



Introduction
Parasites that may affect the central nervous system (CNS) and peripheral nervous system (PNS) of dogs (Canis familiaris) and cats (Felis catus) have long been documented, although the literature available for many of these agents remains heterogeneous and largely composed of descriptive isolated case reports or small case series. This fragmented evidence base has contributed to an incomplete understanding of their overall biological relevance, despite the capacity of several neurotropic parasites to induce significant alterations within neural tissues. In recent years, however, the role of these agents has been increasingly emphasized in parasitological and infectious disease literature, including authoritative reference texts (Sykes, Reference Sykes2022), reflecting a broader interest in their neuroinvasive properties and pathogenetic mechanisms.
While Toxoplasma gondii and Neospora caninum remain the most frequently considered parasites in neurological presentations, a broader spectrum of parasitic agents, ranging from unicellular protozoa to complex helminths and fungi, can invade or affect the nervous system (Dubey and Lindsay, Reference Dubey and Lindsay1996; Dubey, Reference Dubey2010; Vandevelde et al., Reference Vandevelde, Higgins and Oevermann2012).
From a biological standpoint, parasitic invasion or interaction with the CNS and PNS may occur through diverse mechanisms, including direct tissue migration, haematogenous dissemination and complex host–parasite interactions. The resulting neuronal damage or dysfunction is further compounded by multifactorial pathophysiological processes involving vascular alterations, immune-system dysregulation, toxin production or systemic metabolic effects, thereby further complicating the understanding and interpretation of these parasitic diseases (Bowman, Reference Bowman2014; Vitale and Foss, Reference Vitale and Foss2019; Willesen et al., Reference Willesen, Langhorn and Nielsen2022; Deng et al., Reference Deng, Fu, Elsheikha, Cao, Zhu, Wang, Zhang, Xie, Yao and Liu2024).
These pathogenetic processes frequently overlap with those observed in other neurological conditions, thereby hindering their recognition and contributing to the limited inclusion of parasitic agents in broader discussions of neurological disorders in small companion animals (Vandevelde et al., Reference Vandevelde, Higgins and Oevermann2012). Despite the increasing acknowledgement of the relevance of neurotropic parasites, the evidence currently available, apart from that concerning T. gondii and N. caninum, remains too limited and heterogeneous to support robust epidemiological inferences or fully systematic review.
The present review is primarily directed towards veterinary parasitologists and aims to synthesize current knowledge on neurotropic parasitic infections in small companion animals, extending beyond the well-recognized protozoa T. gondii and N. caninum. Particular emphasis is placed on the biological mechanisms underlying CNS and PNS invasion, parasite-associated pathogenetic pathways and diagnostic approaches that enable parasitological confirmation.
Mechanisms of CNS invasion and damage induced by parasites
Parasitic invasion and damage to the CNS and PNS in dogs and cats can occur through both direct and indirect mechanisms, contributing to a wide spectrum of neurological manifestations (Table 1).
Pathogenic mechanisms and neurotropisms of parasites affect the CNS/PNS of dogs and cats

Table 1 Long description
The table lists parasites affecting dogs and cats, grouped as helminths, arthropods, protozoans, and fungi, and outlines each agent’s neurotropism (rare to strong) with the main mechanisms causing CNS and/or PNS disease and the typical host species. Strong neurotropism is noted for Angiostrongylus cantonensis, Gurltia paralysans, Baylisascaris procyonis, ticks (via neurotoxins), Toxoplasma gondii, Neospora caninum, Cryptococcus neoformans/gattii, and Encephalitozoon cuniculi. Moderate neurotropism appears for Angiostrongylus vasorum, Cuterebra spp., Aspergillus spp. and Cladophialophora spp.. Many other agents are marked rare and are associated with ectopic migration or embolization (e.g., Aelurostrongylus abstrusus, Dirofilaria immitis, Spirocerca lupi, Taenia serialis), hematogenous spread (dimorphic fungi), or vascular/coagulopathic injury (e.g., Babesia rossi; also A. vasorum). Some entries emphasize non-migratory systemic effects that secondarily impact nerves, such as hypokalemia with Toxocara canis/cati and hypoglycaemia with Babesia rossi. Host ranges vary by agent, with several primarily in dogs (e.g., A. vasorum, N. caninum), several in cats or wild felids (e.g., G. paralysans, Cytauxzoon felis), and some listed in reservoir hosts with dogs or cats as incidental hosts. A few agents are flagged as having potential neurologic side effects related to treatment rather than infection alone.
CNS, central nervous system; PNS, peripheral nervous system.
a Accidental host.
b Neurological side-effects associated with drug administration for treatment.
Parasites like T. gondii, N. caninum, Encephalitozoon cuniculi and Gurltia paralysans are obligate parasites with a specific neurotropism, invading and replicating within neural tissues or the vascular system (Dubey and Lindsay, Reference Dubey and Lindsay1996; Dubey, Reference Dubey2010; Santaniello et al., Reference Santaniello, Cimmino, Dipineto, Agognon, Beguinot, Formisano, Fioretti, Menna and Oriente2021; Uribe et al., Reference Uribe, López-Osorio and Chaparro-Gutiérrez2021).
Conversely, other parasitic and fungal agents (e.g., D. immitis, Spirocerca lupi, Angiostrongylus vasorum, Aelurostrongylus abstrusus, Capillaria boehmi, Cryptococcus neoformans, Aspergillus spp., Cuterebra spp., Cytauxzoon felis, Babesia rossi and Leishmania infantum) may impact the CNS and PNS through ectopic migration of larvae, adults, spores and protozoa, typically in the presence of high parasitic burden or aberrant host–parasite interactions, as reported for Baylisascaris procyonis or Angiostrongylus cantonensis infections (Gavin et al., Reference Gavin, Kazacos, Tan, Brinkman, Byrd, Davis, Mets and Shulman2002; Vandevelde et al., Reference Vandevelde, Higgins and Oevermann2012; Jarvi and Prociv, Reference Jarvi and Prociv2021).
The pathways of CNS invasion may comprise translocation across the blood–brain barrier (BBB) through mechanisms such as transendothelial migration (e.g., A. vasorum), retrograde axonal transport along peripheral nerves (e.g., A. cantonensis) or direct contiguous dissemination from adjacent anatomical structures, particularly for parasites having a primary nasal localization the nasal sinuses, like C. boehmi, through the cribriform plate (Vandevelde et al., Reference Vandevelde, Higgins and Oevermann2012; Clark et al., Reference Clark, López, Levine, Cooper, Craig, Voges, Johnson and Porter2013; Bowman, Reference Bowman2014; Martins et al., Reference Martins, Tanowitz and Kazacos2015).
