Jentink, Reference Jentink1911); Ningaui yvonnae Kitchener, Stoddart, and Henry, Reference Kitchener, Stoddart and Henry1983; Paramurexia rothschildi (Tate, Reference Tate1938); Parantechinus apicalis (Gray, Reference Gray1842); Phascogale tapoatafa (Meyer, Reference Meyer1793); Phascolosorex dorsalis (Peters and Doria, Reference Peters and Doria1876); Phascomurexia naso (Jentink, Reference Jentink1911); Planigale maculata (Gould, Reference Gould1851); Pseudantechinus bilarni (Johnson, Reference Johnson1954); Pseudantechinus macdonnellensis (Spencer, Reference Spencer1895); Sarcophilus harrisii Boitard, Reference Boitard1841; Sminthopsis crassicaudata (Gould, Reference Gould1844); Sminthopsis floravillensis Archer, Reference Archer and Archer1982; Sminthopsis murina (Waterhouse, Reference Waterhouse1838); Thylacinus cynocephalus (Harris, Reference Harris1808); Thylacinus megiriani Murray, Reference Murray1997; Thylacinus potens Woodburne, Reference Woodburne1967; Thylacinus yorkellus Yates, Reference Yates2015.

Figure 5. Long description
The diagram is a branching phylogenetic tree anchored at the top left with Australasian marsupial orders. The main trunk splits into Didelphimorphia, Keeunamorphia (in green), Microbiotheria, Peramelemorphia, and Dasyuromorphia. Dasyuromorphia divides into four colored clades: Thylacinidae (red), Myrmecobiidae (yellow), Dasyuridae (blue), and Malleodectidae (pink). Keeunamorphia is highlighted in green with the new taxon label in bold. Extinct taxa are marked with italicized daggers. Bootstrap values are shown at nodes, only values above 50 are displayed. Taxon names are listed at branch tips, with modern and extinct species interspersed. The tree structure reflects relationships constrained by molecular data, with each family’s clade distinctly colored and labeled at the right margin. The tree proceeds from broad groupings at the top left to specific taxa at the bottom right, with all major families and their representative species included.
50% majority rule consensus of 1430 most parsimonious trees (length = 1143; CI excluding autapomorphies = 0.255; HI excluding autapomorphies = 0.745; RI = 0.523; RC = 0.134) resulting from maximum parsimony analysis of the 218-character morphological matrix including the sternbergiids Carolopaulacoutoia itaboraiensis, Itaboraidelphys camposi, and Didelphopsis sp., and the herpetotheriid Peratherium constans. Relationships among modern taxa were constrained to match molecular studies (Mitchell et al., Reference Mitchell, Pratt, Watson, Gibb, Llamas, Kasper, Edson, Hopwood, Male and Armstrong2014; Westermann et al., Reference Westerman, Krajewski, Kear, Meehan, Meredith, Emerling and Springer2015). Node values represent 1000 replicate bootstrap values > 50%. Taxa marked with italicized daggers (†) represent extinct species. Clades representing recognized families of Dasyuromorphia are colored: Thylacinidae in red, Myrmecobiidae in yellow; Dasyuridae in blue; Malleodectidae in pink. The clade representing the new metatherian order Keeunamorphia n. ord. is colored in green. Members of the metatherian family Sternbergiidae are colored in gray. Taxon label of new taxon shown in bold.

Figure 6. Long description
Rooted at the top left, the cladogram branches rightward and downward. The main clades are Didelphimorphia, Microbiotheria, Keeunamorphia (green), Peramelemorphia, and Dasyuromorphia. Dasyuromorphia splits into four color-coded families: Thylacinidae (red), Myrmecobiidae (yellow), Dasyuridae (blue), and Malleodectidae (pink). Sternbergiidae members are in gray. Keeunamorphia is labeled in green, with the new taxon Phasmatodon travoolloni in bold. Bootstrap values above 50 percent are shown at nodes. Extinct taxa are marked with italicized daggers. Taxon names are listed at the branch tips, ordered from top to bottom: Didelphimorphia includes taxa such as Pucadelphys andinus and Marmosa murina; Microbiotheria includes Microbiotherium tehuelochqui; Sternbergiidae includes Carolopaulacoutoia itaboraiensis and Didelphopsis spp.; Keeunamorphia includes Tankotria tiraensis and Keeuna woodburnei; Peramelemorphia includes Perameles nasuta; Thylacinidae includes Thylacinus cynocephalus and Thylacinus potens; Myrmecobiidae includes Myrmecobius fasciatus; Malleodectidae includes Malleodectes mirabilis; Dasyuridae includes Dasyurus viverrinus and Sminthopsis crassicaudata. Clade relationships among modern taxa are constrained to match molecular studies. The tree structure reflects maximum parsimony analysis of a 218-character morphological matrix.
50% majority rule consensus tree from the dated total evidence analysis of 218 morphological characters matrix combined with 16.4 kb (kilobases) of mitochondrial and nuclear sequence data, with dating using the Bayesian tip-and-node ‘TipNodeCalib1’ calibration scheme of Kealy and Beck (Reference Kealy and Beck2017). The partitioning scheme for morphological and molecular data uses an IGR clock model. Branch lengths represent time in millions of years. Node bars represent divergence age ranges as 95% Higher Posterior Densities (HDPs). Node color represents clade support as Bayesian Posterior Probabilities (BPP). Recognized families of Dasyuromorphia are colored: Thylacinidae in red; Myrmecobiidae in yellow; Dasyuridae in blue; Malleodectidae in pink. The clade representing the new metatherian order Keeunamorphia n. ord. is colored in green. Members of the metatherian clade Sternbergiidae are colored in gray. Taxa marked with italicized daggers (†) represent extinct species. Taxon label of the new taxon is shown in bold.

Figure 7. Long description
Starting at the upper left, the tree root branches into Didelphimorphia, Microbiotheria, and Australidelphia. Didelphimorphia includes Pucladelyphis and Mayulestes. Microbiotheria contains Microbiotherium and Antarctorya. Australidelphia splits into Keeunamorphia (green), Peramelemorphia, and Dasyuromorphia. Keeunamorphia includes Keeuna woodburnei and Phantasmodon travoulloni, with the new taxon Phantasmodon travoulloni in bold. Dasyuromorphia divides into Thylacinidae (red), Myrmecobiidae (yellow), Dasyuridae (blue), and Malleodectidae (pink). Sternbergiidae is shown in gray. Extinct taxa are marked with italicized daggers. Branch lengths represent millions of years, with a horizontal axis below showing geological periods from the Cretaceous to the Pleistocene. Node bars indicate 95 percent higher posterior densities for divergence ages, and node color intensity reflects Bayesian posterior probabilities from 0.5 to 1.0. Family names are labeled in matching colors along the right margin.
50% majority rule consensus tree from the dated total evidence analysis of 218 morphological characters matrix combined with 16.4 kb (kilobases) of mitochondrial and nuclear sequence data, with dating using the Bayesian tip-and-node ‘TipNodeCalib1’ calibration scheme of Kealy and Beck (Reference Kealy and Beck2017) and taxa including sternbergiids Carolopaulacoutoia itaboraiensis, Itaboraidelphys camposi, and Didelphopsis sp., and the herpetotheriid Peratherium constans. The partitioning scheme for morphological and molecular data uses an IGR clock model. Branch lengths represent time in millions of years. Node bars represent divergence age ranges as 95% Higher Posterior Densities (HDPs). Node color represents clade support as Bayesian Posterior Probabilities (BPP). Recognized families of Dasyuromorphia are colored: Thylacinidae in red; Myrmecobiidae in yellow; Dasyuridae in blue; Malleodectidae in pink. The clade representing the new metatherian order Keeunamorphia n. ord. is colored in green. Members of the metatherian family Sternbergiidae are colored in gray. Taxa marked with italicized daggers (†) represent extinct species. Taxon label of the new taxon is shown in bold.

Figure 8. Long description
The diagram is a left-to-right branching phylogenetic tree with a horizontal time axis at the bottom spanning from 70 million years ago on the left to present on the right, divided into Mesozoic, Paleogene, and Neogene periods. The tree root at the upper left splits into major marsupial clades: Microbiotheria, Didelphimorphia, Peramelemorphia, Keeunamorphia (in green), and Dasyuromorphia. Dasyuromorphia is subdivided into four color-coded families: Thylacinidae in red, Myrmecobiidae in yellow, Dasyuridae in blue, and Malleodectidae in pink. Sternbergiidae members are in gray. Keeunamorphia is highlighted in green, with the new taxon ‘Hannamodon travouilloni’ in bold. Extinct taxa are marked with italicized daggers. Branch lengths represent millions of years, with node bars indicating 95 percent higher posterior density age ranges. Node color intensity reflects Bayesian posterior probability support, from black (high) to gray (low). The time scale below the tree is labeled with geological epochs and the K P g boundary. Taxon names are listed at branch tips, with extinct and extant species distributed across the tree.
Upper dentitions of all known keeunamorphian taxa (1–7) and the plesiomorphic dasyuromorphian Barinya wangala (8). (1–3) Phantasmodon travouilloni n. gen. n. sp. (1) holotype right maxilla retaining M3 and M4 (QM F61741); (2) isolated paratype right M2 (AR22187); (3) isolated paratype right M1 (AR22170); (4, 5) Keeuna woodburnei (4) paratype left maxilla retaining M2 and M3 with alveoli for M4 (QM F7333); (5) isolated holotype right M2 (SAM P18191), flipped horizontal in figure; (6) Djarthia murgonensis holotype right maxilla retaining M1–4 (QM F31458); (7) Ankotarinja tirarensis paratypes right M2 and M3 (QM F7332 and UCR13508); (8) Barinya wangala paratype right maxilla retaining M1–4 (QM F31418). Scale bar represents 2 mm.

Figure 9. Long description
Panel 1 (top-left) shows a right maxilla with two molars labeled M4 stC large, metastylar separate from metacone, and M4 metaconule large. Panel 2 (top row, center-left) displays an isolated right M2 with central cusp and premetacrista unreduced. Panel 3 (top row, center-right) presents an isolated right M1 with a central cusp. Panel 4 (second row, left) is a left maxilla with M2 and M3, showing M4 stC small and central cusp. Panel 5 (second row, right) is an isolated right M2, horizontally flipped, with central cusp. Panel 6 (third row, left) is a right maxilla with M1 to M4, labeled M4 metastylar separate from metacone, M4 metaconule large, M4 premetacrista unreduced, and central cusp. Panel 7 (third row, right) shows right M2 and M3 with central cusp. Panel 8 (bottom row) is a right maxilla with M1 to M4, labeled M4 stC absent, metastylar and metacone fused, M4 metaconule absent, and M4 premetacrista reduced. The scale bar at the bottom right represents 2 millimeters.
Lower dentitions of all known keeunamorphian taxa (1–7) and plesiomorphic dasyuromorphians Joculusium muizoni and Barinya wangala (8, 9). (1) Phantasmodon travouilloni n. gen. n. sp. paratype left dentary retaining m1–4 (QM F61742); (2) Phantasmodon minuferox n. gen. n. sp. holotype left m1 (QM F61739) and paratype left m2 (AR22195); (3) Keeunidae n. fam. gen. and sp. indet. right m1 (QM F61740; flipped horizontal in figure); (4) Keeuna woodburnei paratype left m2 or m3 (UCR15344); (5) Djarthia murgonensis paratype right dentary retaining p3, m1–3, and broken m4 (QM F31458; flipped horizontal in figure); (6) Djarthia murgonensis paratype left dentary retaining m2–4 (QM F31460); (7) Ankotarinja tirarensis holotype right dentary retaining p2–3 and m2–4 (SAM P18190; flipped horizontal in figure) and paratype left m1 (UCR15340); (8) Joculusium muizoni holotype left dentary retaining p2–3 and m1–4 (QM F36442); (9) Barinya wangala paratype right dentary retaining m1–4 (QM F41284; flipped horizontal in figure). Scale bar represents 2 mm.

Figure 10. Long description
From left to right, panel 1 shows a left dentary with four molars labeled for oblique m1 metacristid and m3 talonid wider than trigonid. Panel 2 displays two isolated left molars, m1 and m2, with m1 metacristid oblique. Panel 3 presents a right m1, horizontally flipped. Panel 4 shows a left m2 or m3. Panel 5 features a right dentary with p3, m1 to m3, and a broken m4, horizontally flipped. Panel 6 displays a left dentary with m2 to m4, labeled for transverse m1 metacristid and m3 talonid wider than trigonid. Panel 7 includes a right dentary with p2 to p3 and m2 to m4, horizontally flipped, and a left m1. Panel 8 shows a left dentary with p2 to p3 and m1 to m4. Panel 9 presents a right dentary with m1 to m4, horizontally flipped, labeled for oblique m1 metacristid and m3 talonid narrower than trigonid. The scale bar below panel 2 represents 2 millimeters.
Holotype
QM F61741 is a right maxillary fragment retaining M3–4 (Fig. 1).
Paratypes
AR22170 is an isolated right M1 (Fig. 2.1). AR22926–7 and AR22171–3 are isolated left M1s (Fig. 2.2–2.6). AR22187 is an isolated right M2 (Fig. 2.7). QM F30872 is an isolated left M2 (Fig. 2.8). AR22928, AR22188, AR22200 (Fig. 2.9–2.11), and AR22931 are isolated right M3s. AR22929 is an isolated left M3 (Fig. 2.12). QM F61742 is a left dentary retaining m1–4 (Fig. 3). AR22182 and AR22208 are isolated left m1s (Fig. 4.1 and 4.2). AR22201 is a broken right dentary retaining m2 and alveoli for m3 (Fig. 4.3). AR22193 is an isolated right m2 (Fig. 4.4). AR22209 is an isolated left m2 (Fig. 4.5). AR22207 and AR22181 are isolated left m3s (Fig. 4.6 and 4.7). Dental measurements of all specimens of Phantasmodon travouilloni are shown in Table 1.
Dental measurements of Phantasmodon travouilloni n. gen. n. sp. Tooth position followed by an “A” indicates measurements based on root alveoli. Abbreviations: L, length; AW, anterior width; PW, posterior width; AL, anterior length; PL, posterior length; PPC, preparacrista length; MD, metacone to stD; PM, postmetacrista length; TW, talon width; MRL, molar row length

Table 1. Long description
From the top row, the table columns are: Specimen, Tooth, L, A W, P W, A L, P L, P C, M D, P M, T W, M R L. The holotype right maxilla Q M F six one seven four one has M three and M four teeth with L values of two point five and two point zero, A W two point five and two point eight, P W three point one and two point zero, A L one point zero and missing, P L one point six and missing, P C one point one and one point two, M D one point two and missing, P M one point seven and zero point four, T W one point five and one point one. Paratype right M one A R two two one seven zero has M one with L two point six, A W two point zero, P W two point nine, A L one point two, P L one point five, P C zero point seven, M D zero point nine, P M one point five, T W one point three. Paratype left M one A R two two nine two six has M one with L two point seven, A W one point nine, P W two point eight, A L one point three, P L one point five, P C zero point six, M D zero point eight, P M one point four, T W one point three. Paratype left M one A R two two one seven three has M one with L three point zero, A W two point one, P W two point nine, A L one point three, P L one point seven, P C zero point six, M D one point zero, P M one point six, T W one point four. Paratype left M one A R two two nine two seven has M one with L two point seven, A W two point zero, P W two point eight, A L one point two, P L one point six, P C zero point seven, M D one point zero, P M one point six, T W one point three. Paratype left M one A R two two one seven one has M one with L two point eight, A W one point nine, P W two point eight, A L one point two, P L one point six, P C zero point seven, M D zero point nine, P M one point four, T W one point one. Paratype left M one A R two two one seven two has M one with L two point six, A W one point nine, P W two point seven, A L one point two, P L one point six, P C zero point five, M D zero point eight, P M one point four, T W one point four. Paratype right M two A R two two one eight seven has M two with L two point five, A W two point two, P W two point eight, A L one point two, P L one point five, P C zero point nine, M D one point zero, P M one point four, T W one point four. Paratype left M two Q M F three zero eight seven two has M two with L two point seven, A W two point four, P W two point eight, A L one point one, P L one point six, P C zero point nine, M D one point zero, P M one point four, T W one point five. Paratype right M three A R two two zero zero zero has M three with L two point seven, A W two point six, P W three point two, A L one point one, P L one point five, P C one point one, M D one point three, P M one point six, T W one point five. Paratype right M three A R two two nine two eight has M three with L two point five, A W two point four, P W three point zero, A L one point three, P L one point four, P C one point zero, M D one point one, P M one point five, T W one point three. Paratype right M three A R two two one eight eight has M three with L two point four, A W two point seven, P W three point one, A L zero point nine, P L one point five, P C zero point nine, M D one point one, P M one point six, T W one point four. Paratype right M three A R two two nine three one has M three with L two point six, A W two point four, P W two point seven, A L one point four, P L one point two, P C zero point nine, M D one point zero, P M one point six, T W one point four. Paratype left M three A R two two nine two nine has M three with L two point six, A W two point six, P W three point two, A L one point zero, P L one point six, P C one point one, M D one point one, P M one point seven, T W one point four. Paratype left dentary Q M F six one seven four two has m one to m four with L values two point six, two point four, two point four, two point six, A W one point two, one point five, one point five, one point five, P W one point four, one point six, one point six, one point two, A L one point five, one point six, one point four, one point three, P L zero point nine, zero point nine, one point zero, one point one, M R L nine point eight. Paratype left m one A R two two two zero eight has m one with L two point four, A W one point one, P W one point three, A L one point five, P L zero point nine. Paratype left m one A R two two one eight two has m one with L two point five, A W one point one, P W one point three, A L one point six, P L zero point nine. Paratype right dentary A R two two two zero one has m two and m three A with L two point seven and two point eight, A W one point five, P W one point six, A L one point six, P L one point one. Paratype right m two A R two two one nine three has m two with L two point seven, A W one point four, P W one point six, A L one point five, P L one point one. Paratype left m two A R two two two zero nine has m two with L two point six, A W one point three, P W one point five, A L one point five, P L one point one. Paratype left m three A R two two two zero seven has m three with L two point eight, A W one point six, P W one point six, A L one point five, P L one point three. Paratype left m three A R two two one eight one has m three with L two point four, A W one point four, P W one point four, A L one point four, P L one point zero. Dashes indicate missing data. Abbreviations are L for length, A W for anterior width, P W for posterior width, A L for anterior length, P L for posterior length, P C for preparacrista length, M D for metacone to st D, P M for postmetacrista length, T W for talon width, and M R L for molar row length.
