Refine search
Actions for selected content:
10186 results in Genetics
Chiasma variation and control in pollen mother cells and embryo-sac mother cells of rye
-
- Journal:
- Genetical Research / Volume 23 / Issue 2 / April 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 185-190
-
- Article
-
- You have access
- Export citation
Phenotypic properties of R factors of Pseudomonas aeruginosa: R factors readily transferable between Pseudomonas and the Enterobacteriaceae
-
- Journal:
- Genetical Research / Volume 23 / Issue 3 / June 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 239-250
-
- Article
-
- You have access
- Export citation
The Dictionary of Cell Biology. Edited by J. M. Lackie and J. A. T. Dow. London: Academic Press. 1989. 270 pages. –9.95 paperback. ISBN 0 12 432561 0. Also available in hardback, and in a United States edition from Academic Press Inc. San Diego CA92101.
-
- Journal:
- Genetical Research / Volume 57 / Issue 1 / February 1991
- Published online by Cambridge University Press:
- 14 April 2009, p. 95
-
- Article
-
- You have access
- Export citation
Yeast Biotechnology. Edited by D. R. Berry, I. Russell and G. G. Stewart. London: Allen and Unwin. 1987. 512 pages. £60.00. ISBN 0 04 574042 9.
-
- Journal:
- Genetical Research / Volume 52 / Issue 3 / December 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 245-246
-
- Article
-
- You have access
- Export citation
Non-additive inheritance of glucose phosphate isomerase activity in mice heterozygous at the Gpi-1s structural locus
-
- Journal:
- Genetical Research / Volume 54 / Issue 1 / August 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 27-36
-
- Article
-
- You have access
- Export citation
Books Received
-
- Journal:
- Genetical Research / Volume 45 / Issue 1 / February 1985
- Published online by Cambridge University Press:
- 14 April 2009, p. 117
-
- Article
-
- You have access
- Export citation
Quantitation of nuclear DNA in Ascaris lumbricoides: DNA constancy and chromatin diminution
-
- Journal:
- Genetical Research / Volume 27 / Issue 2 / April 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 339-348
-
- Article
-
- You have access
- Export citation
Lysosomal Storage Diseases. Biochemical and Clinical Aspects. By Richard W. E. Watts and Dorothy A. Gibbs. London: Taylor & Francis, 1968, 284 pages, £35, ISBN 0 85066 326 1.
-
- Journal:
- Genetical Research / Volume 48 / Issue 3 / December 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 205-206
-
- Article
-
- You have access
- Export citation
GRH volume 58 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 58 / Issue 1 / August 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b3
-
- Article
-
- You have access
- Export citation
GRH volume 31 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 31 / Issue 3 / June 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Localization of the properdin factor complement locus Pfc to band A3 on the mouse X chromosome
-
- Journal:
- Genetical Research / Volume 56 / Issue 2-3 / October 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 153-155
-
- Article
-
- You have access
- Export citation
Suppressors of ‘blue’ mutations in yeast
-
- Journal:
- Genetical Research / Volume 23 / Issue 1 / February 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 37-45
-
- Article
-
- You have access
- Export citation
Mutagen specificity among reversions of ultraviolet-induced adenine-1 mutants of Schizosaccharomyces pombe
-
- Journal:
- Genetical Research / Volume 6 / Issue 3 / November 1965
- Published online by Cambridge University Press:
- 14 April 2009, pp. 433-441
-
- Article
-
- You have access
- Export citation
A plasmodial colour mutation in the Myxomycete Physarum polycephalum
-
- Journal:
- Genetical Research / Volume 30 / Issue 3 / December 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 301-306
-
- Article
-
- You have access
- Export citation
Antibodies: A Laboratory Manual. Edited by Ed Harlow and David Lane. Cold Spring Harbor: Cold Spring Harbor Laboratory. New York. 1988. 726 pages. Paper $50.00. ISBN 0 87969 314 2.
-
- Journal:
- Genetical Research / Volume 54 / Issue 2 / October 1989
- Published online by Cambridge University Press:
- 14 April 2009, p. 161
-
- Article
-
- You have access
- Export citation
Genetic divergence in M. Vetukhiv's experimental populations of Drosophila pseudoobscura 2. Longevity
-
- Journal:
- Genetical Research / Volume 6 / Issue 1 / February 1965
- Published online by Cambridge University Press:
- 14 April 2009, pp. 139-146
-
- Article
-
- You have access
- Export citation
The effect of population subdivision on two loci without selection
-
- Journal:
- Genetical Research / Volume 24 / Issue 2 / October 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 151-162
-
- Article
-
- You have access
- Export citation
-
This paper is devoted to the study of the effects of population subdivision on the evolution of two linked loci. Two simple deterministic models of population subdivision without selection are investigated. One is a finite linear ‘stepping stone’ model and the other is a finite linear stepping stone chain of populations stretching between two large populations of constant genetic constitution. At equilibrium in the first model the gene frequencies in each population are equal and there is linkage equilibrium in each population. The rate of decay to zero of the linkage disequilibrium functions is the larger of (1 – c) and
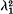 , where λ1 is the rate of convergence of the gene frequencies to equilibrium and c is the recombination frequency. In the second model at equilibrium there will be a linear cline in gene frequencies connecting the two large constant populations. This cline will be accompanied by a ‘cline’ of linkage disequilibria. The rate of convergence to this equilibrium cline is independent of the recombination frequency, and, in fact, the gene frequencies and the linkage disequilibria converge to equilibrium at the same rate.
, where λ1 is the rate of convergence of the gene frequencies to equilibrium and c is the recombination frequency. In the second model at equilibrium there will be a linear cline in gene frequencies connecting the two large constant populations. This cline will be accompanied by a ‘cline’ of linkage disequilibria. The rate of convergence to this equilibrium cline is independent of the recombination frequency, and, in fact, the gene frequencies and the linkage disequilibria converge to equilibrium at the same rate.
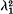 , where λ
, where λGenetic differences between populations of Drosophila melanogaster for a quantitative trait: II. Wild and laboratory populations
-
- Journal:
- Genetical Research / Volume 22 / Issue 1 / August 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 69-78
-
- Article
-
- You have access
- Export citation
The orientation of transfer of the plasmid RP4
-
- Journal:
- Genetical Research / Volume 39 / Issue 1 / February 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 99-103
-
- Article
-
- You have access
- Export citation
Introduction to Molecular Embryology Second, totally revised and enlarged edition. By J. Brachet and H. Alexandre. Berlin, Heidelberg, New York, Tokyo: Springer-Verlag, 1986. 229 pages. Soft cover DM 39, ISBN 3 540 16968 7.
-
- Journal:
- Genetical Research / Volume 50 / Issue 3 / December 1987
- Published online by Cambridge University Press:
- 14 April 2009, p. 255
-
- Article
-
- You have access
- Export citation
