Refine search
Actions for selected content:
10186 results in Genetics
X-linked Traits: A Catalog of Loci in Nonhuman Mammals. By R. Miller. 1990. Cambridge University Press. 198 pages. £25 ($39.50) hardback. ISBN 0 521 37389 1.
-
- Journal:
- Genetical Research / Volume 57 / Issue 3 / June 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 286-287
-
- Article
-
- You have access
- Export citation
Unequal crossing-over within the B duplication of Drosophila melanogaster: a molecular analysis
-
- Journal:
- Genetical Research / Volume 57 / Issue 2 / April 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 105-111
-
- Article
-
- You have access
- Export citation
Genotype, protein, phenotype relationships in self-incompatibility of Brassica
-
- Journal:
- Genetical Research / Volume 20 / Issue 2 / October 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 151-160
-
- Article
-
- You have access
- Export citation
Hemoglobin. By Richard E. Dickerson and Irving Geis. London: Addison-Wesley. £22.45. ISBN 0 8053 2411 9.
-
- Journal:
- Genetical Research / Volume 43 / Issue 1 / February 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 103-104
-
- Article
-
- You have access
- Export citation
Reversion analysis of [psi−] mutations in Saccharomyces cerevisiae
-
- Journal:
- Genetical Research / Volume 37 / Issue 2 / April 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 173-182
-
- Article
-
- You have access
- Export citation
Esterase polymorphism in insecticide susceptible populations of the mosquito Culex pipiens
-
- Journal:
- Genetical Research / Volume 67 / Issue 1 / February 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 19-26
-
- Article
-
- You have access
- Export citation
Mating types and mating-inducing factors (gamones) in the ciliate Euplotes patella syngen 2
-
- Journal:
- Genetical Research / Volume 46 / Issue 2 / October 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 125-132
-
- Article
-
- You have access
- Export citation
Gap Junctions. Edited by M. V. L Bennett, David C. Spray. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory. 1985. 409 pages. $70 in USA; $84 elsewhere. ISBN 0 87969 187 5.
-
- Journal:
- Genetical Research / Volume 48 / Issue 1 / August 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 55-56
-
- Article
-
- You have access
- Export citation
GRH volume 44 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 44 / Issue 2 / October 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Conventional and unconventional analysis of an inversion in Neurospora
-
- Journal:
- Genetical Research / Volume 40 / Issue 2 / October 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 175-190
-
- Article
-
- You have access
- Export citation
GRH volume 32 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 32 / Issue 2 / October 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Chromosomal locations of eleven Elytrigia elongata (= Agropyron elongatum) isozyme structural genes
-
- Journal:
- Genetical Research / Volume 41 / Issue 2 / April 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 181-202
-
- Article
-
- You have access
- Export citation
Interaction of selection and recombination in the fixation of negative-epistatic genes
-
- Journal:
- Genetical Research / Volume 67 / Issue 3 / June 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 257-269
-
- Article
-
- You have access
- Export citation
GRH volume 39 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 39 / Issue 1 / February 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
GRH volume 21 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 21 / Issue 1 / February 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Evidence of non-random X chromosome activity in the mouse
-
- Journal:
- Genetical Research / Volume 19 / Issue 3 / June 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 229-240
-
- Article
-
- You have access
- Export citation
On one of the formulae proposed for interpreting transduction data
-
- Journal:
- Genetical Research / Volume 5 / Issue 2 / July 1964
- Published online by Cambridge University Press:
- 14 April 2009, p. 333
-
- Article
-
- You have access
- Export citation
-
The paper on the analysis of data from transduction experiments with Salmonella typhimurium, which was published in Genetical Research (Dawson, 1963), unfortunately contained an error on p. 419. In calculating the limiting values X/Y, the formulae for d and d′ are correctly stated, but their solutions are transposed. The last and third last lines on p. 419 should therefore be interchanged. In the arithmetic example on pp. 420 and 421, a similar interchange of the calculations of d and d′ is necessary. The maximum value of a/b is then:
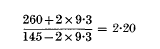
and the calculation of the minimum value should be:
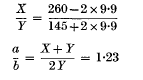
These values of 2·20 and 1·23 are to be compared with the values of 2·23 and 1·24 in the published paper.
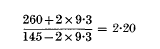
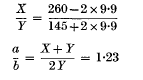
Books received
-
- Journal:
- Genetical Research / Volume 55 / Issue 3 / June 1990
- Published online by Cambridge University Press:
- 14 April 2009, p. 231
-
- Article
-
- You have access
- Export citation
Linkage disequilibrium due to random genetic drift*
-
- Journal:
- Genetical Research / Volume 13 / Issue 1 / February 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 47-55
-
- Article
-
- You have access
- Export citation
Cellular autonomy of hyperactivity in segmental X chromosomal aneuploids of Drosophila and dosage compensation
-
- Journal:
- Genetical Research / Volume 46 / Issue 1 / August 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 19-29
-
- Article
-
- You have access
- Export citation