Diagnosis
Species of Phantasmodon n. gen. differ from all other members of Keeunamorphia n. ord. in the following combination of upper and lower dental features: internal angle of M3 centrocrista larger (~80°, compared to ~60° in D. murgonensis, K. woodburnei, and A. tirarensis); stD higher crowned than stB on M3 (lower crowned than stB in D. murgonensis, K. woodburnei, and A. tirarensis); presence of short posterior cingulum (polymorphic on M1–3 in P. travouilloni n. gen. n. sp.; absent in D. murgonensis, K. woodburnei, and A. tirarensis); metacristids oblique on all molars (transverse in D. murgonensis and A. tirarensis); posthypocristids not transversely oriented on m1–3 (transverse in D. murgonensis and A. tirarensis); central cusp comprised of two smaller cusps in the unworn condition (rather than a single cusp in D. murgonensis, A. tirarensis, and K. woodburnei).
In addition to the features mentioned above, the dentitions of species of Phantasmodon n. gen. also differ from that of the early Eocene Djarthia murgonensis in the following features: M1 preparacrista transversely oriented (posterobuccally oriented in D. murgonensis); M3 preparacrista contacts the lingual base of stB (preparacrista contacts anterior base of stB on M3 in D. murgonensis); stD larger than stC on M1–3; stC more proximal to stD than to stB (stC equidistant from stB and stD in D. murgonensis); stC not connected to stB via crista on M2–3; stD situated subequal with metacone along the long axis of the tooth (stD situated posterior to metacone in D. murgonensis); metastylar corner of tooth is less elongated with shorter postmetacrista on M2 and M3; preparacrista longer on M2 (2/3 the length of postmetacrista in the new taxon; 1/2 the length in D. murgonensis); stC smaller and more closely twinned with stD on M2–3 (approaching stB in crown height and not closely twinned with stD in D. murgonensis); ectoflexus less developed on M2–3; preparacrista and postmetacrista less transversely oriented on M3; stE present on M1–3; M4 less reduced in length relative to M1–3 with a less transversely oriented buccal margin; metacone approaches paracone in crown height on M4; central cusp on M4 connected to the metastylar corner of the tooth via posterior crista (M4 central cusp lacks crista in D. murgonensis); stylar cusp C large and comprises a single cusp on M4 (rather than two small cusps in D. murgonensis); M4 metacone more proximal to the metastylar corner; postmetacrista absent on M4 (small but distinct in D. murgonensis); metaconule higher crowned than protocone (approximately subequal in D. murgonensis); m1 internal trigonid angle comprised of the paracristid and metacristid approximately 90° (~70° in D. murgonensis); m3 cristid obliqua less transverse in orientation (~45° off transverse axis, compared to ~35° in D. murgonensis); entoconid larger than hypoconulid on m4.
In addition to the features mentioned above, the dentitions of the new species of Phantasmodon n. gen. differ from that of the late Oligocene Keeuna woodburnei in the following features: M3 ectoloph narrower; M3 preparacrista approximately two-thirds the length of postmetacrista (~3/4 length of postmetacrista in K. woodburnei); M3 metastylar corner more elongated, with less transversely oriented postmetacrista; M3 ectoflexus situated anteriorly (rather than centrally in K. woodburnei); stC and stD are distinct cusps and higher crowned than stB (stylar cusps in the posterior half of the tooth is a row of minute cuspules of indistinct homology in K. woodburnei); stE lower crowned than stC and stD on M3 (stE marginally higher crowned than stC and stD in K. woodburnei); talonids less longitudinally reduced; anterior cingulids narrower; hypoconulids less reduced (minute in K. woodburnei).
In addition to the features mentioned above, the dentition of the new species of Phantasmodon n. gen. differs from that of the late Oligocene Ankotarinja tirarensis in the following features: ectolophs narrower; ectoflexus less developed; stC higher crowned than stB on M2 and subequal with stB on M3 (stB higher crowned than stC on M2–3 in A. tirarensis); metastylar corner less elongated on M2–3; metacone apex oriented dorsally on M2–3 (rather than posterodorsally in A. tirarensis); M2 talon broader with protocone apex more anteriorly situated; stD higher crowned than stB on M3; stD situated anterobuccally to metacone (rather than slightly posterobuccally in A. tirarensis); M3 central cusp larger with well-developed cristae (small and lacks cristae in A. tirarensis); centrocrista angle ~60° (rather than ~80° in A. tirarensis); internal talon angle ~110° (rather than ~95° in A. tirarensis); no anterior lean of the trigonids (present in A. tirarensis); m1 anterior cingulid narrow (broad in A. tirarensis); metacristids not transverse (transverse on m2–4 in A. tirarensis); m4 less reduced in length compared to m1–3; m4 entoconid large (reduced in A. tirarensis); m4 talonid broader.
Occurrence
The holotype maxilla (QM F61741; Fig. 1) and all isolated molar specimens (Figs. 2 and 4) were recovered from Dirk’s Towers Site. The paratype dentary (QM F61742; Fig. 3) was recovered from Neville’s Garden Site. Both localities are Lower Miocene deposits from the Riversleigh World Heritage Area. Neville’s Garden Site has been radiometrically dated to 18.24–17.85 Ma, within Riversleigh’s Faunal Zone B (Woodhead et al., Reference Woodhead, Hand, Archer, Graham, Sniderman, Arena, Black, Godthelp, Creaser and Price2016). Dirk’s Towers Site has not been radiometrically dated but is estimated to be Early Miocene in age within Riversleigh’s Faunal Zone B based on biocorrelation with radiometrically dated Early Miocene sites at Riversleigh (including Neville’s Garden) and the Etadunna Formation (Travouillon et al., Reference Travouillon, Archer, Hand and Godthelp2006, Reference Travouillon, Legendre, Archer and Hand2009, Reference Travouillon, Escarguel, Legendre, Archer and Hand2011; Arena et al., Reference Arena, Travouillon, Beck, Black, Gillespie, Myers, Archer and Hand2015).
Description
The M1 is tribosphenic in overall morphology. There is an anterior cingulum present that is continuous with the preprotocrista and stA. The cusp height in decreasing order is as follows: metacone, stD, stB subequal with paracone, metastylar corner, stC, stE, stA, protocone, metaconule. The preparacrista is short, strongly V-shaped in anterior view, and oriented perpendicular to the long axis of the tooth. This crista connects the paracone to stB, being approximately half the length of the distance between the metacone and stD. The stB is large and is connected to the stC via a short, high-crowned crista. The stC is more proximal to stD than to stB in specimens AR22926–7 (Fig. 2.2 and 2.4) but is equidistant between stB and stD in paratype specimens AR22170 (Fig. 2.1), AR22173 (Fig. 2.3), and AR22171 (Fig. 2.5). The stD is large and situated slightly posterior to the metacone along the long axis of the tooth. The stD possesses a distinct anterior crista emanating directly lingually from the crown into the base of the metacone, and a distinct posterior crista that is oriented posteriorly and contacts a tiny stE proximal to the metastylar corner of the tooth. The buccal margin of stD is the most buccal point of the tooth in occlusal view. The centrocrista is weakly V-shaped, proximal to the buccal margin of the tooth, and raised slightly above the ectoloph basin, with an internal angle of approximately 90–100°. The postparacrista is slightly shorter than the premetacrista. The postmetacrista is the longest tooth blade, being approximately three times longer than the preparacrista, oriented ~30° relative to the transverse axis, and is variable in length among the paratype specimens. The internal angle of the protocone formed by the preprotocrista and postprotocrista is ~120°. The protocone is anteriorly situated on the talon, being almost subequal with the paracone along the long axis of the tooth. The metaconule is large (approaching protocone in height), with a distinct postmetaconulecrista that continues beyond the base of the metacone, forming a narrow posterior cingulum in some of the paratype specimens (AR22170–2 and AR22927; Fig. 2.1 and 2.3–2.6). There are two small central cusps (one on the postparacrista and the other on the premetacrista) present on the ectoloph and situated sightly buccal to the centrocrista.
The morphology of M2 (based on paratypes AR22187 and QM F30872; Fig. 2.7 and 2.8) is similar to the M1, except in the following features: tooth more square in occlusal shape, with maximum width approaching subequal to maximum length; anterior cingulum narrower; buccal margins of stB and stD approximately subequal along transverse axis; metastylar corner less elongated; stA smaller and closer to stB; preparacrista longer and continuous in crown height, being approximately two-thirds length of postmetacrista; stC larger and situated midway between stB and stD; posterior blade emanating from apex of stD shorter; paracone less reduced relative to metacone; lingual angle of centrocrista smaller; postmetacrista oriented more transversely; metaconule larger.
The morphology of M3 (based on holotype QM F61741; Fig. 1) is similar to that of M2, except in the following features: stA higher crowned and almost subequal in height with stB; buccal boundary of stB in occlusal view extends farther than buccal margin of stD and of metastylar corner; preparacrista approximately three-quarters length of postmetacrista and slightly longer than postparacrista and premetacrista; small crista emanating posterolingually from stB that contacts central cusp, which in its entirety forms a raised ridge that runs lingual to stC and terminates just prior to anterior crista of stD in holotype QM F61741 but absent in AR22928–9 and AR22200 (likely a consequence of increased wear of the principal cusps); central cusp situated between stC and stD along long axis of tooth; stC larger and higher crowned than stB, and situated more proximal to stD than to stB; stD tallest cusp on crown and situated anterior to metacone apex along long axis of tooth; slight ectoflexus situated on buccal margin of tooth between stB and stC; no posterior cingulum present in holotype specimen QM F61742 (Fig. 1) and isolated paratype specimen AR22928 (Fig. 2.10), with postmetaconulecrista terminating ~0.5 mm beyond lingual base of metacone, but short and narrow posterior cingulum in paratype specimens AR22200 (Fig. 2.9), AR22188 (Fig. 2.11) and AR22929 (Fig. 2.12); metaconule higher crowned than protocone; distinct wear facet on postmetaconulecrista just posterior to metaconule in holotype (QM F61741; Fig. 1) and paratype (AR22200; Fig. 2.10).
The M4 is similar to M3 (based on holotype QM F61741; Fig. 1), except in the following features: stA closer to stB; stB tallest cusp on crown; preparacrista and central cusp anterior crista do not contact stB apex directly, but terminate at its lingual base; large stC situated midway along stylar shelf slightly closer to metastylar corner than to stB; stC possesses distinct anterior crista that emanates lingually from apex of cusp, before turning abruptly anterobuccal to contact preparacrista at lingual base of stB; posterior crista emanating from apex of stB that buccally overlaps with anterior crista of stC and terminates approximately halfway between stylar cusps B and C; posterior crista of stC descends lingually from apex to contact central cusp before turning ~90° posteriorly to contact metastylar corner of tooth (blade encroaches lingually onto ectoloph; likely the result of apical wear); metacone and metastylar corner close to one another but unfused; postmetacrista absent; premetacrista 2/3 length of postparacrista and 1/3 length of preparacrista; metacone and metastylar corner taller than paracone and stC, but slightly shorter than stB; talon narrower, with smaller internal protocone angle (~90°); postprotocrista 1/3 length of preprotocrista and contacts large metaconule, which is higher crowned than protocone and transversely twisted such that postmetaconulecrista terminates directly within lingual base of metacone; deep wear facet in cristid valley between postprotocrista and metaconule, which constitutes lowest point in talon basin.
The m1 is tribosphenic in overall morphology. The trigonid is narrower than the talonid, but they are approximately subequal in length. This tooth in paratype dentary QM F61742 (Fig. 3) is heavily worn; therefore, unworn cusp height on m1 in decreasing order is based on paratypes AR22182 (Fig. 4.1) and AR22208 (Fig. 4.2) and is as follows: protoconid, metaconid, paraconid = entoconid = hypoconid, hypoconulid. There is a narrow anterior cingulid present on the anterobuccal face of the trigonid. The internal angle of the trigonid formed by the paracristid and metacristid is approximately 90°. The paraconid is unreduced relative to the likely australidelphian plesiomorphic condition (inferred from dasyuromorphians and peramelemorphians) and is situated at the anterior extremity of the tooth. The paracristid is approximately three times the length of the metacristid. There is a paracristid carnassial notch separating the postparacristid from the longer preprotocristid (based on paratypes AR22182 and AR22208). In the paratype dentary specimen (QM F61742), the size of the protoconid and metaconid cannot be determined because the apices are considerably worn and form a continuous trigonid shelf that slopes posterolingually from the protoconid to the metaconid, approaching the talonid basin in height. However, in the unworn isolated paratype specimens (AR22182 and AR22208), the metaconid is unreduced with respect to the likely plesiomorphic australidelphian condition (retained by plesiomorphic dasyuromorphians and peramelemorphians) and is situated posterolingual to the protoconid. The metacristid is short with a distinct carnassial notch and is oriented ~10–15° off the transverse axis. The entoconid and hypoconid are markedly more robust and higher crowned than the hypoconulid. There is a well-developed preentocristid carnassial notch where the preentocristid meets the posterior base of the metaconid. The hypoconulid is reduced relative to the plesiomorphic dasyuromorphian condition, but similar to the condition in plesiomorphic peramelemorphians and projects posteriorly into a shallow hypoconulid notch on the anterior face of the m2 trigonid (hypoconulid notch absent in peramelemorphians). The hypoconulid is situated posterobuccal to the entoconid and is not closely twinned to that cusp. The hypoconid is the most buccally situated cusp on the crown. The posthypocristid is marginally longer than the cristid obliqua and is oriented approximately 30° off the transverse axis. The cristid obliqua is oriented approximately 20° off the long axis and contacts the trigonid base just buccal to the position of the metacristid carnassial notch. The internal angle of the hypoconid, which is comprised of the cristid obliqua and the posthypocristid, is approximately 90°. There is a broad posterior cingulid present on the posterior face of the crown that at its lowest contacts the buccal extremity of the hypoconid crown base, and at its highest terminates just ventral to the hypoconulid. There is a small but distinct buccal cingulid present in the hypoflexid region between the buccal extremities of the protoconid and hypoconid, which does not contact the anterior or posterior cingulids.
The m2 (based on paratypes QM F61742, AR22201, AR22193, and AR22209; Figs. 3 and 4.3–4.5) is the same as m1, except in the following features: trigonid broader overall, but still narrower than talonid; internal angle of trigonid smaller (~75°); paraconid more lingually situated, being aligned with lingual margin of metaconid; metaconid larger and less proximal to protoconid; metacristid larger, approximately 2/3 length of paracristid; cristid obliqua longer and oriented farther off long axis of tooth (~30°); posthypocristid longer and oriented more transversely (~15° off transverse axis).
The m3 (based on paratypes QM F61742, AR22207, and AR22181; Figs. 3, 4.6, 4.7) is the same as m2, except in the following features: trigonid subequal or slightly narrower than talonid; internal angle of trigonid smaller (~65°); metacristid longer, approximately 3/4 length of paracristid; metacristid carnassial notch situated less proximal to metaconid, being approximately midway between that cusp and protoconid; entoconid less robust at base, with less-developed preentocristid carnassial notch; cristid obliqua oriented farther off long axis of tooth (~40°).
The m4 (based on paratype QM F61742; Fig. 3) is the same as m3, except in the following features: talonid narrower than trigonid; entoconid smaller, with less developed preentocristid carnassial notch; entoconid larger than hypoconid; cristid obliqua shorter and oriented almost parallel to long axis of tooth; posthypocristid shorter and approximately subequal in length to cristid obliqua; posterior cingulid absent; buccal cingulid shorter and retains a distinct vertical wear facet.
The paratype dentary (QM F61742) is missing the premolars, canine, incisors, articular condyle, angular process, posterior half of ascending ramus, and masseteric fossa. In lateral view, the dentary is dorsoventrally deepest below m3 and narrowest between the m4 and the ascending ramus. There are two mental foramina on the buccal face of the dentary that are ~2 mm apart and situated below the posterior root of m1. A molar length (~2.5 mm) separates the posterior extremity of m4 from the lowest point of the ascending ramus.
The following meristic gradients are based on all type specimens. Upper molar length decreases from M1 to M2, is subequal from M2 to M3 and decreases from M3 to M4. Upper molar width increases from M1 to M4. Paracone size increases from M1 to M4. Metacone size decreases from M1 to M2, is subequal from M2 to M3 and decreases from M3 to M4. Size of stB increases from M1 to M4. Size of stC increases from M1 to M3 and decreases from M3 to M4. Size of stD decreases from M1 to M3. Size of metastylar corner decreases from M1 to M2, is subequal from M2 to M3, and increases from M3 to M4. Central cusp size increases from M1 to M3 and decreases from M3 to M4. Protocone size is subequal from M1 to M3 and decreases from M3 to M4. Metaconule size decreases from M1 to M4. Protoconule size decreases from M1 to M4. Preparacrista length increases from M1 to M4. Preprotocrista length decreases from M1 to M2, is subequal from M2 to M3, and increases from M3 to M4. Postparacrista length increases from M1 to M3 and decreases from M3 to M4. Premetacrista length increases from M1 to M3 and decreases from M3 to M4. Postmetacrista length decreases from M1 to M2, is subequal from M2 to M3, and decreases from M3 to M4. Lower molar length increases from m1 to m2, is subequal from m2 to m3, and decreases from m3 to m4. Paraconid size increases from m1 to m3 and is subequal from m3 to m4. Protoconid size increases from m1 to m2, is subequal from m2 to m3, and decreases from m3 to m4. Entoconid, hypoconid, and hypoconulid size increase from m1 to m2, are subequal from m2 to m3, and decrease from m3 to m4. Paracristid length decreases from m1 to m4. Metacristid and cristid obliqua length increase from m1 to m3 and decrease from m3 to m4. Posthypocristid length increases from m1 to m2, is subequal from m2 to m3, and decreases from m3 to m4.