Once within the CNS and/or PNS, parasites may inflict direct damage to cells of the nervous tissue through cytopathic effects and mechanical disruption, resulting in necrosis and gliosis (Dubey, Reference Dubey2010; Kettenmann and Ransom, Reference Kettenmann and Ransom2013). They often trigger a cascade of inflammatory responses characterized by microglial activation, cytokine release and disruption of the neurovascular unit with consequent loss of BBB integrity and increased permeability (Kettenmann and Ransom, Reference Kettenmann and Ransom2013).
In some cases, parasite persistence within nervous tissue results in long-term neuroinflammation (as observed in the case of T. gondii), demyelination and neurodegeneration, which are associated with clinical manifestations such as seizures, behavioural abnormalities or focal neurological deficits including vestibular signs (e.g., head tilt, ataxia) linked to cerebellar or brainstem involvement (Dubey, Reference Dubey2010). In addition, even in the absence of detectable parasites in the tissue, indirect mechanisms may significantly contribute to CNS and/or PNS damage; this is exemplified by complement activation (e.g., L. infantum) or vascular involvement that can induce vasculitis, thrombosis, haemorrhage or ischaemia (e.g., A. vasorum, Cy. felis and B. rossi) (Veronesi et al., Reference Veronesi, Ravagnan, Cerquetella, Carli, Olivieri, Santoro, Pesaro, Berardi, Rossi, Ragni, Beraldo and Capelli2016; Smith et al., Reference Smith, Goddard, Boddapati, Brooks, Schoeman, Lack, Leisewitz and Ackerman2021; Willesen et al., Reference Willesen, Langhorn and Nielsen2022). Moreover, certain parasites or their symbiotic organisms can release neurotoxic metabolites, proteases or other secreted molecules that interfere with neurotransmission or glial cell homeostasis. For instance, the tick Ixodes holocyclus produces neurotoxins causing acute neuromuscular paralysis (Deng et al., Reference Deng, Fu, Elsheikha, Cao, Zhu, Wang, Zhang, Xie, Yao and Liu2024).
Severe parasitic infestations may lead to metabolic disorders such as hyponatremia, hypoglycaemia and acid–base imbalances, which can indirectly affect CNS function by altering neuronal excitability and increasing the risk of seizures or encephalopathy. This phenomenon has been observed, for example, in instances of heavy Toxocara canis burdens in puppies, where significant electrolyte imbalances can compromise normal neural activity; similarly, in canine babesiosis by B. rossi, hypoglycaemia is a recognized complication that can result in neurological manifestations (Keller et al., Reference Keller, Jacobson, Nel, De Clerq, Thompson and Schoeman2004; Bowman, Reference Bowman2014; Sarchahi and Borji, Reference Sarchahi and Borji2017).
Comparable neurological manifestations have also been described as sequelae of antiparasitic treatments for istance following melarsomine administration in canine D. immitis infections (Hettlich et al., Reference Hettlich, Ryan, Bergman, Marks, Lewis, Bahr, Coates, Mansell and Barton2003), neurotoxicity associated with diminazene treatment in canine babesiosis (Lee et al., Reference Lee, Yun and Song2025) and to the potential neuroinflammatory responses after anthelmintic drug administration in cases of A. cantonensis.
Despite growing understanding of the pathogenic mechanisms involved in damage, specific data regarding the neurotropism and anatomical predilection of most parasites in small companion animals remain limited. While T. gondii and N. caninum exhibit some localization patterns (e.g., cortical or cerebellar involvement) (Nishimura et al., Reference Nishimura, Kohara, Hiasa, Muroi, Yokoyama, Kida, Xuan, Furuoka and Nishikawa2013; Evans et al., Reference Evans, Strassmann, Lee and Sapolsky2014; El Saftawy et al., Reference El Saftawy, Shash, Aboulhoda, Arsanyos, Albadawi, Abou-Fandoud and Amin2021), such patterns are often absent or inconsistent in parasites characterized by erratic migratory behaviour or in those inflicting damage via indirect mechanisms. Consequently, lesion distribution is frequently unpredictable and clinical presentations are often variable and non-specific (Dubey and Lindsay, Reference Dubey and Lindsay1996; Dubey, Reference Dubey2010). Moreover, parasites with marked neurotropism are more frequently associated with evident and specific neurological syndromes; in contrast, such tropism is typically lacking in those parasites that induce sporadic neurological involvement, often representing incidental or secondary complications of the primary parasitic infection (Vandevelde et al., Reference Vandevelde, Higgins and Oevermann2012).
Helminths affecting the CNS and/or PNS
Metastrongyloids
Metastrongyloids are a group of nematodes of dogs and cats characterized by an indirect life cycle involving terrestrial or aquatic gastropods as intermediate hosts and occasionally paratenic hosts.
These parasites are implicated in a diverse array of clinical diseases in small companion animals, some of which manifest as severe neurological signs. In dogs, A. vasorum and A. cantonensis have been identified as causative agents of clinical presentations that range from cardiorespiratory impairment to both direct and indirect neurological signs. Conversely, in cats, G. paralysans and, less frequently, Ae. abstrusus have been associated with neurological disorders; however, the available data on these associations remain extremely limited.
A. vasorum (Order Strongylida, Family Angiostrongylidae) is a metastrongyloid nematode inhabiting the pulmonary arteries and right ventricle of domestic and wild canids (Schnyder et al., Reference Schnyder, Fahrion, Riond, Ossent, Webster, Kranjc, Glaus and Deplazes2010; Di Cesare and Traversa, Reference Di Cesare and Traversa2014). The parasite is traditionally endemic to Western Europe but has recently been reported in previously non-endemic areas, including Northwestern Italy, Eastern Europe and parts of Canada and South America (Olivieri et al., Reference Olivieri, Zanzani, Gazzonis, Giudice, Brambilla, Alberti, Romussi, Lombardo, Mortellaro, Banco, Vanzulli, Veronesi and Manfredi2017; Penagos-Tabares et al., Reference Penagos-Tabares, Lange, Chaparro-Gutiérrez, Taubert and Hermosilla2018).
Significant risk factors for canine infection include young age (typically <3 years), an outdoor lifestyle with exposure to habitats harbouring infected gastropods, predatory behaviour and travel to endemic regions (Di Cesare and Traversa, Reference Di Cesare and Traversa2014).
