Refine search
Actions for selected content:
10186 results in Genetics
A theoretical analysis of linkage disequilibrium produced by genetic drift in Drosophila populations
-
- Journal:
- Genetical Research / Volume 48 / Issue 3 / December 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 161-166
-
- Article
-
- You have access
- Export citation
Methods for the isolation and analysis of plasmodial mutants in Physarum polycephalum
-
- Journal:
- Genetical Research / Volume 24 / Issue 2 / October 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 175-187
-
- Article
-
- You have access
- Export citation
Low rates of proviral integration in SWR/J–RF/J hybrid mice
-
- Journal:
- Genetical Research / Volume 66 / Issue 1 / August 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 45-52
-
- Article
-
- You have access
- Export citation
GRH volume 63 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 63 / Issue 2 / April 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b4
-
- Article
-
- You have access
- Export citation
Books received
-
- Journal:
- Genetical Research / Volume 28 / Issue 3 / December 1976
- Published online by Cambridge University Press:
- 14 April 2009, p. 309
-
- Article
-
- You have access
- Export citation
Estimating the proportion of neutral mutants
-
- Journal:
- Genetical Research / Volume 50 / Issue 2 / October 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 155-163
-
- Article
-
- You have access
- Export citation
-
Kimura used the heterozygosity and the number of low-frequency alleles to estimate that about 14% of mutations are selectively neutral. The method is shown to be subject to biases and to disruption due to bottleneck effects. Let deleterious alleles have selective disadvantage, s, compared with neutral alleles and let Ne denote the effective diploid population size. The estimator,
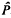 , of the proportion of neutral alleles is positively biased if (roughly) 4NeS < 25 or if 4Nes > 200. In the former case, one cannot adequately detect the different influences of deleterious and neutral alleles, whereas in the latter case, deleterious alleles will rarely appear in the sample. These difficulties cause the biases in , and are likely to cause similar biases for any estimation method based solely on allele frequencies. There is substantial sampling variability in in cases of practical interest, when data from 11 loci, or even as many as 31 loci, are pooled. If there has been a recent contraction in population size, will be positively biased, often yielding values greater than 1 or even being infinite. But after a recent expansion in population size, the heterozygosity will not have made as quick an increase and will be negatively biased. Population expansion alone can produce values close to those observed by Kimura, even if all alleles are neutral. In an appendix, a new method for simulating samples of neutral and deleterious genes is described.
, of the proportion of neutral alleles is positively biased if (roughly) 4NeS < 25 or if 4Nes > 200. In the former case, one cannot adequately detect the different influences of deleterious and neutral alleles, whereas in the latter case, deleterious alleles will rarely appear in the sample. These difficulties cause the biases in , and are likely to cause similar biases for any estimation method based solely on allele frequencies. There is substantial sampling variability in in cases of practical interest, when data from 11 loci, or even as many as 31 loci, are pooled. If there has been a recent contraction in population size, will be positively biased, often yielding values greater than 1 or even being infinite. But after a recent expansion in population size, the heterozygosity will not have made as quick an increase and will be negatively biased. Population expansion alone can produce values close to those observed by Kimura, even if all alleles are neutral. In an appendix, a new method for simulating samples of neutral and deleterious genes is described.
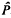 , of the proportion of neutral alleles is positively biased if (roughly) 4N
, of the proportion of neutral alleles is positively biased if (roughly) 4NEstimating variance effect of QTL: an important prospect to increase the resolution power of interval mapping
-
- Journal:
- Genetical Research / Volume 67 / Issue 2 / April 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 187-194
-
- Article
-
- You have access
- Export citation
-
Equal variances within quantitative trait locus (QTL) groups in the segregating population are a usual simplifying assumption in QTL mapping. The objective of this paper is to demonstrate the advantages of taking into account potential variance effect of QTLs within the framework of standard interval mapping approach. Using backcross case as an example, we show that the resolution power of the analysis may be increased in the presence of variance effect, if the latter is allowed for in the model. For a putative QTL (say, A/a) one can compare two situations, (i)
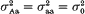 and (ii)
and (ii) 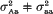 . It was found that, if the variance effect of A/a is large enough, then in spite of the necessity to evaluate an increased number of parameters, the more correctly specified model provides an increase in the resolution power, as compared to the situation (i). This is not unexpected, if either
. It was found that, if the variance effect of A/a is large enough, then in spite of the necessity to evaluate an increased number of parameters, the more correctly specified model provides an increase in the resolution power, as compared to the situation (i). This is not unexpected, if either 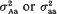 in (ii) is lower than
in (ii) is lower than 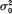 from (i). But our conclusion holds even if
from (i). But our conclusion holds even if 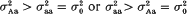 . These advantages are illustrated on sweet corn data data (F3 families of F2 genotypes). In particular, the log-likelihood test statistics and the parameter estimates obtained for a QT locus in the distal region of chromosome 2 show that the allele enhancing the trait is recessive over the opposite allele simultaneously for the mean value and variance.
