



Introduction
The classification of parasitic mites has historically relied heavily on conserved morphological characters, leading to the recognition of genera with remarkably broad geographic distributions and host ranges. Within Mesostigmata, this pattern is exemplified by genera such as Androlaelaps Berlese, 1903, Laelaps Koch, 1836 (Laelapidae) and Ornithonyssus Sambon, 1928 (Macronyssidae). These mite genera include species traditionally considered globally distributed and associated with a wide array of hosts (Tipton, Reference Tipton1960; Radovsky, Reference Radovsky and Kim1985, Reference Radovsky and Houck1994).
This study focuses on Laelaps, a genus reported from all continents, except Antarctica, parasitizing primarily rodents of the families Cricetidae and Muridae (Tipton, Reference Tipton1960; Domrow, Reference Domrow1964; Matthee et al., Reference Matthee, Engelbrecht and Matthee2018; Savchenko and Lareschi, Reference Savchenko and Lareschi2022; Yang et al., Reference Yang, Yang and Dong2022; Lareschi, Reference Lareschi, Claps, S and Morrone2023). The genus was erected by Koch (1836) describing 2 species, Laelaps agilis and Laelaps hilaris, collected from murid and cricetid rodents, respectively, from Germany. Subsequently, Laelaps underwent multiple taxonomic revisions based on morphology that progressively refined the boundaries of the genus (Zachvatkin, Reference Zachvatkin1948; Zumpt, Reference Zumpt1950; Zumpt and Patterson, Reference Zumpt and Patterson1951; Bregetova, Reference Bregetova1956; Evans and Till, Reference Evans and Till1966). However, these boundaries are still not entirely clear. Tipton (Reference Tipton1960) and Furman and Tipton (Reference Furman and Tipton1961) restricted Laelaps sensu stricto, excluding related laelapid genera (e.g. Echinolaelaps Ewing, 1929), whereas other authors treated Echinolaelaps and Laelaps sensu stricto as subgenera within a broader Laelaps sensu lato by incorporating additional morphological traits (see Strandtmann and Mitchell, Reference Strandtmann and Mitchell1963; Allred, Reference Allred1969; Furman, Reference Furman1971). This taxonomic framework has a certain degree of subjectivity, and it may have contributed to the long-standing perception of Laelaps as a single, widely distributed genus.
The reports of Laelaps in South America began with the descriptions of species, such as Laelaps differens, Laelaps manguinhosi and Laelaps paulistanensis, all associated with cricetid rodents from Brazil (Fonseca, Reference Fonseca1936a). Currently, there is an uncertain number of Laelaps species recognized worldwide. Some species parasitize Old World murid rodents, whereas other are associated with South American cricetids (Fonseca, Reference Fonseca1936a, Reference Fonseca1939; Tipton, Reference Tipton1960; Furman, Reference Furman1972a, b; Gettinger, Reference Gettinger1992; Savchenko and Lareschi, Reference Savchenko and Lareschi2022). This pattern has generally been interpreted as evidence of a single, broadly defined genus with a global distribution. However, this interpretation becomes problematic when viewed in an evolutionary context. Cricetidae and Muridae are sister families – if fossil taxa are excluded – that diverged around 25 million years ago and have since followed markedly different evolutionary trajectories, biogeographic histories and ecological associations (e.g. Steppan and Schenk, Reference Steppan and Schenk2017; López-antoñanzas et al., Reference López-antoñanzas, Simões, Condamine, Dirnberger and Peláez-campomanes2024). Some species of murids, such as Rattus rattus (Linnaeus, 1758) and Rattus norvegicus (Berkenhout, 1769), have achieved global distributions largely through human-mediated dispersal and are strongly associated with anthropized environments (Steppan et al., Reference Steppan, Adkins, Spinks and Hale2005; Aghová et al., Reference Aghová, Kimura, Bryja, Dobigny, Granjon and Kergoat2018; Byers et al., Reference Byers, Lee, Patrick and Himsworth2019). In contrast, cricetids underwent a rapid radiation in South America, remaining predominantly linked to natural or less anthropized habitats (Patton et al., Reference Patton, Pardiñas and D’elía2015; Pardiñas et al., Reference Pardiñas, Myers, León-paniagua, Ordóñez Garza, Cook, Kryštufek, Haslauer, Bradley, Shenbrot, Patton, Wilson, Lacher and Mittermeier2017; Ronez et al., Reference Ronez, Carrillo-briceño, Hadler, Sánchez-villagra and Pardiñas2023).