Clinical disease is primarily cardiorespiratory and results from vascular injury and inflammatory responses to eggs and migrating larvae in the lung parenchyma. A characteristic feature of infection is the development of coagulopathy, which may result in haemorrhagic diathesis, spontaneous mucosal bleeding, haematochezia or disseminated intravascular coagulation (DIC). These abnormalities are associated with thrombocytopenia, depletion of coagulation factors (notably II and V) and hyperfibrinolysis (Cury et al., Reference Cury, Lima, Guimarães and Carvalho2002; Schnyder et al., Reference Schnyder, Fahrion, Riond, Ossent, Webster, Kranjc, Glaus and Deplazes2010; Silva et al., Reference Silva, Hindenberg, Balzhäuser, Moritz, Hermosilla and Taubert2021; Willesen et al., Reference Willesen, Langhorn and Nielsen2022).
Neurological and ocular manifestations, although less frequently reported, are increasingly recognized as clinically relevant outcomes, occurring either in isolation or in association with respiratory and hemorrhagic signs (Colombo et al., Reference Colombo, Traversa, Grillotti, Pezzuto, De Tommaso, Pampurini, Schaper, Drake, Crisi, Russi, Ripamonti and Di Cesare2021). Neurological involvement is reported in approximately 4–16% of infected dogs and may result from 2 main mechanisms: (i) ectopic migration of larvae or adult parasites into the CNS, a relatively rare event (Oliveira-Júnior et al., Reference Oliveira-Júnior, Barçante, Barçante, Ribeiro and Lima2004); and (ii) haemorrhagic complications secondary to microvascular damage and parasite-induced coagulopathy, which represent the more common pathway (Wessmann et al., Reference Wessmann, Lu, Lamb, Smyth, Mantis, Chandler, Boag, Cherubini and Cappello2006; Schnyder et al., Reference Schnyder, Fahrion, Riond, Ossent, Webster, Kranjc, Glaus and Deplazes2010). Cerebral lesions, particularly affecting the frontal and temporal lobes, are frequently associated with seizure activity, whereas spinal haemorrhages may lead to paraparesis, paraplegia or ataxia.
A. cantonensis (Order Strongylida, Family Angiostrongylidae), commonly known as the ‘rat lungworm’, is a metastrongyloid nematode characterized by a pronounced neurotropism. It is the aetiological agent of canine neural angiostrongylosis, a condition that, although sporadic, has been increasingly reported in endemic regions such as Australia and Hawaii (Schnyder et al., Reference Schnyder, Fahrion, Riond, Ossent, Webster, Kranjc, Glaus and Deplazes2010; Griffin et al., Reference Griffin, Ezenwa and Cowie2025). Rats (Rattus norvegicus and Rattus rattus) act as definitive hosts, harbouring adult worms within the pulmonary arteries and right ventricle and shedding L1 larvae in the faeces, whereas dogs represent aberrant, non-permissive hosts (Jarvi and Prociv, Reference Jarvi and Prociv2021; Griffin et al., Reference Griffin, Ezenwa and Cowie2025). Infection occurs commonly in young, free-roaming or predatory dogs with potential exposure to gastropods or amphibians (Walker et al., Reference Walker, Spielman, Malik, Graham, Ralph, Linton and Ward2015). Dogs become infected following ingestion of gastropods (intermediate hosts) or paratenic hosts (e.g., frogs, crustaceans, planarians) containing infective L3 larvae. Following ingestion, larvae typically migrate via lymphovascular pathways and gain access to the CNS by ascending along peripheral nerves and spinal nerve roots, or directly via the subarachnoid space (Martins et al., Reference Martins, Tanowitz and Kazacos2015; Griffin et al., Reference Griffin, Ezenwa and Cowie2025).
Pathological damage arises through both mechanical and immunopathogenic mechanisms: migrating larvae induce direct neural injury and haemorrhage, while the release of larval metabolic, excretory and moulting antigens triggers intense eosinophilic meningoencephalomyelitis (Martins et al., Reference Martins, Tanowitz and Kazacos2015; Jarvi and Prociv, Reference Jarvi and Prociv2021). Moreover, rapid larval death, particularly following anthelmintic administration may trigger severe neuroinflammation, mirroring the Jarisch–Herxheimer-like reactions documented in human angiostrongyliasis and murine models after (Murphy and Johnson, Reference Murphy and Johnson2013).
Lesions are mainly detected in the spinal cord (especially in thoracolumbar segments) and brain, but may also involve peripheral nerves, resulting in a combination of central and peripheral neurological manifestations (Lunn et al., Reference Lunn, Lee, Smaller, MacKay, King, Hunt, Martin, Krockenberger, Spielman and Malik2012). Consequently, mixed upper and lower motor neuron signs are frequently reported, reflecting the multifocal involvement of the spinal cord and nerve roots. Affected dogs, most of which are young, typically present with an acute to subacute onset of paraparesis or proprioceptive ataxia, lumbosacral and tail-base hyperesthesia, urinary incontinence and progressive ascending paralysis (Lunn et al., Reference Lunn, Lee, Smaller, MacKay, King, Hunt, Martin, Krockenberger, Spielman and Malik2012; Walker et al., Reference Walker, Spielman, Malik, Graham, Ralph, Linton and Ward2015). Cerebral involvement may manifest as altered mentation, muscle atrophy, seizures and, in severe cases, coma. Neurological deterioration is often rapid, reflecting the severity of inflammation and the unpredictable nature of larval migration (Lee et al., Reference Lee, Pai, Churcher, Davies, Braddock, Linton, Yu, Bell, Wimpole, Dengate, Collins, Brown, Reppas, Jaensch, Wun, Martin, Sears, Šlapeta and Malik2020).
G. paralysans (Order Strongylida, Family Angiostrongylidae) is a neglected metastrongyloid nematode mainly distributed across South America, with additional reports from Tenerife Island and mainland Spain (Udiz-Rodríguez et al., Reference Udiz-Rodríguez, Garcia-Livia, Valladares-Salmerón, Dorta-Almenar, Martín-Carrillo, Martin-Alonso, Izquierdo-Rodriguez, Feliu, Valladares and Foronda1998; Gómez et al., Reference Gómez, Muñoz, Moroni, Mieres, Bernal, Rosenfeld, Taubert and Hermosilla2024). This parasite infects domestic cats and wild felids of the genus Leopardus, which serve as definitive hosts; adult nematodes reside within the venous system of the leptomeninges surrounding the spinal cord (Uribe et al., Reference Uribe, López-Osorio and Chaparro-Gutiérrez2021). Several aspects of the life cycle of G. paralysans remain poorly understood, including the route of transmission, the role of intermediate hosts (terrestrial or aquatic gastropods), larval development, migration pathway and larval shedding. Notably, neither eggs nor L1 larvae have been detected in faeces, blood or other body fluids of infected hosts, complicating diagnosis (Sépulveda-García et al., Reference Sépulveda-García, Gómez, Moroni, Muñoz and Muller2021; Uribe et al., Reference Uribe, López-Osorio and Chaparro-Gutiérrez2021).
