Refine search
Actions for selected content:
10186 results in Genetics
The Molecular Pathology of Alcoholism. Molecular Medicine Series. Edited by T. Norman Palmer. Oxford University Press, 1991. 293 pages. Paperback £22.50. ISBN 0 19 261903 9.
-
- Journal:
- Genetical Research / Volume 59 / Issue 3 / June 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. 244-245
-
- Article
-
- You have access
- Export citation
The genetic basis of response in mouse lines divergently selected for body weight or fat content. I. The relative contributions of autosomal and sex-linked genes
-
- Journal:
- Genetical Research / Volume 62 / Issue 3 / December 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. 169-175
-
- Article
-
- You have access
- Export citation
Cytology of the XXY mouse
-
- Journal:
- Genetical Research / Volume 5 / Issue 2 / July 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 328-329
-
- Article
-
- You have access
- Export citation
An attempt to estimate the induction by X-rays of recessive lethal and visible mutations in mice
-
- Journal:
- Genetical Research / Volume 2 / Issue 2 / July 1961
- Published online by Cambridge University Press:
- 14 April 2009, pp. 296-305
-
- Article
-
- You have access
- Export citation
Cancer Cells 2/Oncogenes and Viral Genes. Edited by G. F. Vande Woude, A. J. Levine, W. C. Topp and J. D. Watson. New York: Cold Spring Harbor Laboratory. 1984. 650 pages (approx.). $65 U.S.A. elsewhere $78 ISBN 0 87969 168 7.
-
- Journal:
- Genetical Research / Volume 45 / Issue 2 / April 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 229-231
-
- Article
-
- You have access
- Export citation
GRH volume 28 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 28 / Issue 3 / December 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Lethal effect of attached-X deletion in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 13 / Issue 3 / June 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 325-327
-
- Article
-
- You have access
- Export citation
GRH volume 18 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 18 / Issue 1 / August 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Protein and nucleic acid metabolism in organs from mice selected for larger and smaller body size
-
- Journal:
- Genetical Research / Volume 22 / Issue 3 / December 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 255-278
-
- Article
-
- You have access
- Export citation
Inbreeding coefficients and coalescence times
-
- Journal:
- Genetical Research / Volume 58 / Issue 2 / October 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 167-175
-
- Article
-
- You have access
- Export citation
Genetic analysis of larval feeding behaviour in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 24 / Issue 2 / October 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 163-173
-
- Article
-
- You have access
- Export citation
Books received
-
- Journal:
- Genetical Research / Volume 55 / Issue 1 / February 1990
- Published online by Cambridge University Press:
- 14 April 2009, p. 69
-
- Article
-
- You have access
- Export citation
The epistasis of Adh and Gpdh allozymes and variation in the ethanol tolerance of Drosophila melanogaster larvae
-
- Journal:
- Genetical Research / Volume 52 / Issue 3 / December 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 179-184
-
- Article
-
- You have access
- Export citation
GRH volume 47 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 47 / Issue 2 / April 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
GRH volume 40 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 40 / Issue 1 / August 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
A model of mutation appropriate to estimate the number of electrophoretically detectable alleles in a finite population*
-
- Journal:
- Genetical Research / Volume 22 / Issue 2 / October 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 201-204
-
- Article
-
- You have access
- Export citation
-
A new model of mutational production of alleles was proposed which may be appropriate to estimate the number of electrophoretically detectable alleles maintained in a finite population. The model assumes that the entire allelic states are expressed by integers (…, A−1, A0, A1, …) and that if an allele changes state by mutation the change occurs in such a way that it moves either one step in the positive direction or one step in the negative direction (see also Fig. 1). It was shown that for this model the ‘effective’ number of selectively neutral alleles maintained in a population of the effective size Ne under mutation rate υ per generation is given by
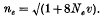
When 4Neυ is small, this differs little from the conventional formula by Kimura & Crow, i.e. ne = 1 + 4Neυ, but it gives a much smaller estimate than this when 4Neυ is large.
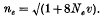
GRH volume 63 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 63 / Issue 1 / February 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Directional selection and the evolution of sex and recombination
-
- Journal:
- Genetical Research / Volume 61 / Issue 3 / June 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. 205-224
-
- Article
-
- You have access
- Export citation
GRH volume 13 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 13 / Issue 2 / April 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
A quick and efficient method for interruption of bacterial conjugation*
-
- Journal:
- Genetical Research / Volume 6 / Issue 2 / July 1965
- Published online by Cambridge University Press:
- 14 April 2009, pp. 300-303
-
- Article
-
- You have access
- Export citation