From an evolutionary perspective, the deep phylogenetic divergence of Cricetidae and Muridae raises doubts about the plausibility of long-term persistence of the same parasite mite lineages across both rodent families. Host–parasite co-evolutionary theory predicts that parasites with relatively strong host associations may tend to track host phylogeny over long evolutionary timescales, making true cosmopolitanism across deeply divergent host lineages unlikely (Hugot, Reference Hugot, Morand, Krasnov and Poulin2007). Under this framework, the apparent cosmopolitan distribution of Laelaps may reflect historical reliance on morphology-based taxonomy rather than genuine evolutionary continuity.
Despite extensive documentation of host associations in Laelaps, phylogenetic relationships within the genus remain poorly explored, a gap particularly relevant given the role of its hosts as potential vectors of zoonotic pathogens (Bowen et al., Reference Bowen, Peters and Nichol1997; Babolin et al., Reference Babolin, Almeida-silva, Potenza, Fava, Castro, Harakava, Okuda, Rebouças and Campos2016). Previous molecular studies have suggested that host association may be influential in shaping Laelaps diversification. For example, host-specialist species such as Laelaps giganteus Berlese, 1918, show phylogenetic structure closely linked to murid hosts, whereas more generalist species such as Laelaps muricola Trägårdh, 1910 exhibit weaker host-associated patterns (Engelbrecht et al., Reference Engelbrecht, Matthee, Ueckermann and Matthee2014, Reference Engelbrecht, Matthee, Du Toit and Matthee2016; Matthee et al., Reference Matthee, Engelbrecht and Matthee2018). Similarly, Savchenko and Lareschi (Reference Savchenko and Lareschi2022) reported a dichotomy between Laelaps associated with South American cricetids and those parasitizing South African murids, although their analysis was based on a single molecular marker (18S-ITS1-5.8S-ITS2), leaving the evaluation of the monophyly of the genus unresolved.
The primary aim of this study is to evaluate the monophyly of the genus Laelaps. To this end, we integrate new molecular data from Laelaps echidninus Berlese, 1887 associated with Rattus norvegicus (Muridae) in Argentina, a recently introduced species in contact with native cricetids, together with sequences from laelapid mites parasitizing native cricetids from nearby localities and congeners from South Africa available in GenBank. This study design allows us to assess whether Laelaps is monophyletic and whether host association structures laelapid phylogeny.
Materials and methods
Studied material
Mites analysed in this study were collected by the authors and collaborators as part of broader research on rodents and parasites conducted in various localities across Argentina in previous years. Specimens were preserved in 96% ethanol and stored frozen at −20 °C. A total of 23 mites from 11 localities from Argentina (see Figure S1 in supplementary material) were selected from 15 rodents specimens representing the following species: Cricetidae: Calomys callosus (Rengger, 1830), Calomys sorella (Thomas, 1900), Holochilus brasiliensis (Desmarest, 1819), Holochilus chacarius Thomas, 1906, Nectomys squamipes (Brants, 1827), Oligoryzomys nigripes (von Olfers, 1818), Oligoryzomys flavescens (Waterhouse, 1837), Oligoryzomys fornesi (Massoia, 1973), Scapteromys aquaticus Thomas, 1920; Muridae: Rattus norvegicus (Berkenhout, 1769) (see details in Table 1).
DNA sequences included in phylogenetic analysis of nuclear region 18S-ITS1-5.8S-ITS2-28S and cytochrome oxidase I (COI)

Table 1 Long description
The table lists mite taxa used for phylogenetic sampling, paired with their host species, host family and tribe, country of mite origin, and accession numbers for a nuclear ribosomal region and the COI gene. Sampling is dominated by Laelaps species associated with rodents, especially Cricetidae and Muridae, with most records originating from Argentina and several from South Africa, Italy, Israel, and Peru. Dermanyssus gallinae on domestic chicken is included as an outgroup, with one Italian record providing a nuclear accession and one Israeli record providing a COI accession, indicating missing data across markers for that species. Several Laelaps species have multiple entries reflecting different hosts or multiple sequences per marker, such as Laelaps mazzai across several Calomys hosts in Argentina plus one Peruvian record. Many COI accessions are newly generated in the study, while some nuclear and COI sequences are drawn from prior publications. Missing values are explicitly indicated for some marker-by-record combinations, so comparisons across taxa may rely on different gene coverage.
Sequences obtained in this study are indicated with asterisks (*), no data (nd).
Following molecular procedures, mite exoskeletons were recovered from Chelex®100 solution, were cleared in lactophenol and mounted in Hoyer’s medium for their study by light microscopy. Mite identification was conducted based on original descriptions (Fonseca, Reference Fonseca1936b, Reference Fonseca1958; Savchenko and Lareschi, Reference Savchenko and Lareschi2019, Reference Savchenko and Lareschi2022).
