Refine search
Actions for selected content:
10186 results in Genetics
An exact test for neutrality based on the Ewens sampling distribution
-
- Journal:
- Genetical Research / Volume 64 / Issue 1 / August 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 71-74
-
- Article
-
- You have access
- Export citation
GRH volume 22 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 22 / Issue 3 / December 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Erratum
-
- Journal:
- Genetical Research / Volume 41 / Issue 1 / February 1983
- Published online by Cambridge University Press:
- 14 April 2009, p. 103
-
- Article
-
- You have access
- Export citation
-
Genet. Res., Camb. (1982), 39, pp. 157–168
The relationship between heterochromatic homology and meiotic segregation of compound second autosomes during spermatogenesis in Drosophila melanogaster
By Arthur J. Hilliker, David G. Holm and R. Appels
Several errors occurred in the headings of Table 4, on p. 162: the table should read as follows:


Triplications and the problem of non-homologous crossing-over
-
- Journal:
- Genetical Research / Volume 11 / Issue 2 / April 1968
- Published online by Cambridge University Press:
- 14 April 2009, pp. 201-208
-
- Article
-
- You have access
- Export citation
Genetics of sexual isolation in females of the Drosophila simulans species complex
-
- Journal:
- Genetical Research / Volume 60 / Issue 1 / August 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. 25-31
-
- Article
-
- You have access
- Export citation
Estimating the covariance structure of traits during growth and ageing, illustrated with lactation in dairy cattle
-
- Journal:
- Genetical Research / Volume 64 / Issue 1 / August 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 57-69
-
- Article
-
- You have access
- Export citation
GRH volume 56 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 56 / Issue 1 / August 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b3
-
- Article
-
- You have access
- Export citation
Fixation probability of a mutant influenced by random fluctuation of selection intensity*
-
- Journal:
- Genetical Research / Volume 19 / Issue 1 / February 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 33-38
-
- Article
-
- You have access
- Export citation
Effects of alleles at the agouti locus on minor skeletal variants in C57BL/6 house mice
-
- Journal:
- Genetical Research / Volume 47 / Issue 1 / February 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 35-41
-
- Article
-
- You have access
- Export citation
Selection for a threshold character in Drosophila I. An analysis of the phenotypic variance on the underlying scale
-
- Journal:
- Genetical Research / Volume 5 / Issue 2 / July 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 198-210
-
- Article
-
- You have access
- Export citation
GRH volume 2 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 2 / Issue 1 / February 1961
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Dislodgement and thymineless elimination of N-group plasmids
-
- Journal:
- Genetical Research / Volume 30 / Issue 1 / August 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 21-30
-
- Article
-
- You have access
- Export citation
-
Thymineless strains of Escherichia coli C600 were constructred harbouring both an R factor of the N incompatibility group (R46 or R447b) and a compatible plasmid (Plac-
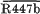 of the A-C group or the Iα plasmid R62), which contained a segment of N group DNA. Selection was made for the transferred plasmid and dislodgement phenomena were manifest either as loss of an entire plasmid or as deletions of a region of plasmid DNA. Even after the two R factors had become established as separate replicons, the N group R factor but not the other plasmid exhibited instability.
of the A-C group or the Iα plasmid R62), which contained a segment of N group DNA. Selection was made for the transferred plasmid and dislodgement phenomena were manifest either as loss of an entire plasmid or as deletions of a region of plasmid DNA. Even after the two R factors had become established as separate replicons, the N group R factor but not the other plasmid exhibited instability.Thymine starvation of strain C600 thy (R447b/R62) increased the elimination rate of the N group plasmid R447b but no elimination of R62 was observed. However, thymine starvation of strain C600 thy (R46/Plac-
) not only increased the rate of elimination of R46 but also increased the rate of loss of Plac-. There was no detectable increase in nuclease activity in unstarved R46/Plac- strains and it is concluded that dislodgement of R46 from these strains is not due to induction of the nuclease that has been proposed to be responsible for the elimination of N group plasmids during thymine starvation.Two variants of Plac-
were isolated. These did not dislodge R46 from unstarved R46/Plac- strains and were not lost during thymine starvation even though thymineless elimination of R46 occurred at normal frequency.
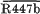 of the A-C group or the Iα plasmid R62), which contained a segment of N group DNA. Selection was made for the transferred plasmid and dislodgement phenomena were manifest either as loss of an entire plasmid or as deletions of a region of plasmid DNA. Even after the two R factors had become established as separate replicons, the N group R factor but not the other plasmid exhibited instability.
of the A-C group or the Iα plasmid R62), which contained a segment of N group DNA. Selection was made for the transferred plasmid and dislodgement phenomena were manifest either as loss of an entire plasmid or as deletions of a region of plasmid DNA. Even after the two R factors had become established as separate replicons, the N group R factor but not the other plasmid exhibited instability.Age differences between distributions of genotypes among pregnant women: evidence of fertility selection
-
- Journal:
- Genetical Research / Volume 53 / Issue 3 / June 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 207-214
-
- Article
-
- You have access
- Export citation
Biochemical investigation of the slow-growing non-perithecial (sgp) mutants of Aspergillus nidulans
-
- Journal:
- Genetical Research / Volume 17 / Issue 3 / June 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 237-244
-
- Article
-
- You have access
- Export citation
A subvital gene in Rana pipiens linked to the Burnsi locus*
-
- Journal:
- Genetical Research / Volume 20 / Issue 3 / December 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 263-268
-
- Article
-
- You have access
- Export citation
GRH volume 32 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 32 / Issue 1 / August 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Molecular Biology of RNA. UCLA Symposia on Molecular and Cellular Biology, New Series, Volume 94. By Thomas R. Cech. New York: Alan R. Liss and Sussex, England: John Wiley & Sons. 1988. 392 pages. $80.00 from Alan R. Liss. ISBN 0 8451 2693 8.
-
- Journal:
- Genetical Research / Volume 55 / Issue 1 / February 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 67-68
-
- Article
-
- You have access
- Export citation
Extensions to the model of an infinite number of selectively neutral alleles in a finite population
-
- Journal:
- Genetical Research / Volume 25 / Issue 2 / April 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 145-153
-
- Article
-
- You have access
- Export citation
-
The model, suggested by Ohta & Kimura, of an infinite number of selectively neutral alleles, in which alleles can only mutate to neighbouring alleles, has been extended to include a migratory pattern. The stepping-stone and island models of migration have been considered. In the latter case, it has been found that as the number of colonies becomes large, the expected number of alleles, ne, becomes approximately equal to
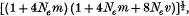
where m is the migration rate, v is the mutation rate and Ne is the effective size of each population. This solution tends to that of Ohta & Kimura as m → 0 but one can see that for any appreciable value of Nem, a large increase in ne is obtained. In order to check the validity of iterative results, models with a finite number of alleles have been considered, and their solutions have been found to converge quickly to those of the infinite case. The results exemplify the great power of migratory structure and neutral alleles to maintain a large amount of heterozygosity. Double step mutation and the finite time solution have also been considered.
Competitive mating in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 40 / Issue 2 / October 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 201-205
-
- Article
-
- You have access
- Export citation
Meiotic drive of t haplotypes: chromosome segregation in mice with tertiary trisomy
-
- Journal:
- Genetical Research / Volume 57 / Issue 1 / February 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 51-54
-
- Article
-
- You have access
- Export citation