Etymology
The species name travouilloni honors Dr Kenny Travouillon who has made an enormous contribution to dasyuromorphian and peramelemorphian paleontology at Riversleigh and to marsupial paleontology overall.
Remarks
Phantasmodon travouilloni n. gen. n. sp. is distinguished from P. minuferox n. gen. n. sp. and all other keeunamorphians by a coherent suite of upper- and lower-molar characters that together define the morphology of the type species. In contrast to P. minuferox, P. travouilloni exhibits a narrower and more distinct anterior cingulid on m1–2 that remains separate from the buccal cingulid, longitudinally longer talonids on all lower molars, and longer cristids obliquae that are more strongly oblique relative to the long axis of the tooth. The internal trigonid angle on m1 is notably smaller (~90°), producing a more acute and shearing trigonid configuration than the broader (~110°) condition seen in P. minuferox n. gen. n. sp. Additionally, the posthypocristids of P. travouilloni n. gen. n. sp. are less transversely oriented, consistent with the oblique and non-transverse shearing arrangement that characterizes the genus overall.
The suite of features present in P. travouilloni n. gen. n. sp., including the distinctive stC–stD complex, the consistently higher-crowned stD on M3, the frequent development of short posterior cingula on M1–3, and the doubled central cusp in unworn upper molars, differentiates this taxon from members of Djarthia, Ankotarinja , and Keeuna. Slight differences within the type specimens largely reflect intraspecies variation and differences in wear rather than taxonomically significant disparity, and unworn isolated teeth confirm the consistent development of key diagnostic characters. Collectively, these attributes support recognition of P. travouilloni n. gen. n. sp. as a morphologically discrete species that represents the larger, more plesiomorphic member of the genus.
Phantasmodon minuferox new species
Holotype
QM F61739 is an isolated left m1 (Fig. 4.9).
Paratype
AR22195 is an isolated left m2 (Fig. 4.10). Dental measurements of all specimens of Phantasmodon minuferox are shown in Table 2.
Dental measurements of Phantasmodon minuferox n. gen. n. sp. holotype left m1 (QM F61739) and paratype left m2 (AR22195), and of Keeunidae n. fam. gen. and sp. indet. right m1 (QM F61740). Abbreviations: L, length; AW, anterior width; PW, posterior width; AL, anterior length; PL, posterior length

Table 2. Long description
The table has seven columns labeled Specimen, Tooth, L, AW, PW, AL, and PL. The first row lists Phantasmodon minuferox holotype left m1 QM F61739 with values m1, 2.0, 0.9, 1.1, 1.2, 0.8. The second row lists Phantasmodon minuferox paratype left m2 AR22195 with values m2, 1.8, 0.9, 1.1, 1.0, 0.8. The third row lists Keeunidae gen. and sp. indet. right m1 QM F61740 with values m1, 2.6, 1.1, 1.3, 1.5, 1.1. L is length, AW is anterior width, PW is posterior width, AL is anterior length, PL is posterior length. All values are in the same units.
Diagnosis
Apart from its smaller size (~45 g), Phantasmodon minuferox n. gen. n. sp. differs from P. travouilloni n. gen. n. sp. (~103 g) in the following combination of lower dental features: anterior cingulid continuous with buccal cingulid on m1 and m2; talonids longitudinally shorter; cristids obliquae shorter; greater internal angle of m1 trigonid (~110°, compared to 90° in P. travouilloni n. gen. n. sp.); posthypocristids more transversely oriented.
Occurrence
The holotype and paratype specimens were recovered from Riversleigh’s Dirk’s Towers Site, which is estimated to be Early Miocene in age based on biocorrelation with radiometrically dated Early Miocene sites at Riversleigh (including Neville’s Garden) and the Etadunna Formation (Travouillon et al., Reference Travouillon, Archer, Hand and Godthelp2006, Reference Travouillon, Legendre, Archer and Hand2009, Reference Travouillon, Escarguel, Legendre, Archer and Hand2011; Arena et al., Reference Arena, Travouillon, Beck, Black, Gillespie, Myers, Archer and Hand2015).
Description
The m1 of Phantasmodon minuferox n. gen. n. sp. (QM F61739; Fig. 4.9) is tribosphenic in overall morphology, with a trigonid that is longer, wider, and higher crowned than the talonid. The cusp height in descending height order is as follows: protoconid; metaconid; paraconid = entoconid = hypoconid; hypoconulid. There is a well-developed and narrow anterior cingulid on the anterobuccal face of the crown that extends from just below the anterior tip of the paraconid to the buccal extremity of the protoconid. The paraconid is unreduced relative to the plesiomorphic tribosphenic condition within Australidelphia. The paracristid is the longest blade on the tooth and possesses a well-developed carnassial notch where the postparacristid meets the preprotocristid, with the latter approximately three times longer than the former. The protoconid is the largest cusp and is situated directly over the buccal half of the tooth and slightly anterior the longitudinal midpoint. The metaconid is unreduced relative to the plesiomorphic tribosphenic condition and is situated posterolingual to the protoconid. The metacristid, which is approximately 1/3 the length of the paracristid, is oriented approximately 30° to the transverse axis of the tooth. The postprotocristid and premetacristid are subequal in length and meet at a well-developed metacristid carnassial notch. The paracristid and metacristid form an internal angle of approximately 95°. The hypoconid is large and is the most buccally situated cusp on the crown. The cristid obliqua and the slightly longer posthypocristid meet at the hypoconid apex and form an internal angle of approximately 80°. The cristid obliqua is oriented approximately 25° off the longitudinal axis of the tooth and terminates into the posterior base of the trigonid at a point buccal to the metacristid carnassial notch. The entoconid is large, conical in occlusal view, and situated slightly anterior to the hypoconid relative to the longitudinal axis of the tooth. There is a well-developed preentocristid that emanates directly anteriorly from the entoconid apex to meet posterior base of the metaconid at a well-developed preentocristid carnassial notch. The hypoconulid is much smaller than the entoconid and hypoconid but is only slightly reduced relative to the plesiomorphic tribosphenic condition. The hypoconulid is situated posterobuccal to the entoconid and is not closely twinned with that cusp. The posthypocristid, which is oriented approximately 15° off the transverse axis of the tooth, contacts the hypoconulid at its anterobuccal base where it forms a slight V-shaped cristid notch. There is a well-developed and narrow posterior cingulid on the posterior face of the talonid, which contacts the posterior base of the hypoconulid at its highest and terminates just prior to the buccal extremity of the hypoconid crown base at its lowest. There is a well-developed buccal cingulid at the base of the crown in the hypoflexid region between the buccal extremities of the protoconid and hypoconid, which is continuous with the anterior cingulid, but not the posterior cingulid. The crown base of the talonid is markedly lower than that of the trigonid.
The m2 of this taxon (AR22195; Fig. 4.10) is similar to the m1, except for the following features: trigonid less enlarged in length compared to talonid; trigonid angle formed by paracristid and metacristid smaller (~60°); paracristid approximately 50% longer than metacristid; paraconid situated more lingually relative to metaconid; metaconid situated less posteriorly relative to protoconid resulting in a more transverse metacristid (~10° off transverse axis); cristid obliqua more transversely oriented (~20° off longitudinal axis) and shorter relative to posthypocristid.
Etymology
The species name minuferox is derived from minuere (Latin) meaning to make smaller, and ferox (Latin) meaning fierce: small but fierce.
Remarks
Phantasmodon minuferox n. gen. n. sp. is known only from isolated m1 and m2, but these teeth preserve a combination of features that clearly distinguishes the species from P. travouilloni n. gen. n. sp. Compared with the type species, P. minuferox exhibits an anterior cingulid that is continuous with the buccal cingulid on both lower molars, longitudinally shorter talonids, shorter and less oblique cristids obliquae, a markedly broader internal trigonid angle on m1 (~110°), and more transversely oriented posthypocristids. Although upper molars are unknown, the consistent expression of these differences in both preserved teeth indicates that P. minuferox n. gen. n. sp. represents a discrete and morphologically coherent species within Phantasmodon n. gen.
Keeunidae n. fam. genus incertae sedis
Included species
Keeunidae n. fam. gen. and sp. indet.
Remarks
Keeunidae n. fam. gen. and sp. indet. is established to accommodate a distinctive keeunamorphian represented by a single but highly diagnostic m1 that cannot be accommodated within any previously described genus. The unusual combination of features expressed in Keeunidae n. fam. gen. and sp. indet. including an exceptionally elongate and narrow m1, a trigonid only slightly longer than the talonid, a relatively tall paraconid, a cristid obliqua subequal in length to the posthypocristid, a long preentocristid produced by the entoconid’s close association with the hypoconulid, and non-continuous anterior and buccal cingulids differs markedly from the condition in Djarthia, Keeuna, Ankotarinja , and Phantasmodon n. gen. These combined attributes represent a structural configuration outside the documented range of variation for other keeunamorphian genera, which possess broader m1 crowns, differing relative cusp heights, more transverse posthypocristids, and distinct cingulid patterns. Despite being known only from m1, the preserved morphology of Keeunidae n. fam. gen. and sp. indet. is sufficiently divergent, particularly in overall proportions, cusp arrangement, and shearing blade geometry, to warrant generic distinction pending the discovery of additional material.
Keeunidae n. fam. genus and species indet.
Material
QM F61740 is an isolated right m1 (Fig. 4.8). Dental measurements of the m1 of Keeunidae n. fam. gen. and sp. indet. are shown in Table 2.
Diagnosis
The m1 of Keeunidae n. fam. gen. and sp. indet. differs from that tooth in other keeunamorphians in the following combination of m1 features: tooth occlusally narrower relative to total length; trigonid narrower relative to the talonid; talonid subequal to trigonid in length (rather than longer); paraconid taller than entoconid and hypoconid in unworn condition (rather than subequal); cristid obliqua subequal in length to posthypocristid (rather than shorter); entoconid more proximal to hypoconulid than to metaconid, resulting in a longer preentocristid; posthypocristid oriented less transversely (~30° off transverse axis); buccal cingulid on m1 not continuous with the anterior cingulid.
Occurrence
The specimen was recovered from Riversleigh’s Dirk’s Towers Site, estimated to be Early Miocene in age based on biocorrelation with radiometrically dated Early Miocene sites at Riversleigh (including Neville’s Garden) and the Etadunna Formation (Travouillon et al., Reference Travouillon, Archer, Hand and Godthelp2006, Reference Travouillon, Legendre, Archer and Hand2009, Reference Travouillon, Escarguel, Legendre, Archer and Hand2011; Arena et al., Reference Arena, Travouillon, Beck, Black, Gillespie, Myers, Archer and Hand2015).
Description
The m1 of Keeunidae n. fam. gen. and sp. indet. is tribosphenic in overall morphology. The trigonid is narrower and higher crowned than the talonid, and approximately subequal in length. The crown height of cusps in descending order is as follows; protoconid; metaconid; paraconid; entoconid = hypoconid; hypoconulid. There is a well-developed and narrow anterior cingulid on the anterobuccal face of the trigonid spanning just below the paraconid apex to the buccal extremity of the protoconid at the base of the crown. The paraconid is unreduced relative to the plesiomorphic tribosphenic condition. The paracristid is the longest tooth blade and possesses a well-developed paracristid carnassial notch where the postparacristid meets the 2.5× longer preprotocristid. The protoconid is the largest cusp and is situated anterobuccal of the tooth’s longitudinal and transverse midpoint. The metaconid is unreduced relative to the plesiomorphic tribosphenic condition and is situated posterolingual of the protoconid. The metacristid is approximately 1/3 the length of the paracristid and is oriented approximately 20° off the transverse axis. The metacristid retains a well-developed carnassial notch where the postprotocristid meets the similarly sized premetacristid. The internal angle of the trigonid formed by the paracristid and metacristid is approximately 80°. The hypoconid is large and situated subequal with the entoconid on the tooth’s longitudinal axis. The cristid obliqua is approximately subequal in length to the posthypocristid, with the two blades meeting at the apex of the hypoconid and forming an internal angle of approximately 85°. The cristid obliqua is oriented approximately 20° off the longitudinal axis of the tooth and contacts the posterior base of the trigonid at a point buccal to the metacristid carnassial notch. The entoconid is large, conical in occlusal view, and situated closer to the hypoconulid than to the base of the metaconid, resulting in a particularly long preentocristid. The hypoconulid, which is reduced relative to the plesiomorphic tribosphenic condition, is situated posterobuccal to the entoconid and not closely twinned with that cusp. There is a well-developed and narrow posterior cingulid situated on the posterior face of the crown, which spans the posterior base of the hypoconulid at its highest to the posterior base of the hypoconid at its lowest. There is a slight buccal cingulid present in the hypoflexid region between the buccal extremities of the protoconid and hypoconid, which is not continuous with the anterior cingulid or the posterior cingulid.
Remarks
Keeunidae n. fam. gen. and sp. indet. is represented only by a single, well-preserved m1, but this tooth exhibits a distinct combination of proportions and crest orientations that clearly differentiates the species from other keeunamorphians known from lower dentitions. The narrow and elongate m1, the relatively taller paraconid, the subequal lengths of the cristid obliqua and posthypocristid, and the unusually long preentocristid produced by the entoconid’s proximity to the hypoconulid collectively form a unique morphological profile not observed in Djarthia, Keeuna, Ankotarinja , or Phantasmodon n. gen. Although upper molars remain unknown, the consistency and diagnostic clarity of the preserved cusp patterning and blade geometry support recognition of Keeunidae n. fam. gen. and sp. indet. as a discrete species.
Results
Body mass
The body masses of the new taxa based on their type specimens are shown in Table 3. Phantasmodon travouilloni n. gen. n. sp. is estimated to be 58.8–163.3 g (median = 102.1 g; mean = 100.6 g). Phantasmodon minuferox n. gen. n. sp. is estimated to be 27.4–71.7 g (median = 45.1 g; mean = 47.3 g). Keeunidae n. fam. gen. and sp. indet. is estimated to be 125.7–199.7 g (mean = 162.7 g).
Body mass estimates of Phantasmodon travouilloni n. gen. n. sp., P. minuferox n. gen. n. sp., and Keeunidae n. fam. gen. and sp. indet. based on new material described by this study. All estimates are based on the best two dental regressions following Myers (Reference Myers2001). Abbreviations: 1LML, first lower molar length; 1LMW, first lower molar width; 1UML, first upper molar length; 1UMW, first upper molar width; 2LML, second lower molar length; 2LMW, second lower molar width; 2UML, second upper molar length; 2UMW, second upper molar width; 3LML, third lower molar length; 3LMW, third lower molar width; 3UML, third upper molar length; 3UMW, third upper molar width; LMRL, lower molar row length

Table 3. Long description
The table has three columns: Specimen, Measurement, and Body Mass in grams. The header row is followed by rows for each specimen. For Phantasmodon travouilloni holotype right maxilla (QM F61741), measurements 3UML and 3UMW correspond to body masses 110.3 and 68.6. The paratype left dentary (QM F61742) with LMRL and 3LML gives 131.4 and 79.9. Paratype right M1 (AR22170) with 1UML and 1UMW gives 92.5 and 98.9. Paratype left M1 (AR22926) with 1UML and 1UMW gives 101.1 and 87.4. Paratype left M1 (AR22927) with 1UML and 1UMW gives 101.1 and 107.2. Paratype left M1 (AR22171) with 1UML and 1UMW gives 123.4 and 85.6. Paratype left M1 (AR22172) with 1UML and 1UMW gives 84.4 and 87.4. Paratype left M1 (AR22173) with 1UML and 1UMW gives 153.1 and 135.2. Paratype right M2 (AR22187) with 2UML and 2UMW gives 86.6 and 60.6. Paratype left M2 (QM F30872) with 2UML and 2UMW gives 106.1 and 81.7. Paratype right M3 (AR22928) with 3UML and 3UMW gives 110.3 and 58.8. Paratype right M3 (AR22188) with 3UML and 3UMW gives 94.5 and 85.5. Paratype right M3 (AR22200) with 3UML and 3UMW gives 134.1 and 79.6. Paratype right M3 (AR22931) with 3UML and 3UMW gives 124.8 and 58.8. Paratype left M3 (AR22929) with 3UML and 3UMW gives 124.8 and 79.6. Paratype left m1 (AR22182) with 1LML and 1LMW gives 163.3 and 115.6. Paratype left m1 (AR22208) with 1LML and 1LMW gives 140.5 and 103.1. Paratype right dentary (AR22201) with 2LML and 2LMW gives 123.7 and 134.5. Paratype right m2 (AR22193) with 2LML and 2LMW gives 127.1 and 109.0. Paratype left m2 (AR22209) with 2LML and 2LMW gives 111.0 and 96.4. Paratype left m3 (AR22207) with 3LML and 3LMW gives 126.6 and 93.1. Paratype left m3 (AR22181) with 3LML and 3LMW gives 72.8 and 59.2. For Phantasmodon minuferox holotype left m1 (QM F61739), 1LML and 1LMW give 71.7 and 62.7. Paratype left m2 (AR22195) with 2LML and 2LMW gives 27.4 and 27.3. For Keeunidae gen. and sp. indet. right m1 (QM F61740), 1LML and 1LMW give 199.7 and 125.7. Abbreviations for measurements are defined in the caption.