Molecular procedures
All mites were processed for DNA isolation. This procedure was applied individually to each mite using Chelex®100 following a non-destructive protocol (Savchenko and Lareschi, Reference Savchenko and Lareschi2019). Two Polymerase Chain Reaction (PCR) were performed: one targeting the nuclear 18S-ITS1-5.8S-ITS2-28S region and another amplifying the mitochondrial cytochrome oxidase I (COI) gene. For the 18S-ITS1-5.8S-ITS2-28S region, PCR conditions consisted of an initial denaturation at 95 °C for 10 min, followed by 35 cycles of denaturation at 95 °C for 20 s, annealing at 51 °C for 30 s, extension at 72 °C for 90 s and a final extension at 72 °C for 10 min. The primers used were RhITS-F (5′-AGAGGAAGTAAAAGTCGTAACAAG-3′) and RhITS-R (5′-ATATGCTTAAATTCAGGGGG-3′) (De Rojas et al., Reference De Rojas, Mora, Ubeda, Cutillas, Navajas and Guevara2002).
PCR amplification of COI followed a similar protocol, with an annealing temperature of 50 °C. The primers used were Kmt6 (5′-GGAGGATTTGGAAATTGATTAGTTCC-3′) and HC02198 (5′-TAAACTTCAGGGTGACCAAAAAATCA-3′) (Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994; Zhu et al., Reference Zhu, Hastriter, Whiting and Dittmar2015). PCR products were sequenced by Macrogen (Seoul, South Korea).
Data analysis
Forward and reverse sequences were assembled, edited and aligned using CodonCode Aligner (CodonCode Corporation). The sequences generated for the 18S-ITS1-5.8S-ITS2-28S nuclear region and the COI gene were compared with sequences of laelapine mites available in the GenBank database, including 32 sequences of the nuclear region and 17 sequences of COI (see details in Table 1). Sequences of Androlaelaps fahrenholzi (Berlese, 1911), Gigantolaelaps wolffsohni (Oudemans, 1910), Mysolaelaps microspinosus Fonseca, 1936 and Mysolaelaps parvispinosus Fonseca, 1936 retrieved from GenBank were included to test relationships among laelapid genera. Sequences of Dermanyssus gallinae (De Geer, 1778) (Mesostigmata: Dermanyssidae) were used as outgroups.
Two separate alignments were constructed: one including 41 sequences of the 18S-ITS1-5.8S-ITS2-28S region and another including 41 sequences of the COI gene. These alignments were subsequently concatenated into a single dataset of approximately 1300 bp. Given the distinct evolutionary patterns of nuclear and mitochondrial markers, phylogenetic analyses were conducted using a partitioned dataset. Substitution models were selected independently for each dataset using jModelTest 2 (Darriba et al., Reference Darriba, Taboada, Doallo and Posada2012) under the Akaike information criterion (AIC) and the corrected AIC. The Hasegawa-Kishino-Yano (HKY) + I + G was selected for the 18S-ITS1-5.8S-ITS2-28S region, whereas General Time Reversible (GTR) + I + G was selected for COI.
Phylogenetic relationships were inferred from the concatenated dataset using both maximum likelihood (ML) and Bayesian inference (BI). ML analyses were performed in IQ-Tree (Minh et al., Reference Minh, Schmidt, Chernomor, Schrempf, Woodhams, Von Haeseler and Lanfear2020) with 2000 bootstrap replicates, while BI analyses were conducted in MrBayes 3.2.7 (Ronquist et al., Reference Ronquist, Teslenko, Van der Mark, Ayres, Darling, Höhna, Larget, Liu, Suchard and Huelsenbeck2012) for 10 million generations, with the first 25% of trees discarded as burn-in. Phylogenetic trees were visualized using Figtree v1.4.4. The topology of the ML tree was selected and displayed the ML bootstrap values and Bayesian posterior probabilities.
Pairwise genetic distances among groups of mites, obtained from the phylogenetic tree, were calculated in MEGA11 using the alignments separately, and those results were expressed as percentages.
Results
Of the 23 analysed mites, the following taxa were identified: 1 A. fahrenholzi from C. callosus, 3 G. wolffsohni (1 from O. flavescens and 2 from O. nigripes), 2 L. echidninus from R. norvegicus, 2 Laelaps galliarii Savchenko and Lareschi, 2022 from O. nigripes, 3 L. manguinhosi Fonseca, 1936 (2 from H. brasiliensis and 1 from H. chacarius), 1 Laelaps mazzai (Fonseca, 1939) from C. sorella, 1 Laelaps odysseos Savchenko and Lareschi, 2022 from N. squamipes, 2 L. paulistanensis Fonseca, 1936 from O. nigripes, 3 Laelaps scapteromyos Savchenko and Lareschi, 2022 from S. aquaticus, 1 Laelaps schatzi Lareschi and Savchenko, 2021 from O. fornesi, 2 M. microspinosus and 2 M. parvispinosus, all from O. flavescens. From these specimens, 8 sequences of the 18S-ITS1-5.8S-ITS2-28S nuclear region and 23 COI sequences were generated (Table 1).