G. paralysans shows a marked angio-neurotropism, invading the venous system of the leptomeninges, particularly in the spinal cord, mainly of the thoracic, lumbar and sacral spinal segments (Gómez et al., Reference Gómez, Moroni, Muñoz, Taubert, Hermosilla, Hirzmann and Rojas2021; Uribe et al., Reference Uribe, López-Osorio and Chaparro-Gutiérrez2021). The presence of adults within meningeal veins of the subarachnoid space triggers immune-mediated endothelial injury, leading to thrombophlebitis, thrombus formation, venous congestion and meningeal haemorrhages (Gómez et al., Reference Gómez, Moroni, Muñoz, Taubert, Hermosilla, Hirzmann and Rojas2021; Uribe et al., Reference Uribe, López-Osorio and Chaparro-Gutiérrez2021). The combined effects of thrombotic occlusions and inflammation cause a progressive myelopathy, which often leads to a fatal onset. The clinical signs in domestic cats include chronic, progressive pelvic limb ataxia; ambulatory paraparesis; proprioceptive deficits; hyperactive patellar reflexes (uni- or bilateral); muscular atrophy; and, in some cases, systemic signs such as diarrhoea, weight loss, coprostasis, and urinary and faecal incontinence (Alzate et al., Reference Alzate, Aranzazu, Alzate and Chaparro2011; Gómez et al., Reference Gómez, García, Maldonado, Pantchev, Taubert, Hermosilla, Moroni, Muñoz, Durán, Mieres and Ojeda2020, Reference Gómez, Moroni, Muñoz, Taubert, Hermosilla, Hirzmann and Rojas2021, Reference Gómez, Muñoz, Moroni, Mieres, Bernal, Rosenfeld, Taubert and Hermosilla2024; Rojas-Barón et al., Reference Rojas-Barón, Taubert, Hermosilla, Gómez, Moroni and Muñoz2022).
Ae. abstrusus (Order Strongylida, Family Angiostrongylidae) is the most common feline lungworm, typically inhabiting the terminal bronchioles and alveolar ducts of domestic cats, where it causes chronic respiratory disease (Traversa et al., Reference Traversa, Di Cesare and Conboy2010). Although Ae. abstrusus exhibits well-recognized respiratory tropism, recent evidence has demonstrated its ability to migrate ectopically within the CNS (Tinoco et al., Reference Tinoco, Morelli, de Farias Brito, Oliveira Pereira, Correia Oliveira, Diakou, Colombo, Frangipane Di Regalbono and Traversa2022). In fact, a documented case of ectopic migration and embolization of Ae. abstrusus was described in a young FeLV-positive cat, presented with acute neurological signs including ataxia, tetraparesis and myoclonus. The post-mortem examination revealed haemorrhagic meningoencephalomyelitis, with adult nematodes and larvae identified within cerebral and spinal blood vessels as well as the subarachnoid spaces (Tinoco et al., Reference Tinoco, Morelli, de Farias Brito, Oliveira Pereira, Correia Oliveira, Diakou, Colombo, Frangipane Di Regalbono and Traversa2022). Although extremely rare, such occurrences expand the recognized clinical spectrum of feline aelurostrongylosis and support the inclusion of Ae. abstrusus among the metastrongyloids nematodes capable of inducing neurological manifestations in cats.
Ascarids
Ba. procyonis (Order Ascaridida, Family Ascarididae) is the common roundworm of raccoons (Procyon lotor). Originally confined to North America, particularly the United States and southern Canada, where infection prevalence in raccoon populations frequently exceeds 80% in certain endemic areas (Gavin et al., Reference Gavin, Kazacos and Shulman2005; Blizzard et al., Reference Blizzard, Davis, Henke, Long, Hall and Yabsley2010). Over the last 2 decades, the parasite has been increasingly reported in Europe due to the introduction and establishment of P. lotor beyond its native range.
In Europe, B. procyonis has been documented in free-ranging populations in several countries, including Germany (notably in Hesse, Brandenburg, Saxony-Anhalt and North Rhine-Westphalia) (Rentería-Solís et al., Reference Rentería-Solís, Birka, Schmäschke, Król and Obiegala2018, Reference Rentería-Solís, Flores, Langner, Gawlowska, Grochow, Fietz, Birka, Król and Obiegala2025; Heddergott et al., Reference Heddergott, Steinbach, Schwarz, Anheyer-Behmenburg, Sutor, Schliephake, Jeschke, Striese, Müller, Meyer-Kayser, Stubbe, Osten-Sacken, Krüger, Gaede, Runge, Hoffmann, Ansorge, Conraths and Frantz2020; Reinhardt et al., Reference Reinhardt, Wassermann, Härle, Romig, Kurzrock, Arnold, Großmann, Mackenstedt and Straubinger2023), Austria, Luxembourg, the Netherlands, Switzerland, Denmark and, more recently, in western Poland and northern Italy (Lombardo et al., Reference Lombardo, Brocherel, Donnini, Fichi, Mariacher, Diaconu, Carfora, Battisti, Cappai, Mattioli and De Liberato2022). The progressive expansion of raccoon populations into urban and peri-urban areas has contributed to increased environmental contamination with infective eggs, particularly at habitual defecation sites (‘latrines’), which act as focal points for zoonotic transmission to humans and small companion animals (Graeff-Teixeira et al., Reference Graeff-Teixeira, Morassutti and Kazacos2016). Adult worms reside in the small intestine of raccoons, releasing a large number of highly resistant eggs (approximately 20 000–26 000 eggs per gram of faeces), which become infective within 11–14 days and can remain viable in the environment for several years (Kazacos et al. Reference Kazacos, Samuels, Pybus and Kocan2001; Gavin et al., Reference Gavin, Kazacos and Shulman2005).
Dogs may become infected through the ingestion of embryonated eggs present in the environment and may act either as accidental definitive hosts, developing patent intestinal infections with sporadic egg shedding (Graeff-Teixeira et al., Reference Graeff-Teixeira, Morassutti and Kazacos2016) or, more commonly, as aberrant hosts. Several factors may contribute to the risk of canine infection, including uncontrolled access to raccoon latrines or to areas contaminated with their faeces (a common occurrence in peri-domestic environments where raccoons live in synanthropic conditions, as well as outdoor lifestyles and behavioural traits such as exploratory or coprophagic behaviour, particularly in juvenile dogs). Additional risk factors include contact with contaminated soil, water, fomites and ingestion of infected paratenic hosts (Graeff-Teixeira et al., Reference Graeff-Teixeira, Morassutti and Kazacos2016).