Phylogenetic results
Maximum parsimony
The strict consensus tree (length = 1154; CI excluding autapomorphies = 0.246; HI excluding autapomorphies = 0754; RI = 0.518; RC = 0.131) summarizing the 47520 most parsimonious trees produced from the initial maximum parsimony analysis (Fig. 5) weakly supports a sister taxon relationship between Phantasmodon travouilloni n. gen. n. sp. and Keeuna woodburnei (bootstrap < 50%). The unambiguous synapomorphies (determined using ACCTRAN and DELTRAN optimizations) uniting P. travouilloni n. gen. n. sp. and K. woodburnei to the exclusion of other keeunamorphians are as follows: stylar cusp C large on M4 (character 35, character state 1; CI = 1.0); metacristids oblique on all lower molars (character 58, character state 2, CI = 0.286). The apomorphies differentiating P. travouilloni n. gen. n. sp. from K. woodburnei are as follows: stylar cusp D large on M3 (character 34, character state 0; CI = 0.25). This clade is recovered within a polytomous clade alongside Djarthia murgonensis and Ankotarinja tirarensis (bootstrap = 53%).
The unambiguous synapomorphies uniting the P. travouilloni + K. woodburnei, D. murgonensis, and A. tirarensis clade are as follows: termination of preparacristae at the lingual base of stB on M2–4 (character 20, character state 2; CI = 0.4); central cusp present on upper molars (character 36, character state 1; CI = 1.0); metaconule large on M4 (character 39, character state 1; CI = 1.0); p3 longer than p2 (character 46, character state 0; CI = 0.1); posterior cingulid present on m1–3 (character 59, character state 0, CI = 0.33); three distinct cusps on m4 (character 65, character state 0; CI = 0.083); entoconid large on m4 (character 66, character state 0; CI = 0.2). This clade is recovered as a sister group to the clade containing Dromiciops gliroides + Microbiotherium tehuelchum Ameghino, 1897, all members of Peramelemorphia, and all members of Dasyuromorphia.
Monophyly of Marsupialia is well supported by this analysis (bootstrap = 80%), as is the clade containing all members of Marsupialia and the sparassodontan Mayulestes ferox (bootstrap = 79%), to the exclusion of Andinodelphys cochabambensis and Pucadelphys andinus. This analysis provides moderately strong support for the monophyly of Australidelphia, which includes all members of Keeunamorphia n. ord., Microbiotheria, Peramelemorphia, and Dasyuromorphia (bootstrap = 55%), and excludes members of Didelphimorphia, Sparassodonta, and Pucadelphyidae.
The unambiguous synapomorphies uniting members of Australidelphia including keeunamorphians (among keeunamorphians, only Djarthia murgonensis is scored for cranial and postcranial characters) are as follows: centrocrista strongly V–shaped (character 16, character state 2; CI = 0.5); preparacrista oriented perpendicular to long axis on M1 (character 18, character state 1; CI =0.5); prootic canal absent (character 88, character state 1; CI = 0.2); frontal-squamosal contact on the lateral wall of the braincase (character 89, character state 1; CI = 0.2); C5 anteroposteriorly shorter than T1 (character 133, character state 1; CI = 0.14); T11 position of the diaphragmatic vertebrae (character 137, character state 2; CI = 0.25); dorsoventrally oriented L3–L4 spinous process (character 145, character state 1; CI = 0.17); weakly developed keel in the manubrium (character 159, character state 0; CI = 0.11); extension of the deltopectoral crest of the humerus restricted to the proximal half (character 178, character state 0; CI = 0.2); fossa for the exterior ligament on the ulna extends beyond trochlear notch (character 186, character state 2; CI = 0.14); olecranon process subequal in length to semilunar notch on ulna (character 190, character state 1; CI = 0.25); calcaneocuboid facet of the calcaneus subdivided into auxillary (CaCua), lateral (CaCul), and medial facets (character 215, character state 2; CI = 1.0); merged calcaneal sustentacular facet and posterior calcaneoastragalar facet (character 216, character state 2; CI = 0.4).
Within Australidelphia, Microbiotheria is recovered as sister to the remaining taxa, with Peramelemorphia recovered as sister to Dasyuromorphia (bootstrap < 50%). Within Dasyuromorphia, monophyletic Thylacinidae (bootstrap < 50%) is recovered as sister to all other dasyuromorphians (bootstrap < 50%). Within Thylacinidae, there is moderate support for the clade containing species of Ngamalacinus (bootstrap = 64%), the clade containing species of Thylacinus and Wabulacinus (bootstrap = 82%), the clade containing species of Thylacinus (bootstrap = 78%), and the clade containing species of Wabulacinus (bootstrap = 56%). The putative thylacinid Mutpuracinus archibaldi Murray and Megirian, Reference Murray and Megirian2000, is recovered outside Thylacinidae as a sister taxon to a polytomous clade containing Malleodectidae, Dasyuridae, and Myrmecobiidae (bootstrap < 50%). Monophyly of Malleodectidae containing Protamalleus stevewroei Churchill, Archer, and Hand, Reference Churchill, Archer and Hand2025b, and Barinya wangala Wroe, Reference Wroe1999, as the first and second taxa to diverge, respectively, is weakly supported (bootstrap < 50%). Within Malleodectidae, the clade including Chitinodectes wessechresti Churchill, Archer, and Hand, Reference Churchill, Archer and Hand2025b, Exosmachus robinbecki Churchill, Archer, and Hand, Reference Churchill, Archer and Hand2025b, and species of Malleodectes is moderately supported (bootstrap = 56%), as is the clade containing species of Malleodectes + Exosmachus robinbecki (bootstrap = 53%) and the clade containing species of Malleodectes (bootstrap = 73%). Within Dasyuridae, there is moderate support for the dasyurid subfamily Sminthopsinae (bootstrap = 66%) and for the dasyurid tribes Sminthopsini (bootstrap = 73%), Dasyurini (bootstrap = 51%), and Phascogalini (bootstrap = 75%).
The strict consensus tree (length = 1173; CI excluding autapomorphies = 0.242; HI excluding autapomorphies = 0.758; RI = 0.523; RC = 0.130) summarizing the 300,960 most parsimonious trees produced from the maximum parsimony analysis including the taxa Carolopaulacoutoia itaboraiensis, Itaboraidelphys camposi, Didelphopsis sp., and Peratherium constans (Fig. 6), weakly supports placement of the Phantasmodon travouilloni n. gen. n. sp. as a sister taxon to Keeuna woodburnei (bootstrap < 50%) and within a clade containing Ankotarinja tirarensis and Djarthia murgonensis (bootstrap < 50%). The unambiguous synapomorphies uniting Phantasmodon travouilloni n. gen. n. sp. and K. woodburnei are the same as in the previous parsimony analysis. The unambiguous synapomorphies uniting the clade containing P. travouilloni + K. woodburnei, A. tirarensis, and D. murgonensis are as follows: centrocrista strongly V-shaped (character 16, character state 2; CI = 0.44); termination of preparacristae at the lingual base of stB (character 20, character state 2; CI = 0.4); stD situated directly buccal to metacone on M1–3 (character 32, character 0; CI = 0.33).
This analysis recovers this clade within a weakly supported (bootstrap = < 50%) polytomous clade with the sternbergiids C. itaboraiensis, I. camposi, and Didelphopsis sp., and the herpetotheriid Peratherium constans. The unambiguous apomorphies uniting that clade are as follows: metacone large on M4 (character 15, character state 0; CI = 0.2); stD situated directly buccal to metacone (character 32, character state 1; CI = 0.33); central cusps present on upper molars (character 36, character state 1; CI = 1.0); metaconule large on M4 (character 39, character state 1; CI = 0.5); p3 shorter in height than p2 (character 45, character state 2; CI = 0.18); p3 shorter in length than p2 (character 46, character 0; CI = 0.1); posterior cingulid present on m1–3 (character 59, character state 0; CI = 0.33); three distinct cups on m4 talonid (character 65, character state 0; CI = 0.083); entoconid large on m4 (character 66, character state 0; CI = 0.2). That clade was recovered in a polytomy (bootstrap < 50%) with the microbiotheriids Dromiciops gliroides and Microbiotherium tehuelchum (bootstrap = 70%) and a clade containing all other members of Australidelphia (i.e., peramelemorphians and dasyuromorphians; bootstrap < 50%).
The clade containing keeunamorphians + sternbergiids + microbiotheriids + peramelemorphians + dasyuromorphians was recovered as a sister group to a clade containing members of Didelphimorphia (bootstrap = 71 %) within a moderately supported clade representing Marsupialia (bootstrap = 83%). The unambiguous synapomorphies uniting the clade containing keeunamorphians + P. constans + sternbergiids, microbiotherians, and peramelemorphians + dasyuromorphians are the same as those uniting keeunamorphians and other australidelphians in the previous analysis (Fig. 5), except for including presence of metacone that is indistinct from metastylar corner on M4 (character 15, character state 1; CI = 0.2) and excluding centrocrista V-shaped on upper molars (character 16) and absence of prootic canal (character 88). All other tree topologies are generally consistent with the previous parsimony analysis.
Dated Bayesian total evidence analyses
The 50% majority-rule consensus tree (Fig. 7) produced from the tip-and-node dated Bayesian analysis recovers Phantasmodon travouilloni n. gen. n. sp. within a well-supported clade with Keeuna woodburnei (BPP = 0.79) to which Ankotarinja tirarensis is a sister taxon within a larger, well-supported clade (BPP = 0.97). This larger clade is recovered within a weakly supported polytomy (BPP = 0.52) with a well-supported clade containing microbiotherians (BPP = 1.0) and a weakly supported clade containing peramelemorphians and dasyuromorphians (BPP = 0.55). Djarthia murgonensis is recovered as the sister taxon to the polytomy containing all other members of Australidelphia, all within a well-supported clade (BPP = 0.97) that is the sister group to a clade containing didelphids (BPP = 1.0). The sparassodontan Mayulestes ferox is recovered as the sister taxon to the clade containing australidelphians and didelphids (BPP = 0.5). The pucadelphyids Andinodelphys cochabambensis and Pucadelphys andinus are recovered in a strongly supported clade (BPP = 0.98) that is sister to a clade containing all other taxa in the analysis (BPP = 1.0).
Within Dasyuromorphia, there is moderate support for Thylacinidae excluding Muribacinus gadiyuli Wroe, Reference Wroe1996, Mutpuracinus archibaldi, and Tyarrpecinus rothi Murray and Megirian, Reference Murray and Megirian2000 (BPP = 0.81), moderate support for Malleodectidae excluding Protamalleus stevewroei (BPP = 0.70), and strong support for Dasyuridae (BPP = 0.93). Within Thylacinidae, the clades formed by Ngamalacinus timmulvaneyi Muirhead, Reference Muirhead1997, and Ng. nigelmarveni Churchill, Archer, and Hand, Reference Churchill, Archer and Hand2024 (BPP = 0.95), Badjcinus turnbulli Muirhead and Wroe, Reference Muirhead and Wroe1998, and Bad. timfaulkneri Churchill, Archer, and Hand, Reference Churchill, Archer and Hand2024 (BPP = 0.82), Wabulacinus ridei Muirhead, Reference Muirhead1997, and W. macknessi (Muirhead, Reference Muirhead1992) (sensu Churchill et al., Reference Churchill, Archer and Hand2024) (BPP = 0.86), and Thylacinus species (BPP = 0.99) are all well supported. Nimbacinus dicksoni Muirhead and Archer, Reference Muirhead and Archer1990, and Ni. peterbridgei Churchill, Archer, and Hand, Reference Churchill, Archer and Hand2024, form a weakly supported clade (BPP = 0.50). This analysis strongly supports a sister group relationship between species of Wabulacinus and species of Thylacinus (BPP = 0.99). Sister to Thylacinidae is a polytomous clade (BPP = 0.74) containing Malleodectidae, Dasyuridae, Pr. stevewroei, Myrmecobius fasciatus Waterhouse, Reference Waterhouse1836, Ty. rothi, Mur. gadiyuli, and Mut. archibaldi. Within Malleodectidae, there is moderate support for a clade containing Barinya wangala, Chitinodectes wessechresti + Exosmachus robinbecki + Malleodectes species (BPP = 0.70), strong support for a polytomy containing C. wessechresti, E. robinbecki, and Malleodectes species (BPP = 0.92), a clade containing E. robinbecki and Malleodectes species (BPP = 1.0), and a clade containing species of Malleodectes (BPP = 0.99). Within Dasyuridae (BPP = 0.93), there is strong clade support for the dasyurid subfamilies Sminthopsinae (BPP = 0.97) and Dasyurinae (BPP = 0.98), and for the dasyurid tribes Sminthopsini (BPP = 0.98), Phascogalini (BPP = 1.0), and Dasyurini (BPP = 1.0).
The tip-and-node calibrated date estimates for major divergences in this analysis (Fig. 7) suggest keeunamorphians excluding Djarthia murgonensis diverged from the common ancestor of dasyuromorphians, peramelemorphians, and microbiotherians sometime during the early Eocene (median estimate = 51.8 Ma; 95% HPD = 56.3–46.8 Ma). Djarthia murgonensis diverged from other members of Australidelphia sometime during the late Paleocene (median estimate = 56.8 Ma; 95% HPD = 60.1–54.7 Ma). Ankotarinja tirarensis diverged from the common ancestor of Keeuna woodburnei and Phantasmodon travouilloni n. gen. n. sp. sometime during the early Oligocene (median estimate = 31.4 Ma; 95% HPD = 39.7–25.8 Ma), which diverged from one another sometime during the late Oligocene or very early Miocene (median estimate = 26.9 Ma; 95% HPD = 31.2–24.0 Ma). The earliest common ancestor of dasyuromorphians + peramelemorphians diverged sometime during the early Eocene (median estimate = 48.7 Ma; 95% HPD = 54.8–43.2 Ma), with the last common ancestor of dasyuromorphians present sometime during the middle Eocene (median estimate = 38.5 Ma; 95% HPD = 43.4–34.3 Ma).
Thylacinids are estimated to have diverged into basal lineages sometime during the late Eocene or early Oligocene (median estimate = 35.2 Ma; 95% HPD = 39.7–30.9), followed by myrmecobiids during the early Oligocene (median estimate = 33.9 Ma; 95% HPD = 38.6–29.3 Ma), malleodectids (excluding Protamalleus stevewroei) during the late Oligocene (median estimate = 26.4 Ma; 95% HPD = 31.3–21.6 Ma), and dasyurids during the Early Miocene (median estimate = 22.5 Ma; 95% HPD = 26.2–18.6 Ma). The crown lineages of the dasyurid subfamily Dasyurinae diverged sometime during the Early Miocene (median estimate = 19.5 Ma; 95% HPD = 22.8–15.9 Ma) and the dasyurid subfamily Sminthopsinae sometime during the Middle Miocene (median estimate = 15.7 Ma; 95% HPD = 19.2–11.9 Ma).
The majority-rule tree (Fig. 8) produced from the additional tip-and-node dated Bayesian analysis with the sternbergiids Didelphopsis spp., Carolopaulacoutoia itaboraiensis, and Itaboraidelphys camposi and the herpetotheriid Peratherium constans recovers keeunamorphians excluding Djarthia murgonensis in a well-supported clade (BPP = 0.93) sister to dasyuromorphians. Within the keeunamorphian clade, Phantasmodon travouilloni n. gen. n. sp. forms a moderately supported clade with Keeuna woodburnei (BPP = 0.68) that is sister to Ankotarinja tirarensis (BPP = 0.92). Keeunamorphians (excluding D. murgonensis) and dasyuromorphians form a weakly supported clade (BPP = 0.58) that is sister to peramelemorphians (BPP = 1.0). Peramelemorphians and keeunamorphians (excluding D. murgonensis) + dasyuromorphians form a moderately supported clade (BPP = 0.72) that is recovered in a polytomy (BPP = 0.58) with microbiotherians (BPP = 1.0), didelphids (BPP = 1.0), Djarthia murgonensis, and the sternbergiids Didelphopsis spp., Carolopaulacoutoia itaboraiensis, and Itaboraidelphys camposi. This polytomy is recovered as the sister group to the herpetotheriid Peratherium constans (BPP = 1.0). The remaining phylogenetic tree produced in this analysis (Fig. 8) is generally consistent with that produced by the initial dated Bayesian analysis without sternbergiids and Peratherium constans (Fig. 7), except for in the following results: species of Nimbacinus do not form a clade but instead form part of a basal polytomy containing well-supported clades representing Ngamalacinus, Badjcinus, and Wabulacinus + Thylacinus.
The tip-and-node divergence estimates for the second Bayesian analysis including sternbergiids and Peratherium constans (Fig. 8) closely resemble the results of the first Bayesian analysis (Fig. 7), except for the following key differences: keeunamorphians excluding Djarthia murgonensis share a common ancestor with dasyuromorphians sometime during the middle Eocene (median estimate = 42.0 Ma; 95% HPD = 49.5–35.7 Ma); basal divergence of Dasyuridae approximately 1 million years later (median estimate = 21.5 Ma; 95% HPD = 25.6–15.7 Ma); sternbergiids share a common ancestor with australidelphian and didelphid marsupials sometime during the middle Paleocene (median estimate = 59.5 Ma; 95% HPD = 63.9–57.0 Ma); australidelphians + sternbergiids + didelphids + herpetotheriids share a common ancestor sometime during the early Paleocene (median estimate = 62.6 Ma; 68.6–57.8 Ma).
Discussion
Taxonomy and phylogenetics
There are only two known unambiguous morphological synapomorphies for australidelphians and both are features of the tarsal bones (Szalay, Reference Szalay and Archer1982, Reference Szalay1994; Beck, Reference Beck2012). However, the phylogenetic analyses of Beck et al. (Reference Beck, Voss and Jansa2022) found three features of the petrosal region that may also be synapomorphies of Australidelphia. Although these skeletal regions are not preserved in the new taxa, petrosal and tarsus remains attributed to Djarthia murgonensis support its classification as a stem australidelphian (Beck et al., Reference Beck, Godthelp, Weisbecker, Archer and Hand2008b). However, described D. murgonensis specimens exhibit polymorphism in the petrosal region, which has limited the certainty of the exact phylogenetic placement of D. murgonensis (Sanchez-Villagra and Wible, Reference Sanchez-Villagra and Wible2002; Beck, Reference Beck2008; Beck et al., Reference Beck, Godthelp, Weisbecker, Archer and Hand2008b, Reference Beck, Voss and Jansa2022; Kealy and Beck, Reference Kealy and Beck2017; Sanchez-Villagra and Forasiepi, Reference Sanchez-Villagra and Forasiepi2017).