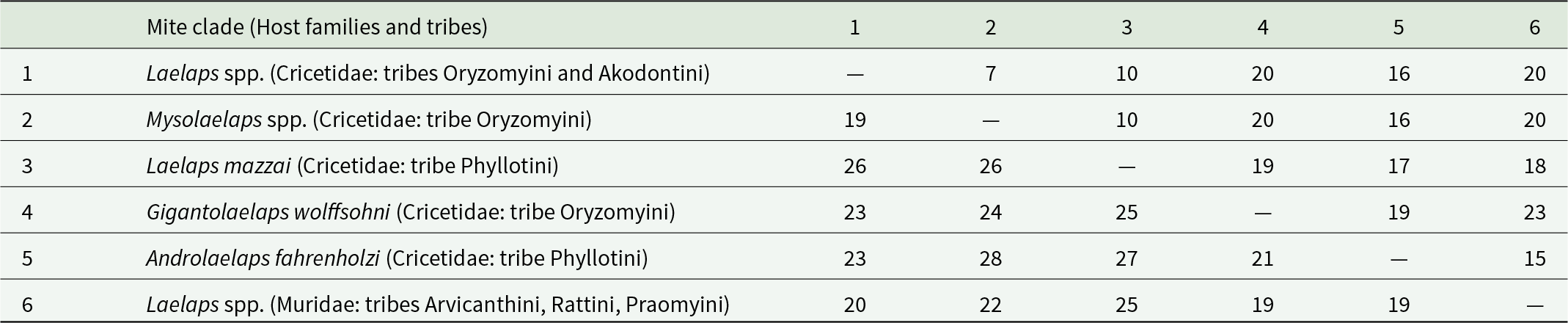
Phylogenetic analyses of sequences generated in this study and those retrieved from GenBank (Table 1) revealed that all cricetid-associated mites formed a well-supported clade, separated from the murid-associated taxa. In contrast, the 3 species of murid-associated mites, L. echidninus (South America), L. giganteus and L. muricola (both from South Africa), did not form a well-supported monophyletic group, but instead were recovered as basal lineages separated from all the cricetid-associated Laelaps. Genetic distances between murid- and cricetid-associated laelapids ranged from 15% to 23% for the 18S-ITS1-5.8S-ITS2-28S region and from 19% to 25% for COI (Table 2).
Genetic distances in percentage between clades of mites obtained from the phylogenetic tree for 18S-ITS1-5.8S-ITS2-28S and cytochrome oxidase I (COI)

Table 2 Long description
The table reports pairwise genetic distances, in percent, among six mite clades linked to different rodent host families and tribes. For each clade pair, the upper-right half gives distances for the 18S-ITS1-5.8S-ITS2-28S region, and the lower-left half gives distances for COI. In the 18S-ITS region, the smallest distance is 7 percent between clades 1 and 2, and most other values fall between 10 and 23 percent, with several pairs at 20 percent. In COI, distances are generally higher, ranging from 19 to 28 percent, with the largest distance between clades 2 and 5 at 28 percent. COI also shows 26 percent between clade 3 and clades 1 and 2, and 25 percent between clade 6 and clades 3 and 4. The lowest COI distance is 19 percent, occurring for multiple pairs including clades 1 and 2, clades 1 and 6, clades 4 and 6, and clades 5 and 6. Overall, COI indicates greater divergence among clades than the 18S-ITS region, but comparisons must use the correct half of the matrix because each marker is reported in a different triangle.
Values above the diagonal indicate genetic distances of the 18S-ITS1-5.8S-ITS2-28S region and under the diagonal for COI.
Within the Cricetidae-associated clade, 5 subclades were recovered. One subclade included A. fahrenholzi associated with Phyllotini rodents and was recovered as sister of the remaining taxa. A second subclade comprised G. wolffsohni associated with Oryzomyini rodents. A third subclade included L. mazzai, also associated exclusively with Phyllotini. A fourth subclade grouped Mysolaelaps species associated with Oryzomyini rodents. The fifth subclade included Laelaps species associated with Oryzomyini and Akodontini rodents, namely L. galliarii, L. paulistanensis, L. schatzi, L. manguinhosi, L. odysseos and L. scapteromyos, though with weaker nodal support (Figure 1). Genetic distances between Mysolaelaps species and Laelaps associated with Oryzomyini rodents were lower (18S-ITS1-5.8S-ITS2-28S: 7%, COI: 19%) than distances between L. mazzai and Laelaps species associated with Oryzomyini and Akodontini rodents (18S-ITS1-5.8S-ITS2-28S: 10%, COI: 26%) (Table 2).