. These advantages are illustrated on sweet corn data data (F3 families of F2 genotypes). In particular, the log-likelihood test statistics and the parameter estimates obtained for a QT locus in the distal region of chromosome 2 show that the allele enhancing the trait is recessive over the opposite allele simultaneously for the mean value and variance.
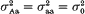 and (ii)
and (ii) 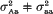 . It was found that, if the variance effect of A/a is large enough, then in spite of the necessity to evaluate an increased number of parameters, the more correctly specified model provides an increase in the resolution power, as compared to the situation (i). This is not unexpected, if either
. It was found that, if the variance effect of A/a is large enough, then in spite of the necessity to evaluate an increased number of parameters, the more correctly specified model provides an increase in the resolution power, as compared to the situation (i). This is not unexpected, if either 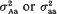 in (ii) is lower than
in (ii) is lower than 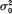 from (i). But our conclusion holds even if
from (i). But our conclusion holds even if 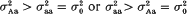 . These advantages are illustrated on sweet corn data data (F
. These advantages are illustrated on sweet corn data data (FGRH volume 15 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 15 / Issue 1 / February 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b3
-
- Article
-
- You have access
- Export citation
Surface exclusion by ColV–K94
-
- Journal:
- Genetical Research / Volume 21 / Issue 3 / June 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 297-299
-
- Article
-
- You have access
- Export citation
GRH volume 64 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 64 / Issue 1 / August 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Practical Developments in Inherited Metabolic Disease: DNA Analysis, Phenylketonuria and Screening for Congenital Adrenal Hypoplasia. Edited by G. M. Addison, R. A. Harkness, D. M. Isherwood and R. J. Pollitt. MTP Press Limited. 1986. 335 pages. £50.00. ISBN 0 85200 690 X.
-
- Journal:
- Genetical Research / Volume 49 / Issue 2 / April 1987
- Published online by Cambridge University Press:
- 14 April 2009, p. 177
-
- Article
-
- You have access
- Export citation
Four new half-and-half mosaic fowls
-
- Journal:
- Genetical Research / Volume 1 / Issue 2 / July 1960
- Published online by Cambridge University Press:
- 14 April 2009, pp. 275-287
-
- Article
-
- You have access
- Export citation
A genetic method for measuring non-disjunction in mice with Robertsonian translocations
-
- Journal:
- Genetical Research / Volume 26 / Issue 3 / December 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 283-295
-
- Article
-
- You have access
- Export citation
Regulation of enzyme activities in Drosophila: II. Characterization of enzyme responses in aneuploid flies
-
- Journal:
- Genetical Research / Volume 24 / Issue 1 / August 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 73-80
-
- Article
-
- You have access
- Export citation
GRH volume 50 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 50 / Issue 3 / December 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
GRH volume 67 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 67 / Issue 3 / June 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Properties of the R factor R144drd3 in Klebsiella pneumoniae strain M5a1
-
- Journal:
- Genetical Research / Volume 25 / Issue 3 / June 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 327-338
-
- Article
-
- You have access
- Export citation
GRH volume 31 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 31 / Issue 2 / April 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Cancer Cytogenetics. By Sverre Heim and Felix Mitelman. New York: Alan R. Liss, 1987; 2nd printing 1989. 309 pages. $35.00. ISBN 0 8451 4239 9. - Catalog of Chromosome Aberrations in Cancer. By Felix Mitelman. New York: Alan R. Liss. 3rd edition1988. 1146 pages $165.00. ISBN 0 8451 4248 8.
-
- Journal:
- Genetical Research / Volume 55 / Issue 2 / April 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 134-135
-
- Article
-
- You have access
- Export citation
Mutant with heat-sensitive capacity for phagocytosis in tetrahymena: isolation and genetic characterization
-
- Journal:
- Genetical Research / Volume 26 / Issue 1 / August 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 11-19
-
- Article
-
- You have access
- Export citation
