Refine search
Actions for selected content:
6997 results in Mathematical modeling and methods
Transformation of Narrowband Wavetrains of Surface GravityWaves Passing over a Bottom Step
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 5 / 2014
- Published online by Cambridge University Press:
- 17 July 2014, pp. 73-82
- Print publication:
- 2014
-
- Article
- Export citation
Importance of VIIIa Inactivation in a Mathematical Model forthe Formation, Growth, and Lysis of Clots
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 6 / 2014
- Published online by Cambridge University Press:
- 31 July 2014, pp. 17-33
- Print publication:
- 2014
-
- Article
- Export citation
Efficient greedy algorithms for high-dimensional parameter spaces with applications to empirical interpolation and reduced basis methods∗
-
- Journal:
- ESAIM: Mathematical Modelling and Numerical Analysis / Volume 48 / Issue 1 / January 2014
- Published online by Cambridge University Press:
- 10 January 2014, pp. 259-283
- Print publication:
- January 2014
-
- Article
- Export citation
Systems Biology and Systems Pharmacology ofThrombosis
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 6 / 2014
- Published online by Cambridge University Press:
- 31 July 2014, pp. 4-16
- Print publication:
- 2014
-
- Article
- Export citation
Constant Versus Periodic Fishing: Age Structured OptimalControl Approach
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 4 / 2014
- Published online by Cambridge University Press:
- 20 June 2014, pp. 20-37
- Print publication:
- 2014
-
- Article
- Export citation
Preface
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 2 / 2014
- Published online by Cambridge University Press:
- 24 April 2014, pp. 1-3
- Print publication:
- 2014
-
- Article
- Export citation
CMV Matrices with Super Exponentially Decaying VerblunskyCoefficients
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 5 / 2014
- Published online by Cambridge University Press:
- 17 July 2014, pp. 282-294
- Print publication:
- 2014
-
- Article
- Export citation
Coupling Vector-host Dynamics with Weather Geography andMitigation Measures to Model Rift Valley Fever in Africa
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 2 / 2014
- Published online by Cambridge University Press:
- 24 April 2014, pp. 161-177
- Print publication:
- 2014
-
- Article
- Export citation
Linear Size-structured Population Models with Spacial Diffusion and Optimal Harvesting Problems
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 4 / 2014
- Published online by Cambridge University Press:
- 20 June 2014, pp. 122-130
- Print publication:
- 2014
-
- Article
- Export citation
Preface
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 4 / 2014
- Published online by Cambridge University Press:
- 20 June 2014, pp. 1-5
- Print publication:
- 2014
-
- Article
- Export citation
Preface
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 1 / 2014
- Published online by Cambridge University Press:
- 07 February 2014, pp. 1-3
- Print publication:
- 2014
-
- Article
- Export citation
Diffusion and Deterministic Systems
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 1 / 2014
- Published online by Cambridge University Press:
- 07 February 2014, pp. 139-150
- Print publication:
- 2014
-
- Article
- Export citation
Tridiagonal Substitution Hamiltonians
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 5 / 2014
- Published online by Cambridge University Press:
- 17 July 2014, pp. 204-238
- Print publication:
- 2014
-
- Article
- Export citation
Variational approximation of a functional of Mumford–Shah typein codimension higher than one
-
- Journal:
- ESAIM: Control, Optimisation and Calculus of Variations / Volume 20 / Issue 1 / January 2014
- Published online by Cambridge University Press:
- 27 January 2014, pp. 190-221
- Print publication:
- January 2014
-
- Article
- Export citation
Patient Specific Haemodynamic Modeling after OcclusionTreatment in Leg
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 6 / 2014
- Published online by Cambridge University Press:
- 31 July 2014, pp. 85-97
- Print publication:
- 2014
-
- Article
- Export citation
Inverse Scattering Problem with UnderdeterminedData
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 5 / 2014
- Published online by Cambridge University Press:
- 17 July 2014, pp. 244-253
- Print publication:
- 2014
-
- Article
- Export citation
Exact null internal controllability for the heat equation on unbounded convex domains∗
-
- Journal:
- ESAIM: Control, Optimisation and Calculus of Variations / Volume 20 / Issue 1 / January 2014
- Published online by Cambridge University Press:
- 27 January 2014, pp. 222-235
- Print publication:
- January 2014
-
- Article
- Export citation
-
The liner parabolic equation \hbox{$\frac{\pp y}{\pp t}-\frac12\,\D y+F\cdot\na y={\vec{1}}_{\calo_0}u$}
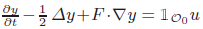 with Neumann boundary condition on a convex open domain 𝒪 ⊂ ℝdwith smooth boundary is exactly null controllable on each finite interval if 𝒪0is an open subset of 𝒪which contains a suitable neighbourhood of the recession cone of \hbox{$\ov\calo$}
with Neumann boundary condition on a convex open domain 𝒪 ⊂ ℝdwith smooth boundary is exactly null controllable on each finite interval if 𝒪0is an open subset of 𝒪which contains a suitable neighbourhood of the recession cone of \hbox{$\ov\calo$}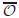 . Here, F : ℝd → ℝd is a bounded, C1-continuous function, and F = ∇g, where g is convex and coercive.
. Here, F : ℝd → ℝd is a bounded, C1-continuous function, and F = ∇g, where g is convex and coercive.
Modelling Biological Evolution: Introduction to the SpecialIssue
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 3 / 2014
- Published online by Cambridge University Press:
- 28 May 2014, pp. 1-4
- Print publication:
- 2014
-
- Article
-
- You have access
- Export citation
Optimal Control of Leukemic Cell PopulationDynamics
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 1 / 2014
- Published online by Cambridge University Press:
- 07 February 2014, pp. 4-26
- Print publication:
- 2014
-
- Article
- Export citation
Optimal Vaccination, Treatment, and Preventive Campaigns inRegard to the SIR Epidemic Model
-
- Journal:
- Mathematical Modelling of Natural Phenomena / Volume 9 / Issue 4 / 2014
- Published online by Cambridge University Press:
- 20 June 2014, pp. 105-121
- Print publication:
- 2014
-
- Article
- Export citation