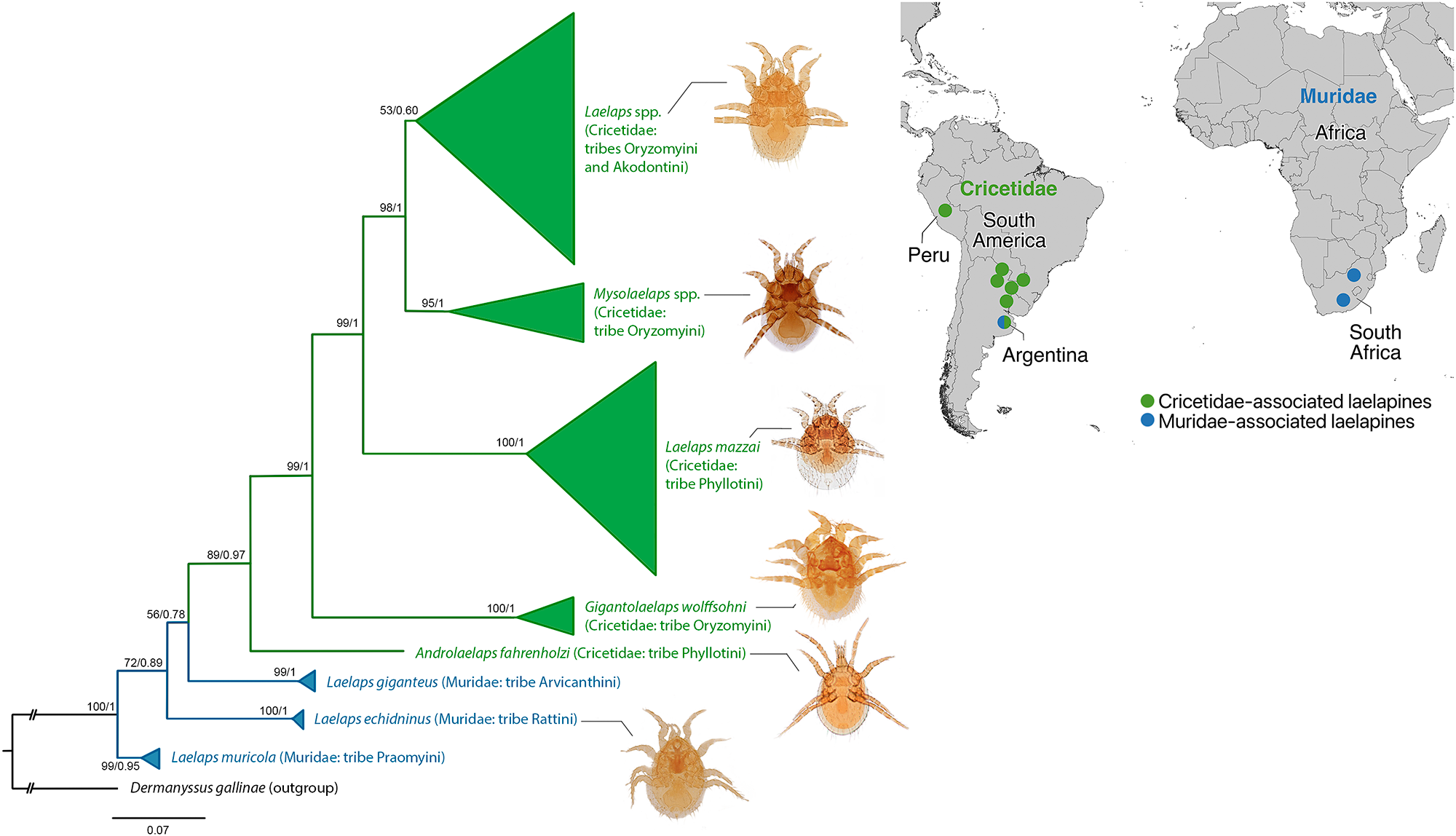
Phylogenetic tree based on concatenated nuclear (18S-ITS1-5.8S-ITS2-28S) and mitochondrial (COI) genes of laelapine mites from Muridae and Cricetidae rodents. The topology of the maximum likelihood (ML) tree was selected and displayed, on the nodes, the ML bootstrap values (left) and Bayesian posterior probabilities (BI, right). Branch colours and coloured circles on the map indicate host family associations. Coloured circles on the map represent approximate locations and are shown for reference only, not precise sampling points. Green denotes lineages associated with Cricetidae, whereas blue denotes lineages associated with Muridae. The presence of both colours in Argentina reflects the occurrence of cricetid-associated and murid-associated Laelaps lineages.

