Refine search
Actions for selected content:
10186 results in Genetics
Direct and correlated responses to selection for plasma thyroxine levels in mice
-
- Journal:
- Genetical Research / Volume 38 / Issue 2 / October 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 157-169
-
- Article
-
- You have access
- Export citation
A world-wide survey of genetic variation in the yellow fever mosquito, Aedes aegypti*
-
- Journal:
- Genetical Research / Volume 34 / Issue 3 / December 1979
- Published online by Cambridge University Press:
- 14 April 2009, pp. 215-229
-
- Article
-
- You have access
- Export citation
GRH volume 52 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 52 / Issue 3 / December 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Inbreeding depression and heterosis of litter size in mice
-
- Journal:
- Genetical Research / Volume 1 / Issue 2 / July 1960
- Published online by Cambridge University Press:
- 14 April 2009, pp. 262-274
-
- Article
-
- You have access
- Export citation
Translational Control. Edited by J. W. B. Hershey, M. B. Mathews and N. Sonenberg. Cold Spring Harbor Laboratory Press, 1996. 794 pages. Price $95 cloth. ISBN 0 87969 458 0.
-
- Journal:
- Genetical Research / Volume 68 / Issue 3 / December 1996
- Published online by Cambridge University Press:
- 14 April 2009, p. 261
-
- Article
-
- You have access
- Export citation
A mathematical study on the distribution of the number of repeated genes per chromosome
-
- Journal:
- Genetical Research / Volume 38 / Issue 1 / August 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 97-102
-
- Article
-
- You have access
- Export citation
Fixation time of overdominant alleles influenced by random fluctuation of selection intensity*
-
- Journal:
- Genetical Research / Volume 20 / Issue 1 / August 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-7
-
- Article
-
- You have access
- Export citation
-
It was demonstrated that the number of generations until fixation or loss of an overdominant alleles is influenced by random fluctuation of selection coefficients. When 2
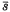 < Vs, where is the mean selection coefficient against either homozygote and Vs is the between -generation variance of the selection coefficient, overdominance generally accelerates rather than retards fixation of segregating alleles. This finding should have important bearing on our consideration of the behaviour of polymorphic variants which are nearly neutral but have very slight overdominance. When the population size (Ne) is extremely large, not only Ne but also /Vs have to be considered in discussing the effectiveness of overdominance.
< Vs, where is the mean selection coefficient against either homozygote and Vs is the between -generation variance of the selection coefficient, overdominance generally accelerates rather than retards fixation of segregating alleles. This finding should have important bearing on our consideration of the behaviour of polymorphic variants which are nearly neutral but have very slight overdominance. When the population size (Ne) is extremely large, not only Ne but also /Vs have to be considered in discussing the effectiveness of overdominance.
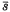 < V
< VRecombination during blocked chromosome replication in temperature-sensitive strains of Ustilago maydis
-
- Journal:
- Genetical Research / Volume 19 / Issue 2 / April 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 145-155
-
- Article
-
- You have access
- Export citation
Genome evolution in mosquitoes: intraspecific and interspecific variation in repetitive DNA amounts and organization
-
- Journal:
- Genetical Research / Volume 51 / Issue 3 / June 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 185-196
-
- Article
-
- You have access
- Export citation
A study on simultaneous conversions in linked genes in Ascobolus immersus
-
- Journal:
- Genetical Research / Volume 10 / Issue 2 / October 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 121-126
-
- Article
-
- You have access
- Export citation
On the nonautonomy of the small-kernel phenotype produced by B–A translocations in maize
-
- Journal:
- Genetical Research / Volume 36 / Issue 1 / August 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 111-116
-
- Article
-
- You have access
- Export citation
Sex Determination in Mouse and Man. Edited by Anne McLaren and M. A. Ferguson Smith. London: The Royal Society, 8 Carlton House Terrace, London SW1Y 5AG. 1988. 158 pages. Price including packing and postage £35.00 (UK) £37.60 (Overseas). ISBN 085403 369 6.
-
- Journal:
- Genetical Research / Volume 54 / Issue 1 / August 1989
- Published online by Cambridge University Press:
- 14 April 2009, p. 80
-
- Article
-
- You have access
- Export citation
Strain variation in spermatozoal glycosidases in inbred mice
-
- Journal:
- Genetical Research / Volume 32 / Issue 2 / October 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. 183-193
-
- Article
-
- You have access
- Export citation
Books Received
-
- Journal:
- Genetical Research / Volume 66 / Issue 2 / October 1995
- Published online by Cambridge University Press:
- 14 April 2009, p. 187
-
- Article
-
- You have access
- Export citation
A genetic analysis of CRM negative mutants at the td locus of Neurospora crassa
-
- Journal:
- Genetical Research / Volume 4 / Issue 3 / November 1963
- Published online by Cambridge University Press:
- 14 April 2009, pp. 470-479
-
- Article
-
- You have access
- Export citation
Direct determination of retrotransposon transposition rates in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 63 / Issue 2 / April 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 139-144
-
- Article
-
- You have access
- Export citation
Mutagenic action of nitrous acid on Aspergillus nidulans
-
- Journal:
- Genetical Research / Volume 3 / Issue 2 / July 1962
- Published online by Cambridge University Press:
- 14 April 2009, pp. 303-314
-
- Article
-
- You have access
- Export citation
Preferential transmission in wild common shrews (Sorex araneus), Heterozygous for Robertsonian rearrangements
-
- Journal:
- Genetical Research / Volume 47 / Issue 2 / April 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 147-148
-
- Article
-
- You have access
- Export citation
Erratum
-
- Journal:
- Genetical Research / Volume 50 / Issue 1 / August 1987
- Published online by Cambridge University Press:
- 14 April 2009, p. 77
-
- Article
-
- You have access
- Export citation
Dilution of gene products in the cytoplasm of heterokaryons in Coprinus lagopus
-
- Journal:
- Genetical Research / Volume 9 / Issue 1 / February 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 63-71
-
- Article
-
- You have access
- Export citation