In dogs, eggs containing L3 larvae of B. procyonis hatch in the intestine, penetrate the mucosa and disseminate hematogenously to multiple tissues, including the CNS, resulting in the neural larva migrans syndrome (Rudman et al., Reference Rudman, Kazacos, Storandt, Harris and Janovitz1996). Tissue invasion results in progressive and extensive mechanical damage to neural parenchyma with an inflammatory response characterized by severe eosinophilic meningoencephalitis, multifocal necrosis, haemorrhages, perivascular cuffing, microgliosis and disruption of the BBB (Thomas, Reference Thomas1988; Hazlett et al., Reference Hazlett, Cai, Sparling and You2018; Heller et al., Reference Heller, Arnold and Dreyfus2019). The time required for the development of clinical signs after infection in dogs is unknown. Still, in experimental infections, it is likely 2–4 weeks, depending on the infectious dose (Kazacos, Reference Kazacos2016).
Affected dogs show rapidly progressive neurological signs, including ataxia, compulsive circling, seizures, dysphagia, lethargy, recumbency, visual impairment, proprioceptive deficits, paresis and, in advanced stages, death (Thomas, Reference Thomas1988; Hazlett et al., Reference Hazlett, Cai, Sparling and You2018). The acute, and often hyperacute, progression reflects the aggressive nature of larval migration and the widespread distribution of lesions throughout the CNS (Hazlett et al., Reference Hazlett, Cai, Sparling and You2018). Although reports in cats are rare and infrequent, cats may also act as aberrant hosts under conditions of high environmental exposure, potentially developing comparable, albeit less frequent, neurological sequelae (Kazacos and Boyce, Reference Kazacos and Boyce1989).
Compared to B. procyonis, other ascarids such as T. canis and T. cati (Order Nematoda, Family Ascarididae), which represent classical models of viscerally migrating nematodes, contribute minimally to neurological disease in small companion animals. T. canis and T. cati are cosmopolitan roundworms that inhabit the small intestine of dogs and cats, respectively, particularly puppies and kittens. Adults release eggs that become infective (containing L3 larvae) in the environment within 10–12 days and remain viable for years. Infection occurs through ingestion of infective eggs, transplacental transmission (in dogs), transmammary transmission (in both dogs and cats) or ingestion of paratenic hosts (Schwartz et al., Reference Schwartz, Bidaisee, Fields, Macpherson and Macpherson2021). Following ingestion, larvae penetrate the intestinal wall and migrate through the liver and lungs via the portal and systemic circulation, causing multifocal granulomatous and eosinophilic inflammation. Pulmonary and hepatic lesions due to larval migration are well documented in both dogs and cats, whereas the CNS does not appear to be a primary target. In adult animals, larvae tend to encyst within somatic tissues in a hypobiotic state, serving as a reservoir for transplacental transmission to offspring (Bowman, Reference Bowman2014).
Although T. canis and T. cati are well known for their extensive larval migration, documented CNS involvement in naturally infected dogs and cats is absent. Evidence from experimental murine models indicates that T. canis larvae can migrate into the CNS (Salama et al., Reference Salama, Elgendy, Elmahy, Eltantawy, Seleem, Elgohary, Docmac, Al Mwafy, Almeldin and Sharaf2025); however, extrapolation of these findings to the canine or feline host should be made with caution. In humans, particularly in children, larval migration of T. canis has been associated with visceral and ocular larva migrans syndromes, and in some cases with eosinophilic meningitis or encephalomyelitis, demonstrating its capacity to elicit inflammatory reactions within the CNS (Magnaval et al., Reference Magnaval, Glickman, Dorchies and Morassin2001; Sánchez et al., Reference Sánchez, García and Nicoletti2018).
Isolated reports of neurological signs in heavily infected puppies; however, such cases are anecdotal are observed, but a direct causal relationship with larval neuroinvasion remains unproven. In the cases described, neurological deficits are more plausibly interpreted as secondary to severe systemic and metabolic disturbances associated with massive intestinal parasitism, i.e. vomiting, diarrhoea, anorexia and marked hypokalaemia (Sarchahi and Borji, Reference Sarchahi and Borji2017). The resulting electrolyte imbalance may impair Na⁺/K⁺-ATPase activity in muscle fibres, leading to weakness or flaccid paralysis. Accordingly, such manifestations should be regarded as indirect metabolic consequences of overwhelming infection rather than as evidence of primary neurotropic behaviour in the definitive host.
Spiruroids
D. immitis (Order Spirurida, Family Onchocercidae) is a worldwide nematode transmitted through the blood meal of Culicidae (genus Aedes and Culex), which inoculates the infective L3 larvae into dogs, which act as primary definitive hosts. Cats can also be infected but are commonly non-fully competent hosts, in which microfilariae are rarely detected. Adult worms are considerable in length (up to 30 cm in females) and live in the right heart and pulmonary arteries, where they release circulating microfilariae (L1 larval stage) into the bloodstream.
Historically, D. immitis was primarily confined to temperate and tropical regions. The parasite’s geographical range of distribution has expanded in recent decades, due to several drivers, such as climate change, increased movement of domestic dogs and introduction of exotic competent vector (i.e., Aedes albopictus). In endemic areas, prevalence in canine populations can reach up to 80% (e.g., in the Mediterranean areas), depending on vector density and local prophylactic practices, whereas in cats, the infection is less frequent (Capelli et al., Reference Capelli, Poglayen, Bertotti, Giupponi and Martini1996; McCall et al., Reference McCall, Genchi, Kramer, Guerrero and Venco2008; Genchi et al., Reference Genchi, Mortarino, Rinaldi, Cringoli, Traldi and Genchi2011). Factors influencing infection risk in both dogs and cats include exposure to infected mosquitoes, lack of chemoprophylaxis, an outdoor lifestyle and residence in endemic regions (McCall et al., Reference McCall, Genchi, Kramer, Guerrero and Venco2008; Genchi et al., Reference Genchi, Mortarino, Rinaldi, Cringoli, Traldi and Genchi2011).
In dogs, the infection consists of a cardiopulmonary disease with clinical signs secondary to pulmonary hypertension, pulmonary thromboembolism and right-sided congestive heart failure (i.e., cough, dyspnoea, exercise intolerance, fatigue and caval-vein syndrome). Aberrant or ectopic migration of D. immitis to atypical sites has been sporadically documented in vascular structures such as the hepatic and azygos veins; left heart chambers; the aorta, iliac and femoral arteries; and extravascular sites, including the anterior chamber of the eye and the CNS. In the CNS, larvae and adults have been documented within intracranial arteries (especially in the prosencephalic region), the ventricular system and the subarachnoid or epidural spaces of the spinal cord (Beasley et al., Reference Beasley, Hiebert, Alcott, Jeffery, LaFoon-Jones and Gambino2021). Cats are less permissive hosts compared to dogs and more frequently exhibit aberrant larval migration and atypical presentations, including sudden death, due to their heightened susceptibility to low parasite burdens (Favole et al., Reference Favole, Cauduro, Opreni, Zanzani, Albonico, Manfredi, Cantile and Lorenzo2013).