Figure 1 Long description
The diagram is a phylogenetic tree with a map indicating host associations of laelapine mites with Cricetidae and Muridae rodents. The tree is oriented vertically, with branches extending from the bottom to the top. The left side of the tree features green branches representing lineages associated with Cricetidae, while blue branches on the right represent lineages associated with Muridae. Each branch is labeled with bootstrap values and Bayesian posterior probabilities. The topmost green clade includes Laelaps species associated with Oryzomyini and Akodontini rodents. Below it, another green clade represents Mysolaelaps species associated with Oryzomyini. Further down, a green clade includes Laelaps mazzai associated with Phyllotini. The lowest green clade groups G. wolffsohni with Oryzomyini. The blue clade at the bottom includes Laelaps species associated with Muridae, such as L. giganteus and L. muricola. The map on the right shows approximate locations of these associations in South America and South Africa, with green circles for Cricetidae and blue circles for Muridae. The presence of both colors in Argentina indicates the occurrence of both cricetid-associated and murid-associated lineages.
Overall, these results indicate that Laelaps is polyphyletic. Within Cricetidae, phylogenetic structure appears linked to host tribal affiliation, with distinct lineages recovered for Phyllotini, Akodontini and Oryzomyini hosts, respectively (Figure 1), separated by substantial genetic distances (18S-ITS1-5.8S-ITS2-28S: 18–20%, COI: 20–25%) (Table 2). The murid-associated lineages, while recovered outside the cricetid clade, lacked sufficient nodal support to be interpreted as a coherent monophyletic group.
Discussion
The primary result of this study is the polyphyly of Laelaps, with cricetid-associated lineages forming a well-supported clade with a tendency for murid-associated taxa to be recovered separately. Within the cricetid-associated clade, phylogenetic structure appears linked to host tribal affiliation, with distinct lineages recovered for Phyllotini, Oryzomyini and Akodontini rodents. Notably, the entanglement of morphologically distinct genera (Androlaelaps, Gigantolaelaps and Mysolaelaps) within the cricetid-associated clade further demonstrates that current generic boundaries in Laelapidae do not reflect evolutionary history.
The murid-associated lineages, while recovered outside the cricetid clade, did not form a well-supported monophyletic group, and their phylogenetic relationships remain unresolved pending broader sampling. However, the substantial genetic distances between cricetid- and murid-associated laelapids are consistent with long-term independent diversification following the divergence of Cricetidae and Muridae around 25 Mya ago (e.g. Steppan and Schenk, Reference Steppan and Schenk2017; López-antoñanzas et al., Reference López-antoñanzas, Simões, Condamine, Dirnberger and Peláez-campomanes2024). From a co-evolutionary perspective, such a temporal depth makes a long-term persistence of the same specialized parasite lineages across both families highly improbable (Hugot, Reference Hugot, Morand, Krasnov and Poulin2007). The polyphyly of Laelaps revealed here is therefore consistent with this expectation and suggests that the traditional treatment of Laelaps as a single, widely distributed genus reflects historical reliance on morphologically conservative characters rather than genuine evolutionary continuity.
Since its description, Laelaps has been repeatedly revised, with successive authors refining its diagnostic limits while emphasizing morphological uniformity and ecological similarity across species (Zachvatkin, Reference Zachvatkin1948; Zumpt, Reference Zumpt1950; Bregetova, Reference Bregetova1956). Although Tipton (Reference Tipton1960) formalized the concept of Laelaps sensu stricto, other authors adopted a broader interpretation (Laelaps sensu lato) by retaining lineages separated by differences in chaetotaxy and shield morphology, including the treatment of Echinolaelaps as a subgenus of Laelaps (e.g. Strandtmann and Mitchell, Reference Strandtmann and Mitchell1963). The polyphyly recovered here is consistent with this long history of taxonomic instability and suggests that the morphological characters traditionally used to delimit Laelaps and related genera are evolutionarily labile and insufficient to capture phylogenetic structure. Notably, species such as L. giganteus and L. muricola were previously included within the Echinolaelaps complex together with L. echidninus (Strandtmann and Mitchell, Reference Strandtmann and Mitchell1963), yet our results show these murid-associated species occupy a very different phylogenetic position from the cricetid-associated lineages. Clarifying the molecular placement of additional species historically assigned to this complex will be essential to evaluate the potential resurrection of Echinolaelaps as a valid genus.