Support for Keeunamorphia n. ord. including Djarthia murgonensis as an australidelphian clade is moderate in both maximum parsimony phylogenetic analyses herein. The strongest support for assignment of Keeunamorphia n. ord. to Australidelphia is the maximum parsimony analysis without sternbergiids and Peratherium constans (Fig. 5), where the keeunamorphian clade is recovered as the sister group to all other australidelphian marsupials within a clade excluding didelphids and pucadelphydans. This result generally concurs with the results of previous phylogenetic studies that place the clade containing Djarthia murgonensis, Ankotarinja tirarensis, and Keeuna woodburnei as sister to the rest of the australidelphian marsupial radiation (Kealy and Beck, Reference Kealy and Beck2017; Beck et al., Reference Beck, Voss and Jansa2022; Churchill et al., Reference Churchill, Archer, Hand, Myers, Gillespie and Beck2023, Reference Churchill, Archer and Hand2024, Reference Churchill, Archer and Hand2025a, Reference Churchill, Archer and Handb). These results weakly support absence of the prootic canal as an australidelphian synapomorphy (CI = 0.20) and add a plethora of potential new unambiguous synapomorphies for that clade.
In the maximum parsimony analysis that includes sternbergiids and P. constans (Fig. 6), keeunamorphians form a weakly supported clade with these taxa, which is sister to Microbiotheria plus all other australidelphians and excludes didelphids and pucadelphyids. These results could imply that keeunamorphians, sternbergiids, herpetotheriids, and microbiotheriids lie just outside Australidelphia, or that they represent stem australidelphians. The former is unlikely given that Australidelphia was explicitly defined to include microbiotheriids and Dromiciops exhibits a characteristic australidelphian-type tarsus, whereas the latter is difficult to reconcile with the absence of australidelphian-type tarsals from Itaborai Formation (Szalay, Reference Szalay1994; Szalay and Sargis, Reference Szalay and Sargis2001; Beck, Reference Beck2017b) and from localities that yielded herpetotheriids such as the type locality of Peratherium constans (Coillot et al., Reference Coillot, Smith, Gigase and Smith2013). Overall, both parsimony analyses lend moderate support to the inclusion of Keeunamorphia n. ord. within Australidelphia but raise questions about which marsupialiform lineages may also belong to that clade (i.e., Sternbergiidae and Herpetotheriidae).
In the Bayesian analyses excluding sternbergiids and Peratherium constans (Fig. 7), Australidelphia is strongly supported, with Djarthia murgonensis recovered as a sister taxon to all other australidelphians and outside the clade containing Phantasmodon travouilloni n. gen. n. sp., Keeuna woodburnei, and Ankotarinja tirarensis. When sternbergiids and Pe. constans are added, an australidelphian clade including keeunamorphians and microbiotherians but excluding didelphimorphians was not recovered. Instead, it weakly supports a basal polytomy containing D. murgonensis, sternbergiid species, microbiotherians, didelphimorphians, and a clade containing all other australidelphians (Fig. 8). Peratherium constans diverges earlier than this basal polytomy. This topology suggests either that D. murgonensis and microbiotherians are not australidelphians (which is contradicted by current morphological and molecular understanding of australidelphian relationships), or that all modern marsupial groups including australidelphians derive from a D. murgonensis or sternbergiid-like common ancestor sometime during the Paleocene.
As with the maximum parsimony analysis, support for monophyletic Australidelphia is considerably reduced when sternbergiids and P. constans are added to the analysis, suggesting there may be a close relationship between those taxa and stem australidelphians such as D. murgonensis. Therefore, we tentatively assign D. murgonensis to the australidelphian radiation but note that our analyses have introduced some uncertainty as to the validity of these superordinal marsupialiform clades in phylogenetic analyses, despite overwhelming molecular support for Australidelphia (Nilsson et al., Reference Nilsson, Churakov, Sommer, Tran, Zemann, Brosius and Schmitz2010; Meredith et al., Reference Meredith, Janecka, Gatesy, Ryder and Fisher2011; Mitchell et al., Reference Mitchell, Pratt, Watson, Gibb, Llamas, Kasper, Edson, Hopwood, Male and Armstrong2014; Duchene et al., Reference Duchene, Bragg, Duchene, Neaves, Potter, Johnson, Ho and Eldridge2018; Beck et al., Reference Beck, Voss and Jansa2022). We note here that Beck et al. (Reference Beck, Voss and Jansa2022) also failed to identify any unambiguous craniodental synapomorphies of Australidelphia.
Djarthia murgonensis, Keeuna woodburnei, Ankotarinja tirarensis, Phantasmodon travouilloni n. gen. n. sp., Ph. minuferox n. gen. n. sp., and Keeunidae n. fam. gen. and sp. indet. are assigned to the new marsupial order, Keeunamorphia n. ord., rather than to Dasyuromorphia, Peramelemorphia, Sternbergiidae, Herpetotheriidae, or some other known metatherian order. This assignment is based on 13 derived craniodental features obtained from our morphological comparisons and maximum parsimony analyses that individually exhibit some homoplasy within Marsupialiformes but in combination are not present in any other known metatherian group. These are: presence of a central cusp in the upper molars; preparacristae that contact stB rather than stA; enlarged metaconules and reduced protoconules on M1–3; metaconule large and appressed to the base of the metacone on M4; large stD on M3; large stC on M4; unreduced metacone on M4; metacone larger than paracone on M1–3; ectolophs narrow via lingual intrusion of centrocrista; posterior cingulids on m1–3; entoconids approaching paraconids in height on m1–3; entoconid large on m4; and hypoconulids reduced on all lower molars.
The maximum parsimony analyses (Figs. 5 and 6) provide further support for the monophyly of Keeunamorphia n. ord., although the level of support varies depending on taxon sampling. When sternbergiids and Peratherium constans were excluded, six unambiguous synapomorphies supported the monophyly of Keeunamorphia with moderate bootstrap support. However, when sternbergiids and P. constans were included, the analysis recovered three synapomorphies for Keeunamorphia with < 50% bootstrap support.
In contrast, the Bayesian dated total evidence analyses place Djarthia murgonensis outside the clade containing Phantasmodon travouilloni n. gen. n. sp., Keeuna woodburnei, and Ankotarinja tirarensis, but rather as a stem member of Australidelphia (Fig. 7) or Marsupialia + Sternbergiidae (Fig. 8). However, support for all higher-level relationships in these analyses is weak. The exclusion of D. murgonensis from Keeunamorphia n. ord. in the Bayesian analyses is most plausibly an artefact of the dated molecular clock framework, which forces accommodation of a 29- to 37-million-year temporal gap between D. murgonensis from the early Eocene and the other known keeunamorphians, including the new taxon described here from the late Oligocene to Early Miocene. This effect has also been observed in recent Bayesian analyses of Riversleigh dasyuromorphians, where D. murgonensis failed to group with Keeuna woodburnei and Ankotarinja tirarensis, despite these taxa forming a clade under parsimony (Churchill et al., Reference Churchill, Archer and Hand2024, Reference Churchill, Archer and Hand2025a, Reference Churchill, Archer and Handb). Given these patterns, and the consistent support from parsimony and qualitative morphological comparisons, we consider the Bayesian placement of D. murgonensis outside Keeunamorphia n. ord. to be less plausible. Additional taxa from the temporal interval between the early Eocene and late Oligocene will be critical for resolving this issue.
Phantasmodon travouilloni n. gen. n. sp. and Ph. minuferox n. gen. n. sp. were assigned to a new genus, and Keeunidae n. fam. gen. and sp. indet. was left in open nomenclature, because they possess a combination of derived and plesiomorphic features not present in other keeunamorphians (see Systematic Paleontology). The upper molars of Ph. travouilloni (Fig. 9.1–9.3) most closely resembles those of K. woodburnei (Fig. 9.4 and 9.5), sharing several derived features also present in basal dasyuromorphians such as Barinya wangala (Fig. 9.8), and both parsimony and Bayesian analyses support a sister-taxon relationship between Ph. travouilloni n. gen. n. sp. and K. woodburnei. Phantasmodon travouilloni n. gen. n. sp. (Fig. 10.1), Ph. minuferox n. gen. n. sp. (Fig. 10.2), and Keeunidae n. fam. gen. and sp. indet. (Fig. 10.3) also share an oblique orientation of the metacristids with K. woodburnei (Fig. 10.4), which is a derived feature not present in D. murgonensis (Fig. 10.5 and 10.6) or A. tirarensis (Fig. 10.7) but is present in plesiomorphic dasyuromorphians such as Joculusium muizoni Wroe, Reference Wroe2001b, and Barinya wangala (Fig. 10.8 and 10.9). Although they share multiple derived dental traits, the new taxa differ sufficiently from K. woodburnei to preclude congeneric assignment, instead suggesting a close familial relationship (Keeunidae n. fam.) within Keeunamorphia n. ord.
Dental morphology suggests a closer relationship between keeunamorphians and dasyuromorphians than with other marsupialiforms. Phantasmodon travouilloni n. gen. n. sp. shares five derived dental features with dasyuromorphians that are absent in peramelemorphians and notoryctemorphians: preparacrista contacts the lingual base of stB, not stA or the anterior base of stB (Archer, Reference Archer1976a); stD larger than stB on M1–3 (Cifelli, Reference Cifelli1993; Muirhead and Filan, Reference Muirhead and Filan1995; Wroe, Reference Wroe1996, Reference Wroe1997b); elongation of the metastylar corner on M1–3 (Archer, Reference Archer1976a); small stC closely twinned with stD on M1–3 (Godthelp et al., Reference Godthelp, Wroe and Archer1999); and presence of stE on M1–3 (Archer, Reference Archer1976a). Keeunamorphians and dasyuromorphians also retain plesiomorphic dental features absent in peramelemorphians: M1 preparacrista oriented perpendicular to the tooth-row axis (Archer, Reference Archer1976a); continuous V-shaped centrocrista that is level with ectoloph basin (Archer, Reference Archer1976a; Muirhead, Reference Muirhead1994; Muirhead and Filan, Reference Muirhead and Filan1995; Kear et al., Reference Kear, Aplin and Westerman2016; Warburton and Travouillon, Reference Warburton and Travouillon2016; Travouillon and Phillips, Reference Travouillon and Phillips2018); hypoconulid notch present on m2–4 (Archer, Reference Archer1976a; Muirhead and Wroe, Reference Muirhead and Wroe1998; Voss and Jansa, Reference Voss and Jansa2009; Beck et al., Reference Beck, Voss and Jansa2022); and termination of the cristid obliqua on m3 buccal to the metacristid carnassial notch (Archer, Reference Archer1976a; Kealy and Beck, Reference Kealy and Beck2017).
The presence of a posterior cingulid on m1–3 in keeunamorphians and dasyuromorphians is particularly noteworthy considering the absence of this feature was the only unambiguous craniodental synapomorphy of Marsupialia found by Beck et al. (Reference Beck, Voss and Jansa2022), with its reacquisition serving as a synapomorphy of Dasyuromorphia (Beck et al., Reference Beck, Voss and Jansa2022). The posterior cingulid can be interpreted in the following ways: it is a synapomorphy supporting a clade relationship between keeunamorphians and dasyuromorphians (suggested by number of dental similarities and the Bayesian phylogenetic analysis including sternbergiids and Pe. constans herein, but not by the other three phylogenetic analyses); it has re-evolved in these two lineages independently as it has in some didelphids (supported by both parsimony phylogenetic analyses and the Bayesian phylogenetic analyses excluding sternbergiids and Pe. constans herein); or keeunamorphians are not part of crown Marsupialia (not supported by morphological comparisons or phylogenetic studies herein). Although a relationship between keeunamorphians and dasyuromorphians received minor phylogenetic support when sternbergiids and herpetotheriids are included in the Bayesian analysis, such a conclusion otherwise lacks robust phylogenetic support from all analyses.
Keeunamorphians also share three derived dental features with peramelemorphians that are absent in dasyuromorphians and other australidelphian clades: lingual invasion of the centrocrista into the ectoloph; large metaconules (reduced in Yarala burchfieldi Muirhead and Filan, Reference Muirhead and Filan1995); and reduction of hypoconulids on all lower molars. Moreover, keeunamorphians and peramelemorphians share three plesiomorphic lower dental characters that are absent in dasyuromorphians but present in didelphids and some stem metatherians: m3 talonid wider than trigonid; talonids longitudinally unreduced relative to the trigonids; and large entoconids on m1–3 to approach paraconids in height.
Based on the dental features discussed above, keeunamorphians are dentally more similar to dasyuromorphians than to peramelemorphians in terms of apomorphies. However, the number of plesiomorphic features shared by members of all three orders suggest that dasyuromorphians and peramelemorphians evolved from a plesiomorphic keeunamorphian-like ancestor. Our phylogenetic analyses did not recover any synapomorphies supporting clade relationships between keeunamorphians and peramelemorphians, and showed only minor support for a clade relationship between keeunamorphians and dasyuromorphians in one of four analyses. Contrary to Murray and Megirian (Reference Murray and Megirian2006) that Keeuna woodburnei and Ankotarinja tirarensis are dasyurids, this study adds to the growing phylogenetic literature that recovers these taxa outside of both crown dasyurids and crown dasyuromorphians and is the first to place them alongside the new taxa and Djarthia murgonensis in a clade of their own.
Keeunamorphians share seven derived dental features with sternbergiid metatherians from Brazil’s early Eocene Itaborai Formation that have led previous authors to suggest assignment of Djarthia murgonensis, Ankotarinja tirarensis, and Keeuna woodburnei to the metatherian family Sternbergiidae (Oliveira, Reference Oliveira1998; Goin, Reference Goin, Jones, Dickman and Archer2003). The most noteworthy of these derived features is the shared presence of a central cusp on the upper molars, which is only observed in keeunamorphians, the bizarre peramelemorphian Numbigilga ernielundeliusi Beck et al., Reference Beck, Archer, Godthelp, Mackness, Hand and Muirhead2008a, sternbergiids (e.g., Carolopaulacoutoia itaboraiensis, Itaboraidelphys camposi, Didelphopsis cabrerai Paula Couto, Reference Paula Couto1952, and Di. n. sp.), some putative and unequivocal herpetotheriids (Maastrichidelphys meurismeti and species of Peratherium) and at least one alphadontid (i.e., Turgidodon rhaister Clemens, Reference Clemens1966) among tribosphenic marsupialiforms (Clemens, Reference Clemens1966; Cifelli, Reference Cifelli1990; Oliveira, Reference Oliveira1998; Godthelp et al., Reference Godthelp, Wroe and Archer1999; Goin and Candela, Reference Goin and Candela2004; Beck et al., Reference Beck, Archer, Godthelp, Mackness, Hand and Muirhead2008a; Hooker et al., Reference Hooker, Sanchez-Villagra, Goin, Simons, Attia and Seiffert2008; Oliveira and Goin, Reference Oliveira and Goin2011, Reference Oliveira, Goin and Cáceres2012; Carneiro and Oliveira, Reference Carneiro, Oliveira, Cáceres and Dickman2023; Gernelle et al., Reference Gernelle, Godinot, Marandat, Téodori, Ladevèze and Tabuce2025).
Keeunamorphians and sternbergiids share six other dental features considered derived among marsupialiforms (Godthelp et al., Reference Godthelp, Wroe and Archer1999), which led to the tentative assignment of some keeunamorphians to the metatherian family Sternbergiidae (Oliveira, Reference Oliveira1998; Goin, Reference Goin, Jones, Dickman and Archer2003): hypertrophy of the M4 preparacrista relative to that of the M3; marked reduction of the paracones relative to the metacones; dilambdodonty; stC present; slight expansion of the m3 talonid relative to that of the trigonid; and termination of the cristid obliqua buccal to the metacristid carnassial notch. However, more recent studies do not support a close relationship between keeunamorphians and sternbergiids (Oliveira and Goin, Reference Oliveira and Goin2011; Carneiro and Oliveira Reference Carneiro and Oliveira2017b, Reference Carneiro, Oliveira, Cáceres and Dickman2023; Rangel et al., Reference Rangel, Carneiro, Bergqvist, Oliveira and Babot2019). Similarly, our phylogenetic results do not support the assignment of Phantasmodon travouilloni n. gen. n. sp., Keeuna woodburnei, Ankotarinja tirarensis, or Djarthia murgonensis to the family Sternbergiidae Szalay, Reference Szalay1994, as proposed by Oliveira (Reference Oliveira1998) and Goin (Reference Goin, Jones, Dickman and Archer2003).
The maximum parsimony analysis that included members of Sternbergiidae and Herpetotheriidae (Fig. 6) suggests keeunamorphians may be more closely related to sternbergiids and herpetotheriids than to dasyuromorphians + peramelemorphians, microbiotherians, and didelphimorphians, but does not support a clade relationship with sternbergiids to the exclusion of herpetotheriids. However, the dated Bayesian total evidence analysis (Fig. 8) suggests that Oligo-Miocene keeunamorphians are closely related to dasyuromorphians, and that D. murgonensis is a stem marsupial alongside sternbergiids, with Pe. constans sister to all members of Marsupialia; a result congruent with the phylogenetic position of Herpetotheriidae within Metatheria (Sanchez-Villagra et al., Reference Sanchez-Villagra, Ladevèze, Horovitz, Argot and Hooker2007; Ladevèze et al., Reference Ladevèze, Selva and Muizon2020). This suggests more recent keeunamorphians are closely related to dasyuromorphians but that their earliest member, D. murgonensis, is a stem marsupial alongside sternbergiids, microbiotherians, and didelphimorphians.