Historically, extensive neurological case reports in dogs are described from the 1970s and 1980s. Neurological signs secondary to massive D. immitis infestations (where high parasitic burdens increased the likelihood of aberrant migration) were documented and more frequently observed in small-breed dogs, in which even moderate parasite loads could result in clinically evident neurological signs (Luttgen and Crawley, Reference Luttgen and Crawley1981; Shires et al., Reference Shires, Turnwald, Qualls and King1982; Blass et al., Reference Blass, Holmes and Neer1989). Currently, improved prophylactic measures and early detection have markedly reduced the prevalence of severe neurological presentations; nonetheless, ectopic CNS migration remains sporadically reported, particularly in small-breed dogs such as Yorkshire Terriers and Chihuahuas, which appear predisposed to such atypical localizations (Healey et al., Reference Healey, Reaugh and Davidson2003; Bonawandt et al., Reference Bonawandt, Berg, Joseph and Stefanacci2017). Moreover, 3 cases of neurologic complications after melarsomine dihydrochloride treatment have been described (Hettlich et al., Reference Hettlich, Ryan, Bergman, Marks, Lewis, Bahr, Coates, Mansell and Barton2003). In these cases, the authors speculated that the compound migrated from the injection site via fascial planes, resulting in secondary extensive inflammation and necrosis of the spinal epidural fat, which subsequently led to several neurologic deficits, including paraparesis and hind limb paralysis (Hettlich et al., Reference Hettlich, Ryan, Bergman, Marks, Lewis, Bahr, Coates, Mansell and Barton2003).
Neurological signs associated with D. immitis infection are primarily attributed to compressive myelopathy, localized inflammatory responses and vascular damage (Beasley et al., Reference Beasley, Hiebert, Alcott, Jeffery, LaFoon-Jones and Gambino2021). Reported neurological signs include hyperesthesia (most commonly cervical), paresis (hemi-, para- or tetraparesis), ataxia, circling behaviour, blindness, epileptic seizures and, in severe cases, death. Clinical signs can appear acutely or progress gradually, depending on the anatomical dislocation of the parasites and the extent of parasitic migration. Routine physical and biochemical examinations frequently fail to reveal distinctive abnormalities, thereby complicating early diagnosis.
Neuromuscular manifestations, including hind limb lameness, paraesthesia and seizures, may also develop in dogs with systemic arterial dirofilariasis and thromboembolism. In cats, D. immitis infection can result in sudden death without premonitory signs; however, the most common clinical signs stem from the development of a syndrome known as heartworm-associated respiratory disease (Atkins et al., Reference Atkins, De Francesco, Coats, Sidley and Keene2000). Neurological signs such as seizures, ataxia, para-paresis and blindness have also been reported in association with migration of adult worms or microfilariae, particularly within the brain and, less frequently, the spinal cord. The small size of spinal vessels in cats prevents filarial migration (Ader, Reference Ader1979; Miura et al., Reference Miura, Kanamoto, Morita and Maseki2001; Favole et al., Reference Favole, Cauduro, Opreni, Zanzani, Albonico, Manfredi, Cantile and Lorenzo2013).
S. lupi (Order Spirurida, Family Spirocercidae) is a further spiruroid nematode infecting dogs mostly in tropical and subtropical areas that may cause uncommon aberrant migration of unknown incidence into the spinal cord (Chai et al., Reference Chai, Yas, Brenner, Rojas, Konstantin, Klainbart and Shamir2018). S. lupi typically colonized the caudal esophageal wall, inducing granulomatous nodule formation (Bailey, Reference Bailey1972; Chai et al., Reference Chai, Yas, Brenner, Rojas, Konstantin, Klainbart and Shamir2018). While the parasite exhibits a well-characterized esophageal tropism, aberrant migration is documented in multiple anatomical sites, including thoracic organs, the gastrointestinal and urinary tracts and subcutaneous tissues (Babero et al., Reference Babero, Fawzi and Al-Dabagh1965; Georgi et al., Reference Georgi, Han and Hatrick1980; Harrus et al., Reference Harrus, Harmelin, Markovics and Bark1996). Involvement of the CNS has been reported exclusively within the spinal cord, encompassing extradural and most frequently intradural cases (Tudury et al., Reference Tudury, Graca, Arias, Da Luz Pereira and Ballarin1995; Dvir et al., Reference Dvir, Perl, Loeb, Shklar-Hirsch, Chai, Mazaki-Tovi, Aroch and Shamir2007; Chai et al., Reference Chai, Shelef, Brenner, Dogadkin, Aroch and Shamir2008; Casoy et al., Reference Casoy, Srugo, Barnoon, Segal, Zafrany, Segev and Shipov2025). The segmental distribution within the spinal cord appears variable and unpredictable, indicating an aberrant migratory pattern, with no evidence of brain or brainstem involvement. Based on clinical observations, the combination of acute, progressive and painful asymmetric paresis or paralysis; presence of eosinophils in cerebrospinal fluid (CSF); and absence of compressive spinal lesions can be considered sufficient to support a presumptive diagnosis in dogs from endemic areas.
Capillarioids
Capillaria (syn. Eucoleus) boehmi (Order Trichocephalida, Family Trichuridae) is a capillarid nematode that parasitizes the nasal cavities of dogs and wild canids, i.e., foxes, that represent the natural reservoir of the infection (Veronesi et al., Reference Veronesi, Lepri, Morganti, Di Palma, Mechelli, Moretti and Traversa2013; Di Cesare et al., Reference Di Cesare, Morelli, Morganti, Simonato, Veronesi, Colombo, Berlanda, Lebon, Gallo, Beugnet and Traversa2021). Transmission is direct, through ingestion of typical barrel-shaped eggs containing the infective L1 stage; however, transmission via earthworms acting as intermediate hosts have also been suggested (Traversa et al., Reference Traversa, Di Cesare and Conboy2010). Nasal eucoleosis is a worldwide parasitosis, but crowded environments (e.g., shelter) may be associated with a higher infection prevalence. Most C. boehmi infections are subclinical; however, heavy parasite loads can cause upper respiratory tract disease, including sneezing or nasal itching, reverse sneezing, mucopurulent nasal discharge and some degree of hyposmia or anosmia (Veronesi et al., Reference Veronesi, Lepri, Morganti, Di Palma, Mechelli, Moretti and Traversa2013, Reference Veronesi, Morganti, Di Cesare, Schaper and Traversa2014; Alho et al., Reference Alho, Mouro, Pissarra, Murta, Lemos, Gomes, Lima and de Carvalho2016). A case of nasal capillariosis presenting with convulsive symptoms was described secondary to an intracranial granolumatous lesion, where eggs of C. boehmi were recorded. Direct migration of C. boehmi eggs through the cribriform plate was considered to be the most likely pathogenic mechanism underlying the intracranial infection observed in this case (Clark et al., Reference Clark, López, Levine, Cooper, Craig, Voges, Johnson and Porter2013).