Within Cricetidae-associated mites, the paraphyly of Laelaps with respect to Mysolaelaps and Gigantolaelaps is also noteworthy. Although these genera are morphologically distinguishable based on major characters, particularly the shape and chaetotaxia of the genitoventral shield (see details in Fonseca, Reference Fonseca1939, Reference Fonseca1958), such differences appear insufficient to ensure taxonomic stability in the absence of molecular evidence. This situation illustrates a recurrent challenge in laelapid systematics: determining the taxonomic level at which morphological variation should be interpreted, whether at the species, genus or higher ranks (e.g. Furman, Reference Furman1972b). Historically, these decisions have involved a considerable degree of subjectivity, contributing to the persistence of broadly defined and potentially artificial genera within Laelapidae (Dowling and OConnor, Reference Dowling and OConnor2010). In Laelaps, this subjectivity is particularly evident in the interpretation of fine differences in idiosomal chaetotaxy and shield morphology, characters that have traditionally been considered sufficient to delimit genera despite their apparent evolutionary lability (Furman, Reference Furman1971).
The case of L. echidninus associated with R. norvegicus in South America provides additional insights into parasite dynamics during host introductions. Despite the relatively recent introduction of R. norvegicus into the New World via human activities (Steppan et al., Reference Steppan, Adkins, Spinks and Hale2005), L. echidninus appears to have been retained by its host and shows no evidence of host switching to native sympatric cricetids. This pattern contrasts with expectations of the enemy release hypothesis, which predicts reduced parasite loads in introduced hosts (Torchin et al., Reference Torchin, Lafferty, Dobson, McKenzie and Kuris2003). Instead, our results are consistent with the view that highly host-specific parasites are more likely to persist during host range expansion, whereas generalist parasites are more prone to loss (Poulin, Reference Poulin1998; Schatz and Park, Reference Schatz and Park2023). Similar patterns have been reported across diverse parasite groups, including arthropods, bacteria, protozoa and viruses, and have been described as uncommon but plausible outcomes of host introductions (Schatz and Park, Reference Schatz and Park2023).
The absence of spillover to native rodents and the lack of spillback from cricetids to Rattus further emphasize the strong host specificity of L. echidninus, with only occasional accidental associations reported in the literature (Lareschi, Reference Lareschi1996). In such cases, the presence of L. echidninus on the native rodent S. aquaticus was interpreted as the use of an alternative host in environments where this rodent is in sympatry with Rattus spp. in ecological disturbed environment (Lareschi, Reference Lareschi1996). Indeed, S. aquaticus has been identified as a suitable alternative host for multiple laelapid mites in riparian environments of the Río de la Plata basin (Lareschi, Reference Lareschi1996).
Within the cricetid-associated clade, A. fahrenholzi emerged as a sister lineage to the remaining taxa. This mite represents a complex species and has been suggested as a transitional stage between free-living predatory mites and obligate parasites (Radovsky, Reference Radovsky and Houck1994; Dowling, Reference Dowling, Morand, Krasnov and Poulin2006; Dowling and OConnor, Reference Dowling and OConnor2010). As occurs with Laelaps, Androlaelaps is a genus that requires taxonomic revision. Additional sampling across host species and geographic regions will be necessary to clarify its evolutionary position.
The remaining mites formed a complex assemblage including Gigantolaelaps, Mysolaelaps and 2 lineages currently assigned to Laelaps, a pattern that directly illustrates the inadequacy of current generic boundaries. Within Laelaps, one lineage represented by L. mazzai was recovered exclusively in association with Calomys spp. (Phyllotini), whereas the other comprised species predominantly associated with Oryzomyini rodents (L. galliarii, L. manguinhosi, L. odysseos, L. paulistanensis and L. schatzi), with the exception of L. scapteromyos associated with S. aquaticus (Akodontini). The fact that Gigantolaelaps and Mysolaelaps, morphologically distinct genera (Fonseca, Reference Fonseca1939, Reference Fonseca1958; Furman, Reference Furman1972b), are recovered as more closely related to some Laelaps lineages than those Laelaps lineages are to each other further underscores that morphological characters alone are insufficient to delimit natural groups within Laelapidae. Some of these lineages do exhibit marked morphological differences (Savchenko and Lareschi, Reference Savchenko and Lareschi2022), but others, such as L. mazzai, remain difficult to distinguish using traditional characters, likely explaining their long-standing inclusion within a broadly defined Laelaps.