Based on the results of this study, it remains unclear whether the presence of a central cusp on the upper molars in keeunamorphians, sternbergiids, some herpetotheriids, and some Cretaceous metatherians is the result of homoplasy or a plesiomorphy shared by these groups, which was subsequently lost by many different lineages independently. However, the central cusp has remained prominent for more than 35 million years in keeunamorphians (based on its presence in D. murgonensis and Phantasmodon travouilloni n. gen. n. sp.) and is not present in any other Australian metatherians known to the authors (except for in the bizarre peramelemorphian Numbigilga ernielundeliusi; Beck et al., Reference Beck, Archer, Godthelp, Mackness, Hand and Muirhead2008a). The fact that the central cusp occurs in three lineages (Keeunamorphia n. ord., Sternbergiidae, and Herpetotheriidae) during the early Paleocene to early Eocene, at a time where the Americas, Antarctica, and Australia shared land-routes and a number of metatherian taxa (Goin et al., Reference Goin, Candela and Abella2009; Sigé et al., Reference Sigé, Archer, Crochet, Godthelp, Hand and Beck2009; Beck, Reference Beck2012), suggests that it is a possible symplesiomorphy in keeunamorphians, sternbergiids, and herpetotheriids from some shared common ancestor that lived during the late Cretaceous to early Paleocene, which was subsequently lost by the Oligo-Miocene in all lineages except for Keeunamorphia n. ord. In addition to Turgidon, several Mesozoic taxa sometimes assigned to Alphadontidae (e.g., Albertatherium, Nortedelphys) have a “central cusp” positioned lingual to stylar cusp C. However, this cusp in putative alphadontids is likely not homologous with the central cusps in keeunamorphians, sternbergiids, and some herpetotheriids, which is situated just buccal of the centrocrista (Fox, Reference Fox1971; Johanson, Reference Johanson1995; Case et al., Reference Case, Goin and Woodburne2005; Carneiro and Oliveira, Reference Carneiro and Oliveira2017a). The results of this study do not strongly support a close phylogenetic relationship between Keeunamorphia n. ord., Sternbergiidae, and Herpetotheriidae, particularly due to the paucity of complete specimens attributable to these groups. However, the mixed results of the phylogenetic analyses herein means that the hypothesis of Oliveira (Reference Oliveira1998) and Goin (Reference Goin, Jones, Dickman and Archer2003) that keeunamorphians are sternbergiids cannot be confidently dismissed.
Biogeographical implications