Cestodes
Taenia serialis (Order Cyclophyllidea, family Taeniidae) is a taeniid cestode whose metacestode stage, namely Coenurus serialis, develops in subcutaneous tissues of lagomorphs that act as intermediate hosts, while canids represent definitive hosts (Jull et al., Reference Jull, Browne, Boufana, Schöniger and Davies2012). Transmission to accidental hosts such as cats occurs via faecal–oral ingestion of embryonated eggs shed by infected dogs. Although T. serialis is considered widespread globally, feline cerebral coenurosis is extremely sporadic, with cases reported to date are mainly from Australia and North America (Hayes and Creighton, Reference Hayes and Creighton1978; Smith et al., Reference Smith, Bailey, Baker and Kock1988; Slocombe et al., Reference Slocombe, Arundel, Labuc and Doyle1989; Huss et al., Reference Huss, Miller, Corwin, Hoberg and O’Brien1994; Jull et al., Reference Jull, Browne, Boufana, Schöniger and Davies2012). In the affected cats, intracranial coenuri develops within the cerebral hemispheres and represents the outcome of aberrant larval migration within the CNS (Jull et al., Reference Jull, Browne, Boufana, Schöniger and Davies2012).
The infection is characteristically chronic, with neurological impairment mediated primarily by the progressive expansion of the coenurus over several months, which results in sustained increase in intracranial pressure, cerebral parenchymal displacement, and in advanced cases, midline shift and brain herniation (Jull et al., Reference Jull, Browne, Boufana, Schöniger and Davies2012). This pathophysiological progression accounts for the rapid terminal clinical decline and closely parallels the mass-effect-driven deterioration described in Taenia solium neurocysticercosis in humans (Rajshekhar, Reference Rajshekhar2010). Clinical outcomes become evident only once the cyst attains a volume sufficient to induce neurological dysfunction; at that point, cats typically exhibit an acute to subacute deterioration over 1–2 weeks (Jull et al., Reference Jull, Browne, Boufana, Schöniger and Davies2012). Reported neurological signs include marked obtundation, non-ambulatory tetraparesis, bilateral cranial nerve deficits and diffuse deficits in postural reactions.
Protozoans affecting the CNS and/or PNS
Cyst-forming parasites
T. gondii and N. caninum (Order Eucoccidiorida, Family Sarcocystidae) are widespread protozoan parasites with complex life cycles. Cats and wild felids are the definitive hosts of T. gondii, while most warm-blooded animals, including humans, act as intermediate hosts (Dubey, Reference Dubey2010). N. caninum exhibits a relatively narrow host range: domestic dogs and some wild canids are definitive hosts, and ruminants, equids, camelids, raptors and rodents are intermediates (Cedillo et al., Reference Cedillo, Martínez, Santacruz, Banda and Morales2008; Zanet et al., Reference Zanet, Veronesi, Giglia, Baptista, Morganti, Mandara, Vada, De Carvalho and Ferroglio2023). Both parasites have sporozoites, tachyzoites and bradyzoites as infectious stages, transmitted via ingestion of oocysts or tissue cysts, or transplacentally. Horizontal transmission via predation predominates for T. gondii, whereas vertical transmission is the primary route for N. caninum in dogs (Barber and Trees, Reference Barber and Trees1998; Dubey, Reference Dubey2010).
Infections from T. gondii and N. caninum are globally distributed, with prevalence influenced by population dynamics and exposure risks. In cats, risk factors include outdoor access, fighting, hunting and ingestion of raw/undercooked meat (Jokelainen et al., Reference Jokelainen, Simola, Rantanen, Näreaho, Lohi and Sukura2012; Kokkinaki et al., Reference Kokkinaki, Saridomichelakis, Mylonakis, Leontides and Xenoulis2023). In dogs, higher prevalence is observed among working or hunting animals, those fed raw or homemade diets or coprophagic individuals with exposure to free-ranging cats (Machacova et al., Reference Machacova, Bartova, Sedlak, Slezakova, Budikova, Piantedosi and Veneziano2016; Dini et al., Reference Dini, Stancampiano, Poglayen and Galuppi2024).
Ingestion of oocysts or tissue cysts by dogs and cats releases the infective stages (sporozoites or bradyzoites), which transform into tachyzoites and replicate within metabolically active host cells, causing tissue damage via direct cytolysis and parasite-derived toxins. During the acute phase, the intestinal epithelium and associated lymph nodes are the first to be affected. Tachyzoites then disseminate through lymphohematogenous routes to multiple organs, including the CNS. Within the CNS, rapid tachyzoite replication destroys neural and glial cells, resulting in multifocal necrotic foci randomly distributed in grey and white matter. Tachyzoites are observed freely, within macrophages, or inside neural cells (Dubey, Reference Dubey2010; Silva and Machado, Reference Silva and Machado2016). The subsequent cell-mediated immune response promotes conversion of tachyzoites into bradyzoites, forming persistent tissue cysts (70–100 µm) within 3 months. In T. gondii, tissue cysts localize in neural tissue and striated muscles, while in N. caninum, cysts are restricted to the CNS. Factors such as immunosuppression, comorbidities or physiological stress (e.g., pregnancy) can trigger cyst rupture, releasing bradyzoites that rapidly convert to tachyzoites, thereby causing local or systemic disease (Silva and Machado, Reference Silva and Machado2016; Sanchez and Besteiro, Reference Sanchez and Besteiro2021). Chronic phase reactivation differs between the parasites: T. gondii typically induces long-lasting IFN-γ–mediated immunity, rarely disrupted by severe immunosuppressive diseases (e.g., canine distemper, FIV, FeLV, FIP) (Dubey et al., Reference Dubey, Murata, Cerqueira-Cézar, Kwok, Yang and Su2020a, Reference Dubey, Cerqueira-Cézar, Murata, Kwok, Yang and Su2020b), whereas N. caninum reactivates easily, even after hormonal changes or immunosuppressive treatments (Silva and Machado, Reference Silva and Machado2016; Henry et al., Reference Henry, Rees, Morrison and Woods2025).