The phylogenetic distinctiveness of L. mazzai is particularly noteworthy given its taxonomic history. This species was previously proposed as belonging to a separate genus, Schistolaelaps Fonseca, 1959, based on morphological differences including a hypertrichous idiosoma, but was later re-included within Laelaps when those differences were considered insufficient at the generic level (Furman, Reference Furman1971, Reference Furman1972b). Interestingly, Dowling and OConnor (Reference Dowling and OConnor2010) had already flagged this taxon as warranting separate generic recognition based on their broader phylogenetic analysis of Dermanyssoidea. Our molecular results, with strong nodal support for the L. mazzai lineage, corroborate this interpretation and lend support to the revalidation of Schistolaelaps for species of Laelaps associated with Phyllotini rodents of the genus Calomys.
The polyphyly of Laelaps recovered here underscores the need for a comprehensive taxonomic revision of Laelapidae integrating molecular, morphological and ecological data. Based on the evidence available, at least 3 distinct evolutionary entities can be recognized within what is currently treated as Laelaps: lineages associated with Muridae (though their monophyly remains unresolved), lineages associated with Cricetidae–Oryzomyini and Akodontini, and lineages associated with Phyllotini (Calomys), the latter likely corresponding to the revalidated genus Schistolaelaps. More broadly, these findings challenge the validity of other parasite taxa-associated murid and cricetid rodents that have traditionally been regarded as cosmopolitan based solely on morphological similarity, such as A. fahrenholzi and Ornithonyssus bacoti (Hirst, 1913). Both remain poorly explored from a phylogenetic perspective and, in light of our results, clearly warrant further integrative study (Till, Reference Till1963; Radovsky, Reference Radovsky and Houck1994; Dowling and OConnor, Reference Dowling and OConnor2010).
From an epidemiological perspective, the persistence of L. echidninus on introduced Rattus populations is noteworthy. This mite has been associated with bacteria of the genera Anaplasma and Rickettsia in Asia and Europe (Mit’ková et al., Reference Mit’ková, Berthová, Kalúz, Kazimírová, Burdová and Kocianová2015; Kuo et al., Reference Kuo, Lee and Wang2020). Given the wide distribution of R. norvegicus, further studies are needed to assess the presence and potential public health relevance of these bacteria in South America.
In conclusion, molecular phylogenetic analyses demonstrate that Laelaps is polyphyletic, with a well-supported clade of cricetid-associated lineages clearly separated from murid-associated taxa. Within Cricetidae, phylogenetic structure is linked to host tribal affiliation, and the entanglement of morphologically distinct genera, Androlaelaps, Gigantolaelaps and Mysolaelaps, within the cricetid clade demonstrates that current generic boundaries in Laelapidae do not reflect evolutionary history. These results support the polyphyly of Laelaps and consequently challenge the wide distribution of this mite, highlighting the need for an integrative taxonomic revision of the species, combining molecular, morphological and ecological data.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182026102224.
Data availability statement
The original data generated in this study are available in GenBank (National Center for Biotechnology Information, https://www.ncbi.nlm.nih.gov/genbank/) under the following accession numbers: OQ241929, OQ241930, OQ241937, OQ241939–OQ241943, OR405892–OR405895, OR405898, PV952202, PV952205–PV952221.
Acknowledgements
We thank the Ministerio de Agroindustria (Buenos Aires Province), Ministerio de Ecología, Recursos Naturales Renovables y Turismo (Misiones Province), Ministerio de la Producción y Ambiente (Formosa Province), Dirección de Recursos Naturales (Corrientes Province), Dirección de Fauna (Chaco Province) and Dirección General de Recursos Naturales (Entre Ríos Province) for granting permits for mite collection.
We are grateful to Luis Giambelluca (Centro de Estudios Parasitológicos y de Vectores, CEPAVE, Argentina) and his family, to Nany Nuñez and her family, Elena Senattori and Fernando Glenza (Cátedra Libre de Soberanía Alimentaria, Universidad Nacional de La Plata) for logistical support. We thank Carla Yauris and Edgardo Rengifo for providing mite specimens from Peru, and Marina Ibáñez and Melisa Moncada (CEPAVE, Argentina) for their assistance with molecular procedures. This study forms part of the doctoral theses of M. Espinoza-Carniglia and E. Savchenko at the Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina.
Author contributions
All authors conceived the study. M.E.C. and E.S. curated the data and conducted the investigation. M.E.C. performed the formal analyses and visualization. M.E.C., E.S., U.F.J.P. and M.L. contributed to the methodology, including collecting mites, drafted the original manuscript, reviewed and edited the final version. M.L. acquired funding and was responsible for project administration, resources and supervision.
Financial support
The study was supported by the Universidad Nacional de La Plata, Argentina [N992], and the Consejo Nacional de Investigaciones Científicas y Técnicas [PIP 628] (both to M. Lareschi).
Competing interests
The authors declare there are no conflicts of interest.
Ethical standards
Not applicable.



 Open access
Open access