The presence of Djarthia murgonensis in southeastern Queensland ca. 55 million years ago, Keeuna woodburnei and Ankotarinja tirarensis in the Etadunna Formation of South Australia ca. 26–23 million years ago, and Phantasmodon travouilloni n. gen. n. sp., Phantasmodon minuferox n. gen. n. sp., and Keeunidae n. fam. gen. and sp. indet. at Riversleigh, northwestern Queensland ca. 18 million years ago suggests this lineage, which may have Gondwanan origins among other stem metatherians (e.g., sternbergiids and herpetotheriids), survived on the Australian continent for at least 37 million years. Keeunamorphia n. ord. is therefore both the oldest and the longest persisting undoubted metatherian lineage in the Australian fossil record (unless Thylacotinga is in fact a stem diprotodontian; see Crichton et al., Reference Crichton, Beck, Couzens, Worthy, Camens and Prideaux2023). The presence of multiple keeunamorphian lineages during the late Oligocene suggests there was early radiation of the order sometime between the late Eocene and early Oligocene. By the Miocene, most small carnivorous niches were occupied by dasyuromorphians, peramelemorphians, or diminutive thylacoleonids. This may explain why keeunamorphians are relatively rare in the Miocene fossil record, particularly at Riversleigh, which otherwise exhibits an extremely rich fossil diversity for most metatherian lineages. Based on its relatively plesiomorphic dentition and diminutive size, P. travouilloni n. gen. n. sp. (and probably P. minuferox n. gen. n. sp. and Keeunidae n. fam. gen. and sp. indet.) was likely a relatively unspecialized insectivore (Fig. 11), a niche also proposed for other keeunamorphians (Archer, Reference Archer1976b; Godthelp et al., Reference Godthelp, Wroe and Archer1999). The extinction of keeunamorphians appears to have occurred at some point during the Miocene because there are no fossils attributable to this clade known from Plio-Pleistocene deposits.
Reconstruction of Phantasmodon travouilloni n. gen. n. sp. in the Early Miocene rainforests of northwestern Queensland. Illustration by Peter Schouten.

Figure 11. Long description
At the center, a small quadrupedal marsupial with a pointed snout, rounded ears, and dense gray-brown fur stands on a patch of overlapping green and gray leaves. Its long, thin tail extends diagonally from the lower left to the upper left. The animal’s head is angled to the right, showing sharp teeth and dark eyes. Surrounding the animal, the ground is covered with textured foliage in shades of green, yellow, and brown, with fern-like plants radiating outward. The background is a diffuse wash of pale greens and yellows, suggesting filtered rainforest light. The artist’s signature, Peter Schouten 2025, appears in the lower right corner.
Acknowledgments
We gratefully acknowledge the many individuals and institutions whose contributions made this research possible. We thank UNSW Sydney, Environment Australia, Queensland Parks and Wildlife Service, Queensland Museum, Outback at Isa, and A. Rackham for their valuable assistance, and the Waanyi Nation of north-western Queensland for their continued support of work on Country. We are also indebted to the Riversleigh World Heritage Advisory Committee and the Riversleigh Society Inc. for their ongoing commitment to the Riversleigh Project. Additional thanks are extended to Mount Isa City Council, the Lawn Hill Riversleigh