Refine search
Actions for selected content:
10186 results in Genetics
Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 44 / Issue 3 / December 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 365-366
-
- Article
-
- You have access
- Export citation
Lysogenization by phage P22 carrying amber mutations in Genes 12 and 18
-
- Journal:
- Genetical Research / Volume 27 / Issue 2 / April 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 335-337
-
- Article
-
- You have access
- Export citation
GRH volume 64 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 64 / Issue 2 / October 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
The time course of recombinant production in Streptomyces coelicolor
-
- Journal:
- Genetical Research / Volume 18 / Issue 2 / October 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 133-145
-
- Article
-
- You have access
- Export citation
The general importance of disruptive selection (A comment on Waddington and Robertson's recent paper)
-
- Journal:
- Genetical Research / Volume 9 / Issue 1 / February 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 119-120
-
- Article
-
- You have access
- Export citation
The Power of Bacterial Genetics: A Literature-Based Course. By Jonathan Beckwith and Thomas J. Silhavy. Cold Spring Harbor Press1992. 828 pages. Price: Cloth $85 (ISBN 0 87969 411 4). Paper $59 (ISBN 0 87969 379 7).
-
- Journal:
- Genetical Research / Volume 62 / Issue 3 / December 1993
- Published online by Cambridge University Press:
- 14 April 2009, p. 243
-
- Article
-
- You have access
- Export citation
The genetic control of antibody production. A study of isoimmune antibodies in cattle twins*
-
- Journal:
- Genetical Research / Volume 11 / Issue 3 / June 1968
- Published online by Cambridge University Press:
- 14 April 2009, pp. 271-288
-
- Article
-
- You have access
- Export citation
Genes and Development, volume 1, no. 1, 031987. New Journal, published monthly by Cold Spring Harbor Laboratory in association with The Genetical Society of Great Britain. First year's subscription $65 plus postage for individuals, $195 plus postage for Institutions from Cold Spring Harbor Fulfillment Dept. P.O. Box 100, Cold Spring Harbor, N.Y. 11724.
-
- Journal:
- Genetical Research / Volume 49 / Issue 3 / June 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 264-265
-
- Article
-
- You have access
- Export citation
The synergistic effect of X-rays and deficiencies in DNA repair in P-M hybrid dysgenesis in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 58 / Issue 1 / August 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 15-26
-
- Article
-
- You have access
- Export citation
Correlation between genetic distances based on single loci and on skeletal morphology in inbred mice
-
- Journal:
- Genetical Research / Volume 53 / Issue 1 / February 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 45-55
-
- Article
-
- You have access
- Export citation
Plant Cell and Cell Interactions. Edited by I. Sussex, A. H. Ellingboe, M. Crouch and R. L. Malberg. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory. 1986. 151 pages. $27 paper. ISBN 0 87969 189 1.
-
- Journal:
- Genetical Research / Volume 48 / Issue 1 / August 1986
- Published online by Cambridge University Press:
- 14 April 2009, p. 59
-
- Article
-
- You have access
- Export citation
Chromosome instability related to gene suppression in Aspergillus nidulans
-
- Journal:
- Genetical Research / Volume 10 / Issue 2 / October 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 173-183
-
- Article
-
- You have access
- Export citation
The intrachromosomal mapping of a glucose phosphate isomerase structural gene, using allelic variation among stocks of Chinese Spring wheat
-
- Journal:
- Genetical Research / Volume 41 / Issue 3 / June 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 221-226
-
- Article
-
- You have access
- Export citation
On the gene conversion model as a mechanism for maintenance of homogeneity in systems with multiple genomes
-
- Journal:
- Genetical Research / Volume 30 / Issue 1 / August 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 89-91
-
- Article
-
- You have access
- Export citation
An experiment on recombination load in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 25 / Issue 3 / June 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 267-273
-
- Article
-
- You have access
- Export citation
GRH volume 57 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 57 / Issue 1 / February 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Sheep and Man. By M. L. Ryder. Duckworth, The Old Piano Factory, 43 Gloucester Crescent, London NW1, 1983. 846 pp. Price £55.00. ISBN 0 7156 16552
-
- Journal:
- Genetical Research / Volume 45 / Issue 1 / February 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 113-114
-
- Article
-
- You have access
- Export citation
Genetic analysis of the right (3′) end of the rosy locus in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 47 / Issue 2 / April 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 109-116
-
- Article
-
- You have access
- Export citation
Molecular analysis of opaque-2 alleles from Zea mays L. reveals the nature of mutational events and the presence of a hypervariable region in the 5′ part of the gene
-
- Journal:
- Genetical Research / Volume 65 / Issue 1 / February 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 11-19
-
- Article
-
- You have access
- Export citation
A genetic study of primary and secondary reversions of some tryptophanA auxotrophs of Salmonella typhimurium
-
- Journal:
- Genetical Research / Volume 9 / Issue 3 / June 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 269-282
-
- Article
-
- You have access
- Export citation
-
1. The linkage order of four tryA mutants of S. typhimurium, and cysB-12, is:
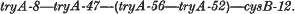
Attempts to plot the position of tryA-50 were unsuccessful.
2. Some of the reversions of tryA-8, tryA-47, tryA-56 and tryA-50 were analysed genetically; tryA-52 does not revert. All four auxotrophs gave reversions that were phenotypically and genetically indistinguishable from that expected by back-mutation of the original mutant site.
3. Both tryA-8 and tryA-50 produced reversions that grew as wild-type but were due to unlinked suppressor mutations. Some of these were super-suppressors in that they suppressed both tryA-8 and tryB-4; others suppressed many site mutants in the tryA gene but did not suppress tryB-4.
4. All the slow-growing reversions of tryA-8, tryA-50 and tryA-56, and a minority of the semi-fast reversions of tryA-8, were due to unlinked suppressors.
5. All the slow-growing reversions of tryA-47, the semi-fast reversions of tryA-56 and the majority of the semi-fast reversions of tryA-8 were due to genetic changes that were inseparable, in very extensive experiments, from their original mutant site.
6. Slow-growing reversions of tryA-47 produced faster growing mutants. Some of these were due to mutation in unlinked modifying genes and in others the genetic change was within the tryA gene. Nine of the latter had the genetic change just to the left of the 47S site; in one the change was inseparable from the 47S site. None had this further change to the right of the 47S site. These further changes, in the absence of the 47S site, gave prototrophic phenotypes; they are inter-site suppressors.