Pastoral Company, and the numerous volunteers and collaborators from UNSW Sydney and other institutions who have contributed to this work. We further acknowledge the efforts of UNSW fossil preparators A. Gillespie, T. Myers, H. Godthelp, S. Williams, G. Hickey, and C. Nock for their careful preparation of the Riversleigh specimens. Computational resources were provided by the Katana high-performance computing facility, supported by Research Technology Services at UNSW Sydney.
Competing interests
The authors declare no competing interests.
Data availability statement
Data supporting the findings of this study are available within the paper and its Supplementary files. Supplementary file 1 includes all the specimens examined as part of this study. Supplementary file 2 includes the methods used for the maximum parsimony and Bayesian total evidence phylogenetic analyses. Supplementary file 3 includes the character analysis of new morphological characters added to the morphological matrices used in both phylogenetic analyses. Supplementary file 4 includes a list of morphological characters used to code the morphological and total evidence matrices for use in the phylogenetic analyses in this study. Supplementary file 5 contains the morphological matrix used in the maximum parsimony analysis in this study. Supplementary file 6 contains the molecular scaffold tree used in the maximum parsimony analysis to constrain the phylogenetic positions of modern taxa. Supplementary files 7 and 8 contain the strict consensus trees produced by the maximum parsimony analyses. Supplementary files 9 and 10 contain the matrix and MrBayes script for the tip-and-node calibrated Bayesian total evidence phylogenetic analyses. Supplementary files 11 and 12 contains the 50% majority-rule consensus trees produced by the dated Bayesian total evidence analyses. Supplementary file 13 contains the tip-and-node calibrations used in the dated Bayesian total evidence phylogenetic analyses. Supplementary file 14 contains the age ranges used in the tip-and-node date calibrations within the dated Bayesian total evidence phylogenetic analyses. New taxon names are registered in ZooBank under: urn:lsid:zoobank.org:pub:A2699457-5B2C-4F0C-B8B6-DD41401C3A17. All files associated with the phylogenetic analyses in this study (Supplementary files 1–14) are also available to download from Morphobank: https://www.morphobank.org/permalink/?P6097
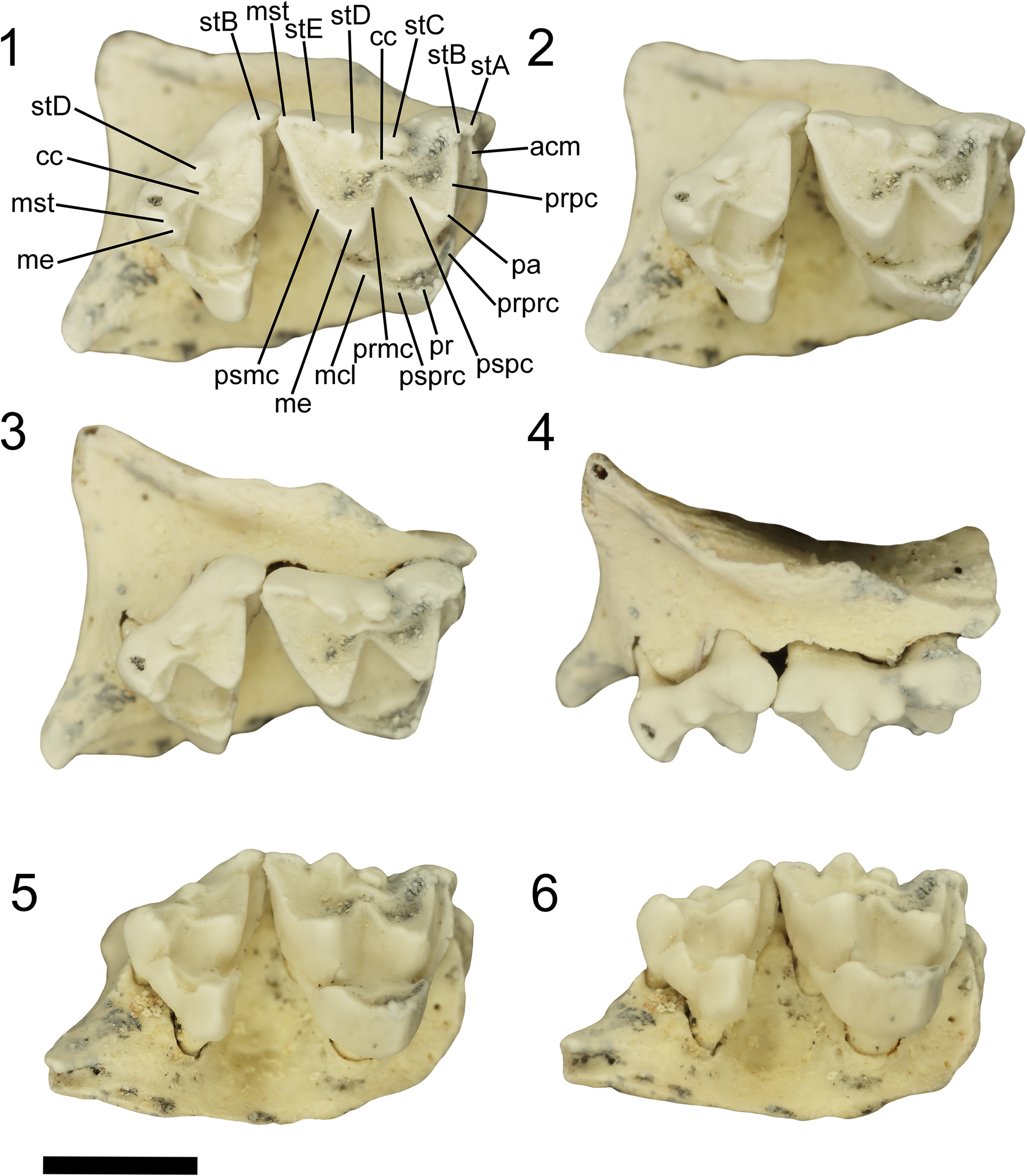
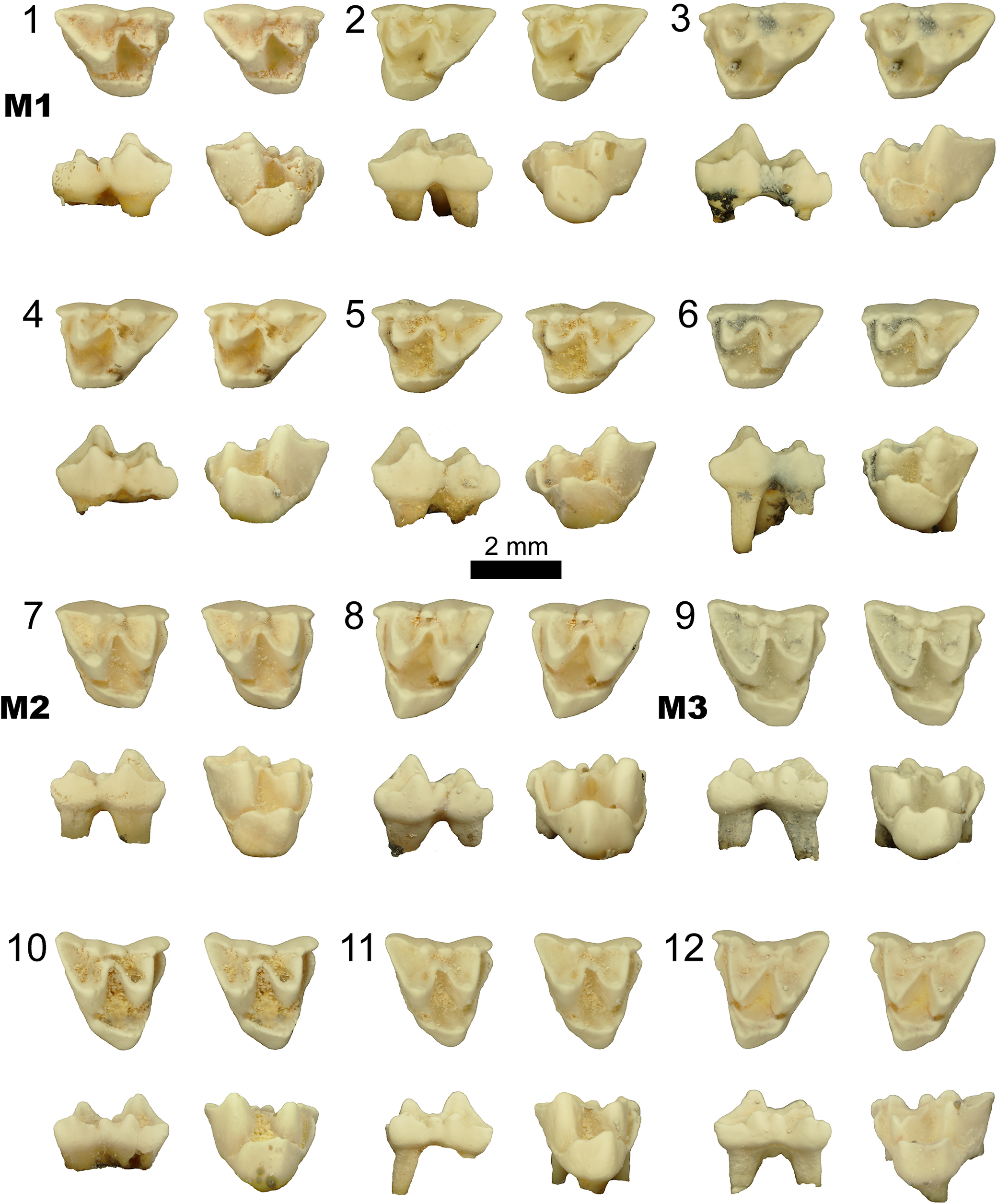
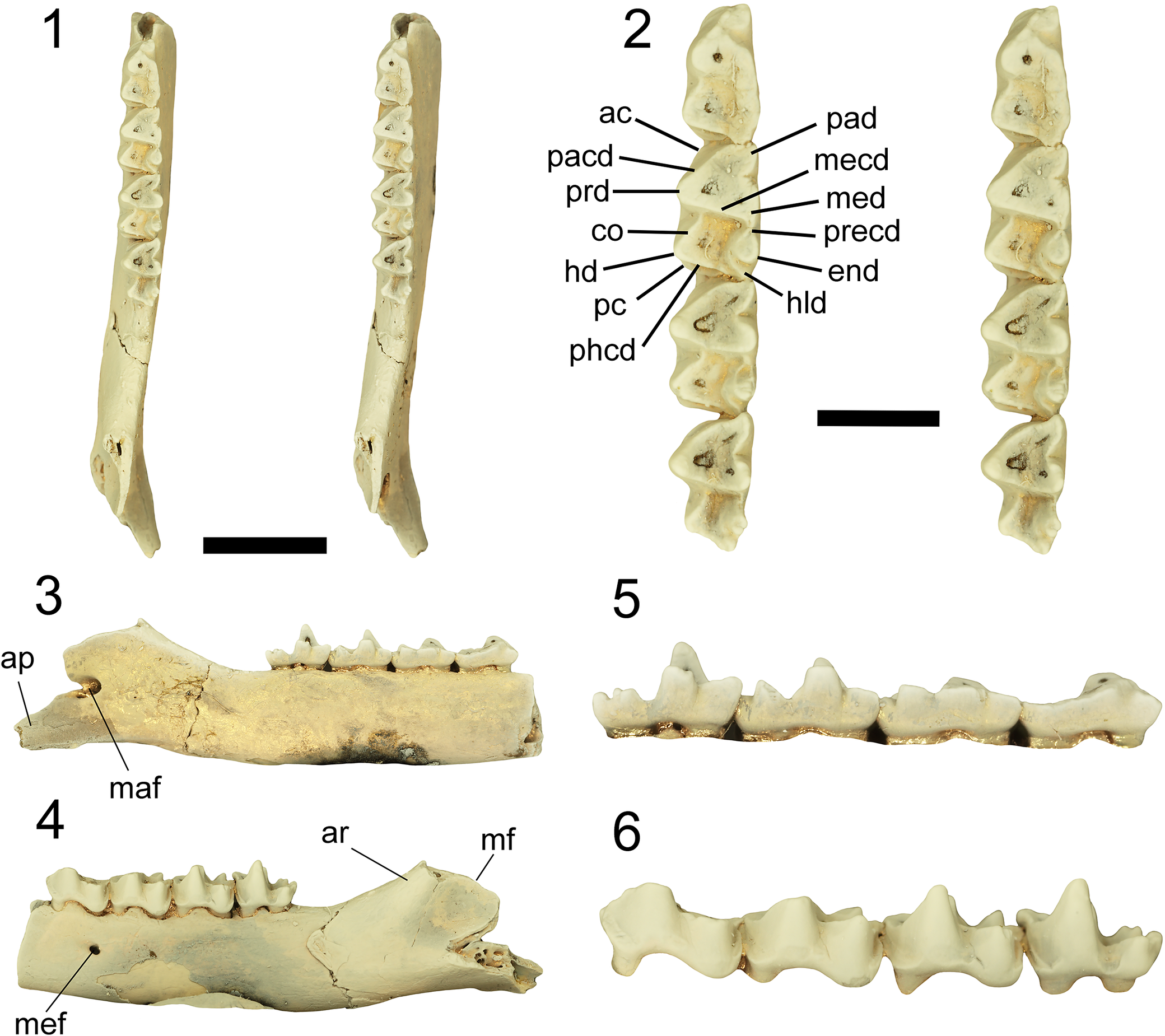
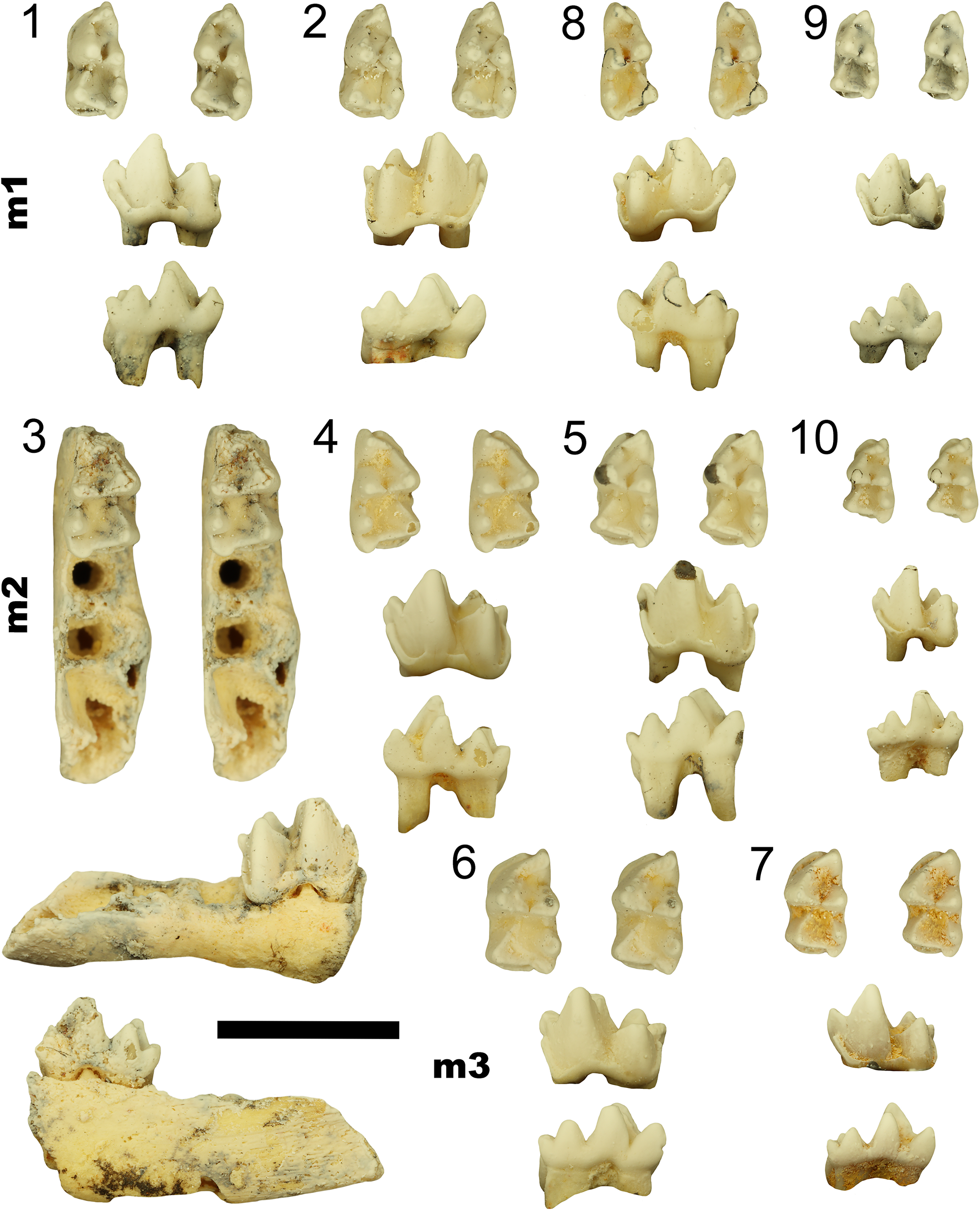
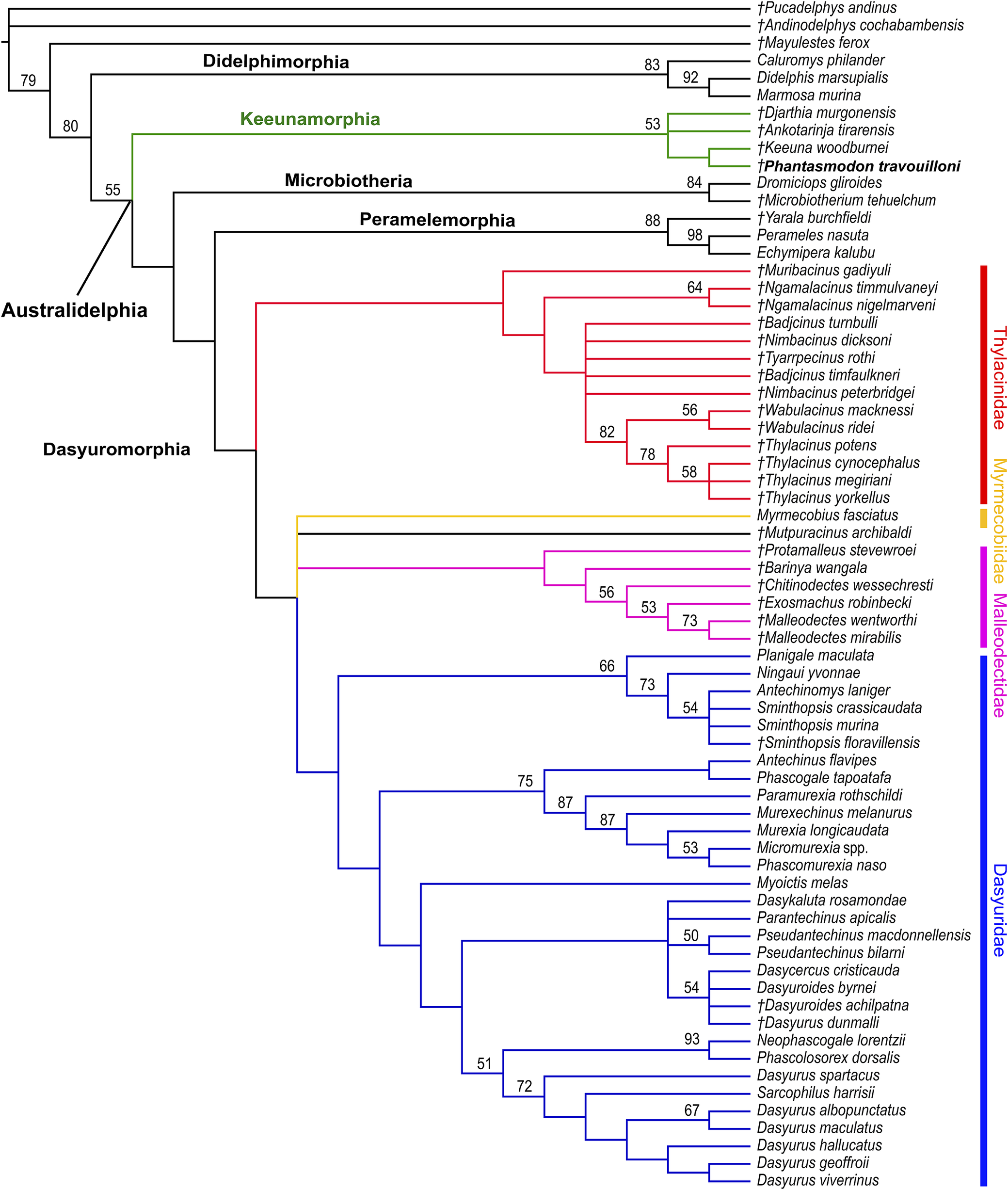
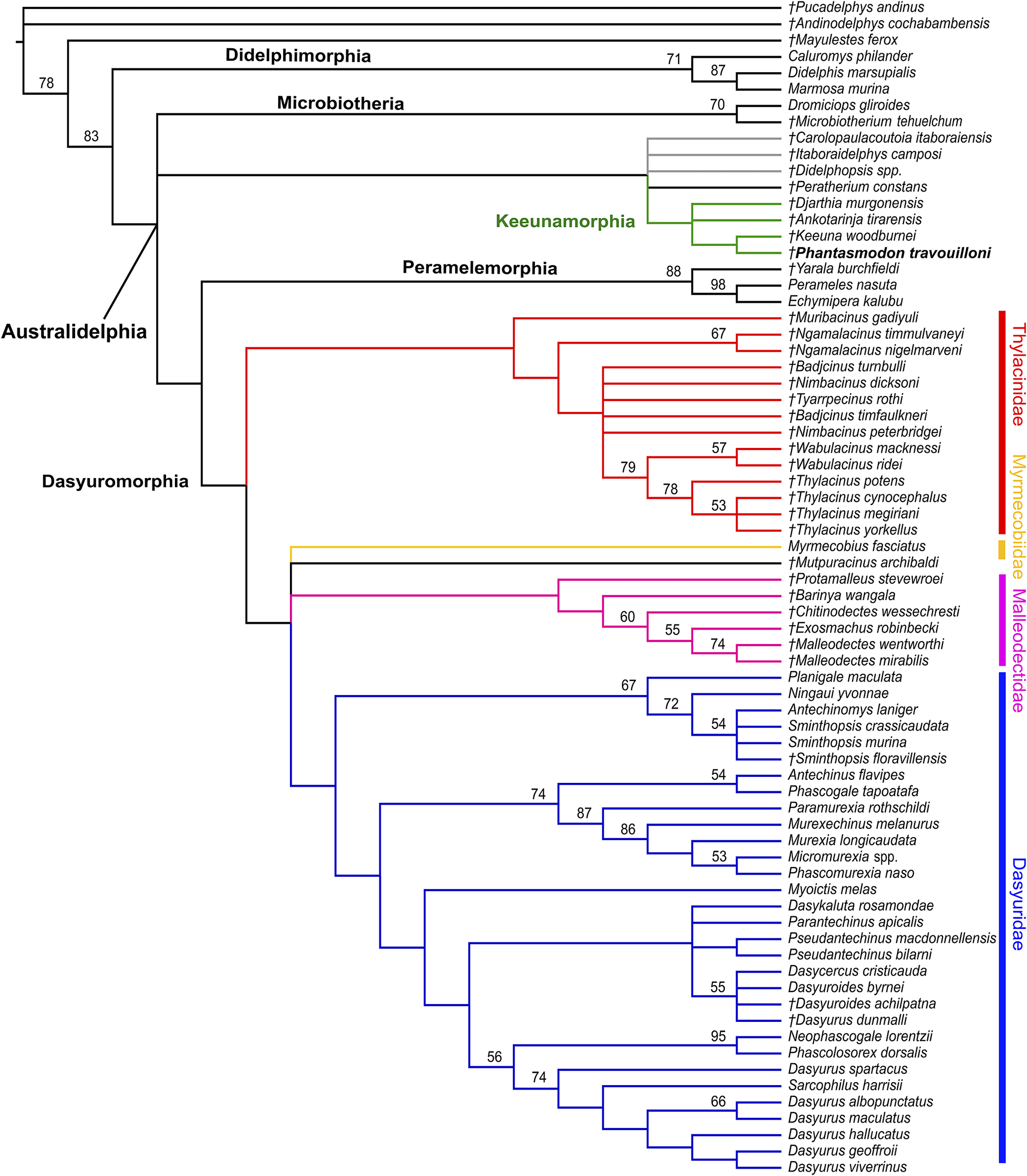
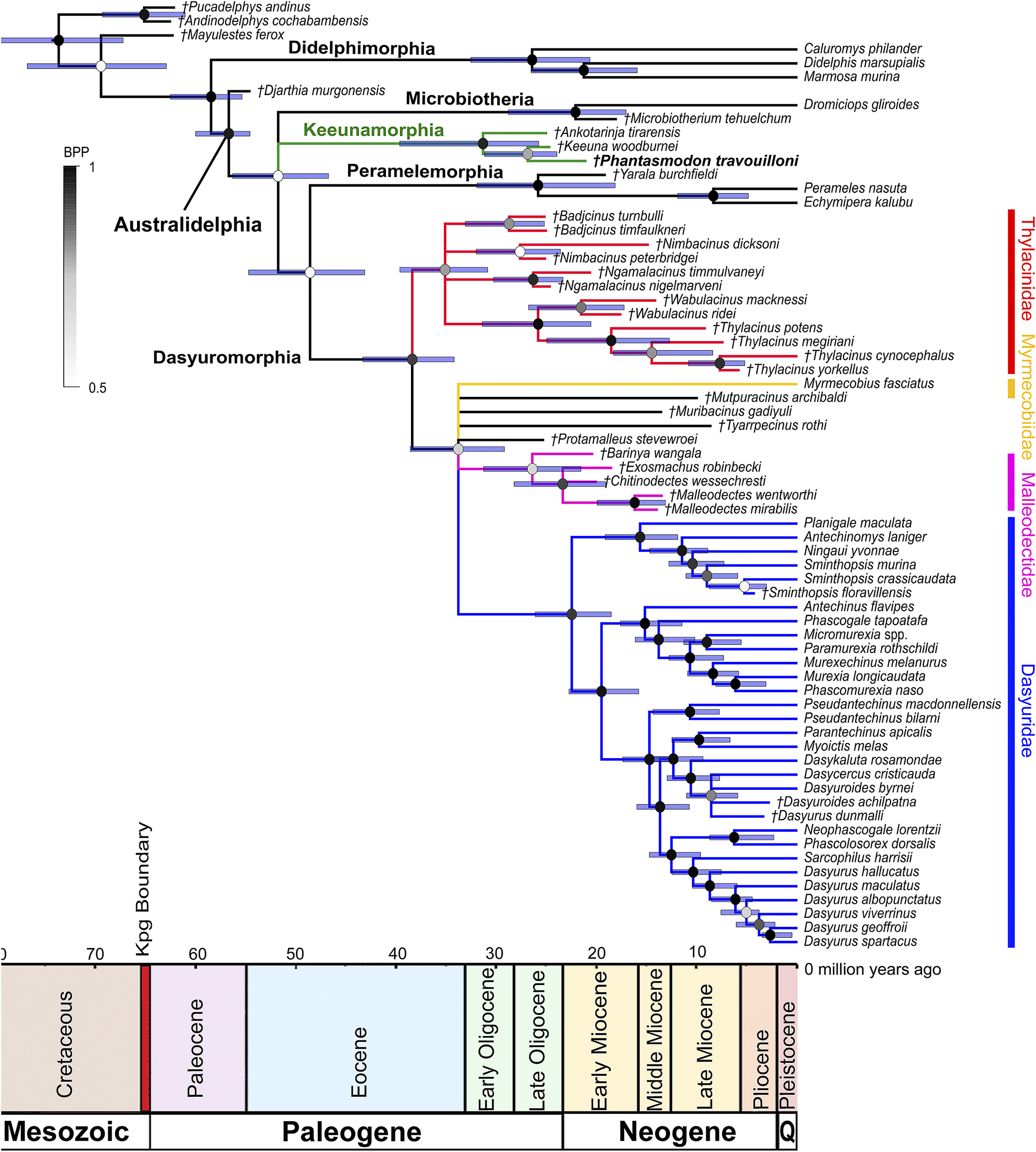
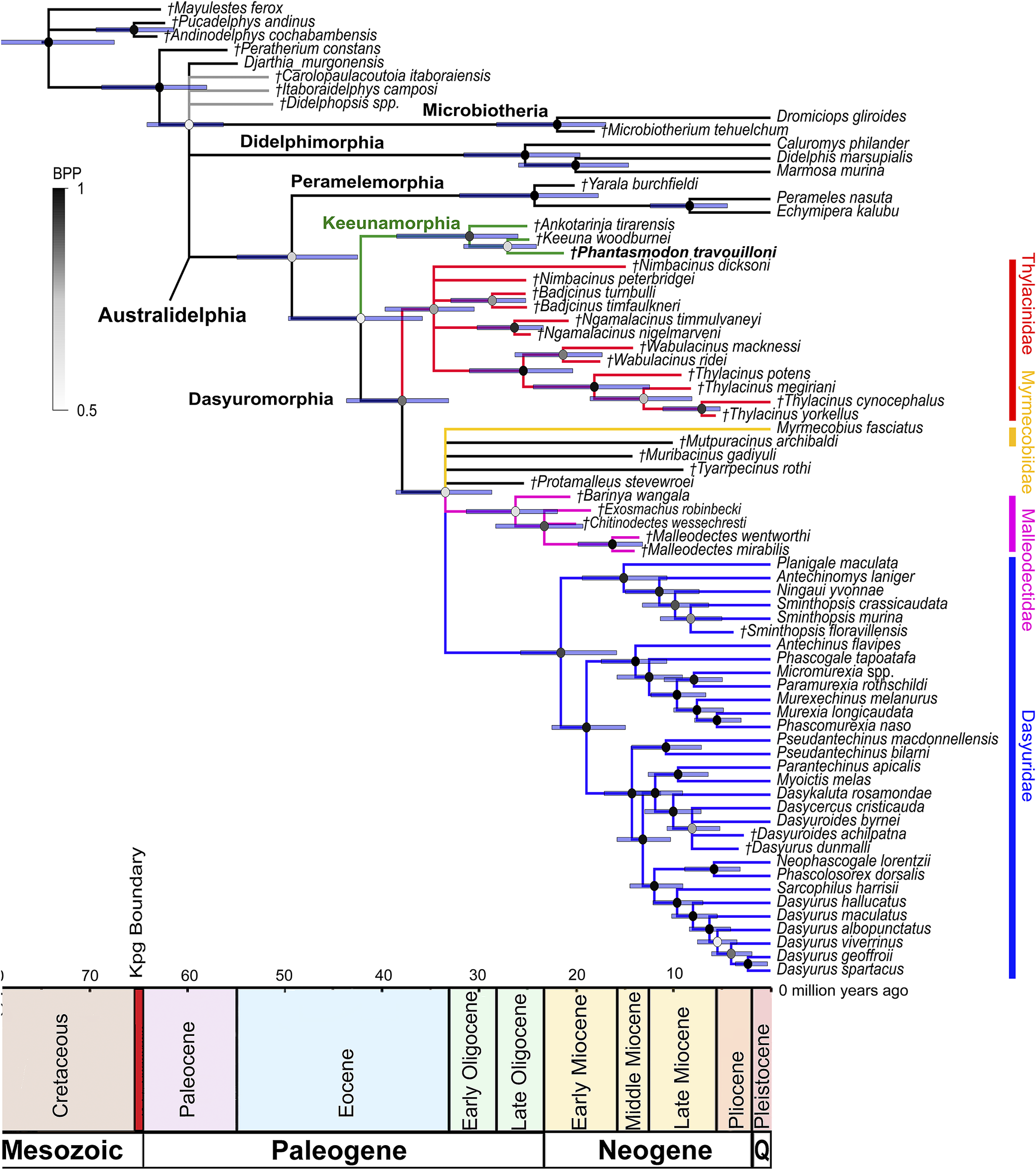
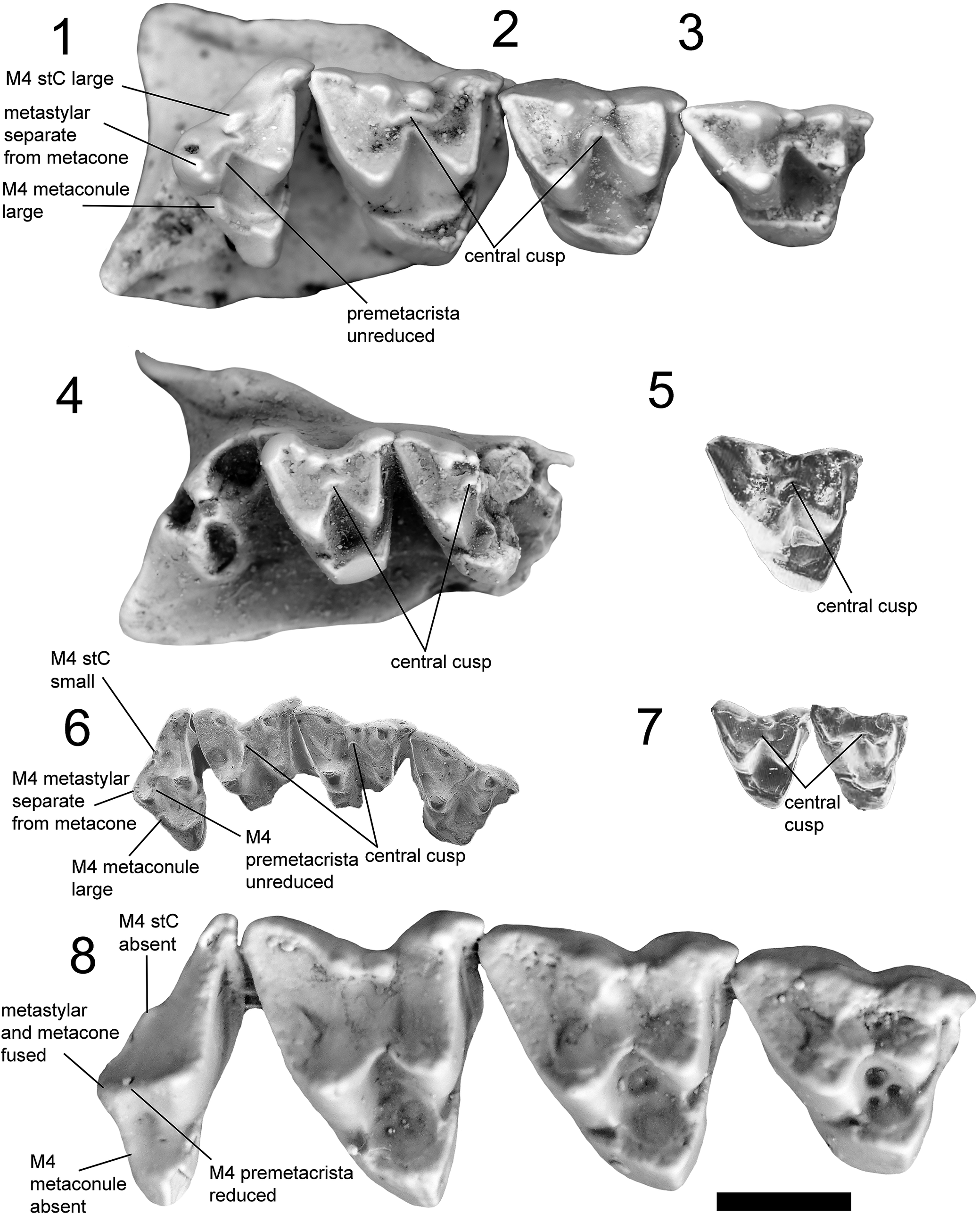
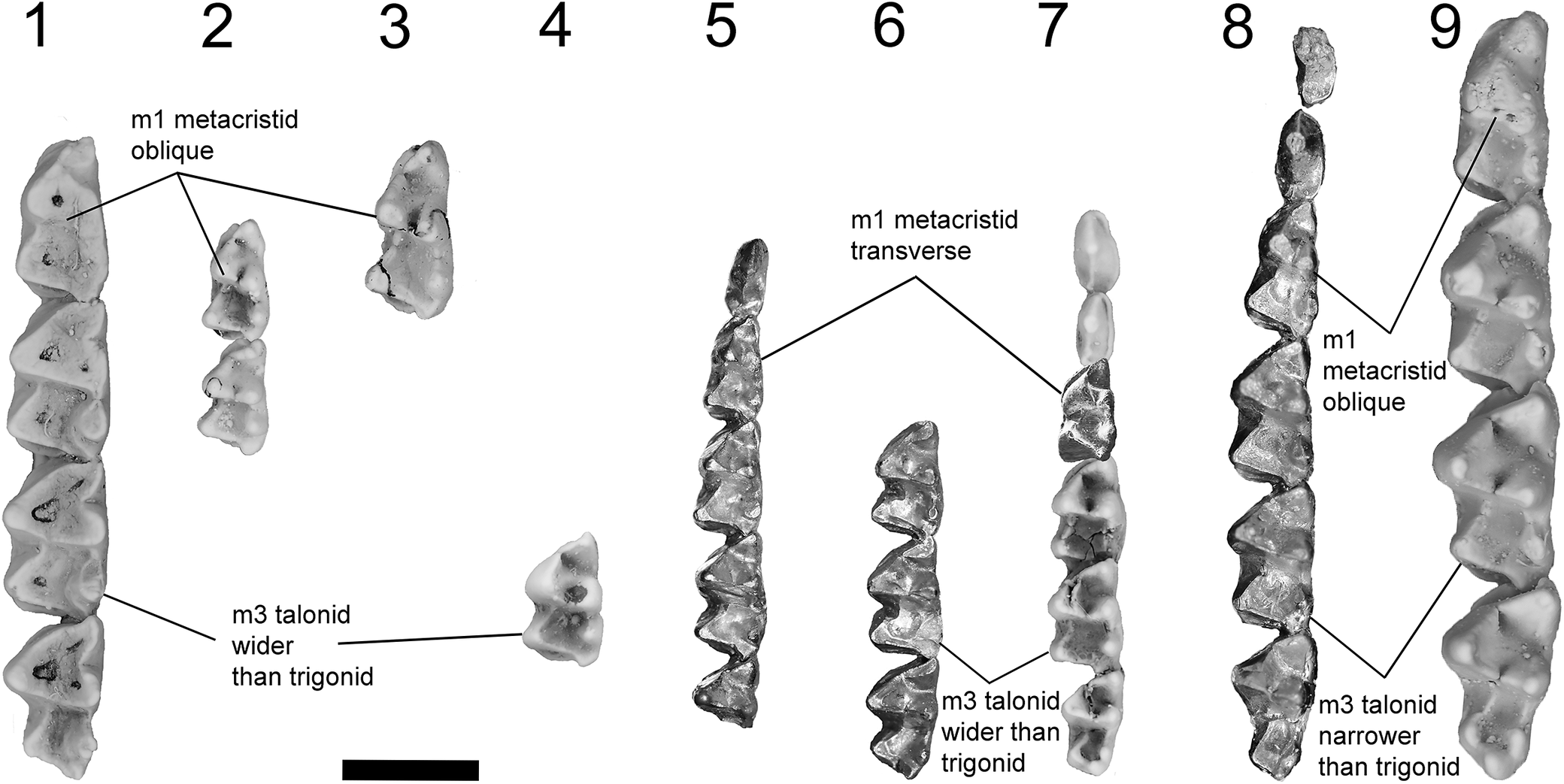
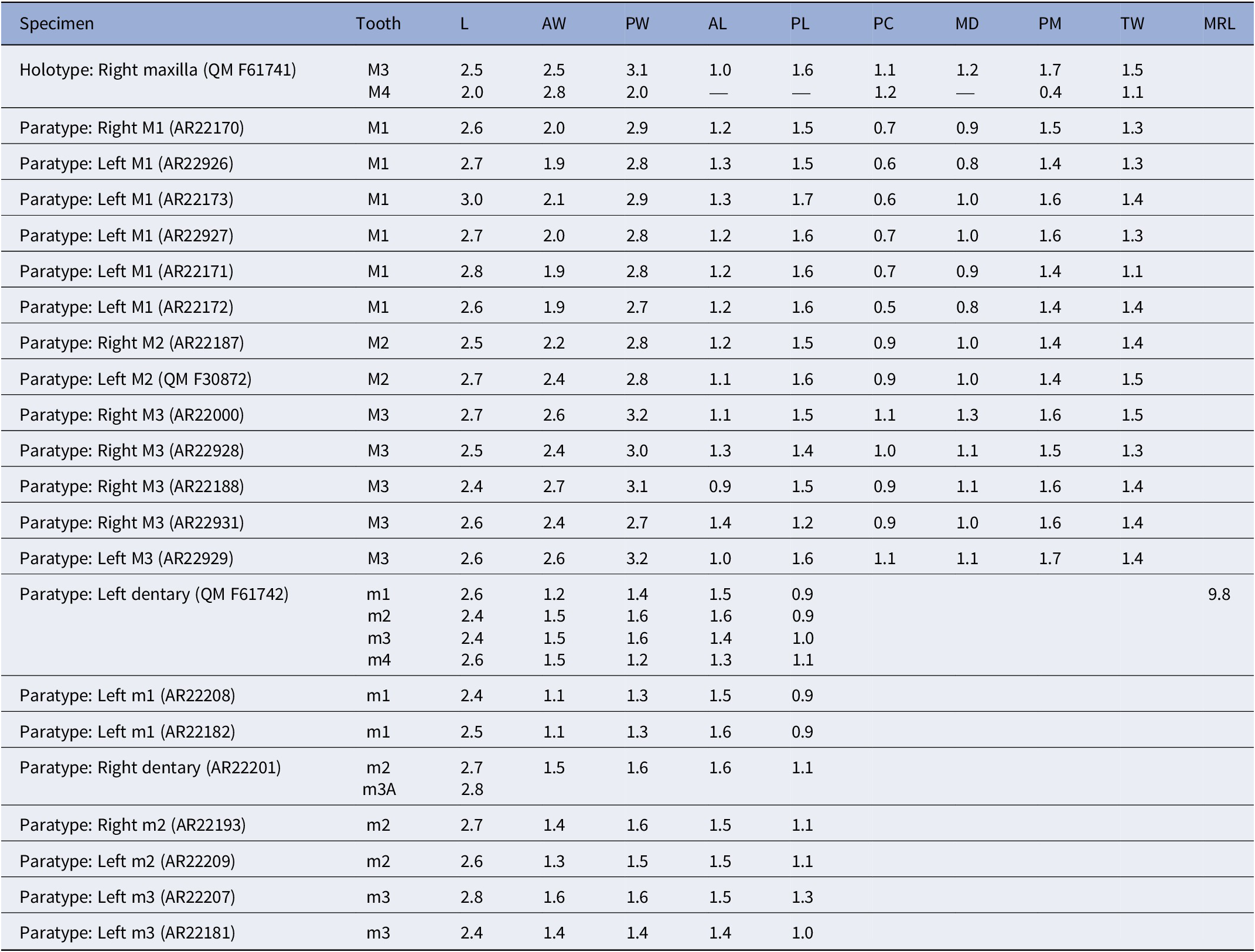
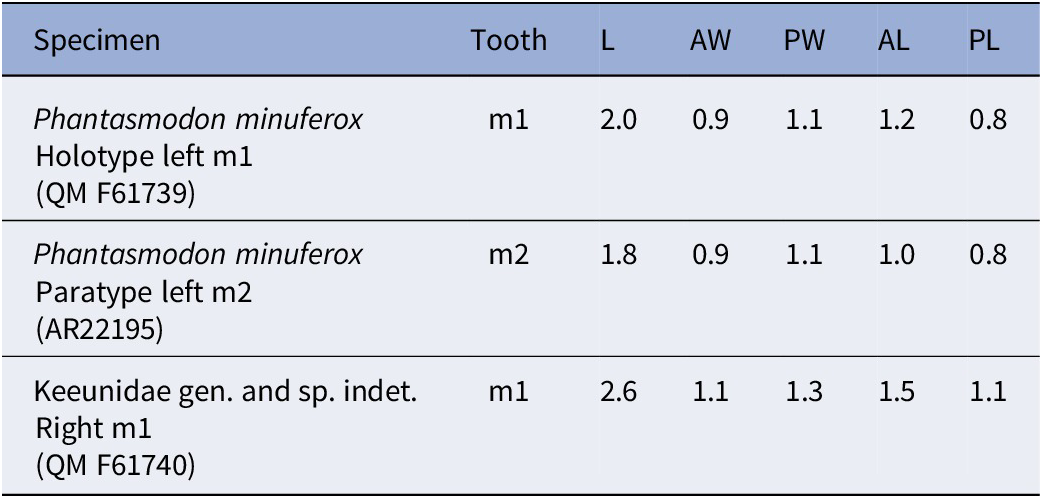
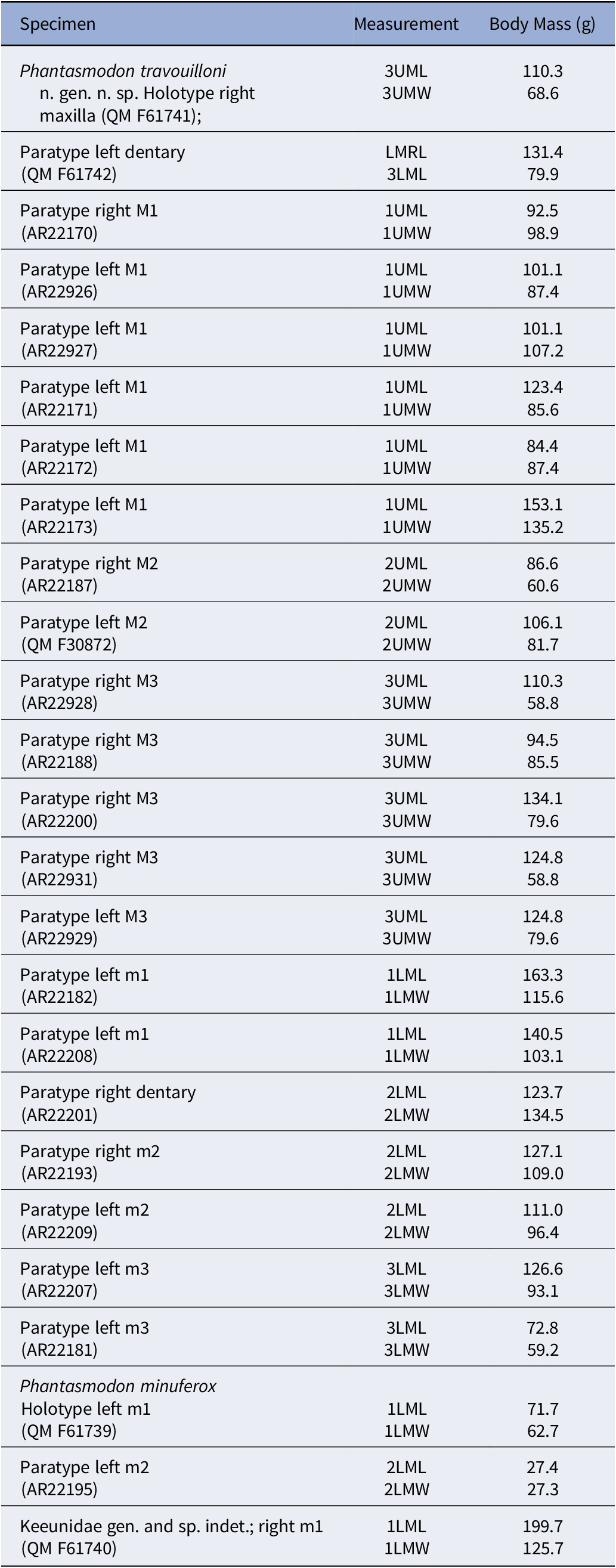


 Open access
Open access