The clinical expression of toxoplasmosis in dogs and cats is influenced by host immune status, infectious dose, coinfections and the geographical distribution of parasite genotypes, although the contribution of genotype to disease outcome remains debated (Calero-Bernal and Gennari, Reference Calero-Bernal and Gennari2019). In cats, T. gondii infection is usually subclinical, as immunocompetent hosts efficiently control parasite multiplication. Clinical disease typically results from reactivation of latent infection rather than new exposure, particularly under immunosuppressive conditions such as FeLV, FIV, FIP or iatrogenic immunosuppression from high-dose glucocorticoids, cyclosporin, oclacitinib or chemotherapy (Murakami et al., Reference Murakami, Mori, Takashima, Nagamune, Fukumoto, Kitoh, Sakai and Maruo2018; Lo Piccolo et al., Reference Lo Piccolo, Busch, Palić, Geisen, Hartmann and Unterer2019; Moore et al., Reference Moore, Burrows, Malik, Ghubash, Last and Remaj2022; Fietz et al., Reference Fietz, Grochow, Schares, Töpfer and Heilmann2023). While any organ can be affected, the CNS, lungs, liver, muscles and eyes are most commonly involved.
Neurological signs related to toxoplasmosis manifestations vary depending on the site and extent of the lesions. They may include seizures, head tremors, anisocoria, torticollis, paresis, progressive pelvic limb ataxia, vestibular dysfunction, nystagmus and polyneuropathy (Alves et al., Reference Alves, Gorgas, Vandevelde, Gandini and Henke2011; Mari et al., Reference Mari, Shelton and De Risio2016; Tyroller et al., Reference Tyroller, Haas, Posch, Hettlich, Schwandt and Pfleghaar2023). Focal or multifocal granulomatous encephalitis and myelitis are characteristic pathological findings, sometimes leading to the formation of intracranial granulomas or extradural spinal masses detectable through advanced imaging techniques (Falzone et al., Reference Falzone, Baroni, De Lorenzi and Mandara2008; Tyroller et al., Reference Tyroller, Haas, Posch, Hettlich, Schwandt and Pfleghaar2023). Congenital infection can also result in severe CNS involvement in kittens, with tremors, ataxia or seizures, often accompanied by hepatomegaly and interstitial pneumonia (Calero-Bernal and Gennari, Reference Calero-Bernal and Gennari2019; Crouch et al., Reference Crouch, Mittel, Southard, Cerqueira-Cézar, Murata, Kwok, Su and Dubey2019).
In dogs, toxoplasmosis is relatively uncommon but can present with severe clinical forms in puppies, immunocompromised individuals or those debilitated by other infectious diseases such as distemper virus or monocytic ehrlichiosis (Dubey et al., Reference Dubey, Murata, Cerqueira-Cézar, Kwok, Yang and Su2020a; Sandu et al., Reference Sandu, Deak, Turcitu, O’Brien and Mircean2025). The acute phase may include fever, lethargy, anorexia and weight loss, while specific signs depend on the organs involved. Neurological involvement is considered to be the most frequent and characteristic presentation: infected dogs may show ataxia, tremors, and flaccid or spastic paralysis (Migliore et al., Reference Migliore, La Marca, Stabile, Di Marco Lo Presti and Vitale2017; Chen et al., Reference Chen, Boulay, Chong, Ho, Chan, Ong, Fernandez, Chang and Yap2023; Morganti et al., Reference Morganti, Rigamonti, Brustenga, Calgaro, Angeli, Moretta, Diaferia and Veronesi2024a). A recent study evaluated the correlation between clinical epilepsy and T. gondii seropositivity but did not support a causal role of the parasite in canine epilepsy (Morganti et al., Reference Morganti, Rigamonti, Marchesi, Maggi, Angeli, Moretta, Brustenga, Diaferia and Veronesi2024b). In more severe cases, multifocal encephalomyelitis and vestibular or cerebellar signs may occur due to necrotic foci and diffuse inflammation within the CNS (Dubey et al., Reference Dubey, Murata, Cerqueira-Cézar, Kwok, Yang and Su2020a). Vertical transmission, although rare in dogs, has been documented in several litters born to bitches infected during pregnancy, leading to foetal resorption, abortion, neonatal death or new-born puppies containing congenital encephalitis or necrotizing pneumonia (Calero-Bernal and Gennari, Reference Calero-Bernal and Gennari2019).
Behavioural studies in rodents have demonstrated that T. gondii can manipulate host neuronal circuits to enhance its transmission potential. Chronically infected animals exhibit a loss of innate aversion to feline predators, thereby increasing the likelihood of predation and completion of the parasite’s life cycle (Vyas et al., Reference Vyas, Kim, Giacomini, Boothroyd and Sapolsky2007). Although less studied in domestic dogs and cats, similar behavioural changes, including increased risk-taking and anxiety-related behaviour, have been proposed, reflecting the subtle neurobiological effects of chronic infection (Simion et al., Reference Simion, Fernoaga and Codreanu2019). However, direct evidence in small companion animals remains limited.
A single clinical case has been reported in a female Collie infected with T. gondii that exhibited noise hypersensitivity and abnormal phobic responses (Papini et al., Reference Papini, Marangi, Mancianti, Giangaspero and Barchiesi2009). Moreover, in dogs habitually fed raw meat, a stronger association has been observed between T. gondii seropositivity and anxious behavioural states, suggesting a potential link among diet, parasite exposure and behaviour (Dini et al., Reference Dini, Marliani, Amadei, Tosco, Cavallini, Accorsi and Galuppi2025).
More consistent observations come from studies in wild canids. Notably, T. gondii-seropositive red foxes and grey wolves have demonstrated reduced neophobia, increased exploratory behaviour in anthropogenic environments and increased risk-prone behaviours, such as approaching roads or human settlements (Milne et al., Reference Milne, Fujimoto, Bean, Peters, Hemmington, Taylor, Fowkes, Martineau, Hamilton, Walker, Mitchell, Léger, Priestnall and Webster2020; Meyer et al., Reference Meyer, Cassidy, Stahler, Brandell, Anton, Stahler and Smith2022). These behaviour alterations have been interpreted as potential consequences of parasite-induced modulation of dopaminergic neural circuits, analogous to mechanisms described in experimentally infected rodents. It has also been hypothesized that coinfections with neurotropic viruses, including fox circovirus or canine distemper virus, may modulate the neuroinflammatory responses and exacerbate the behavioural alterations associated with T. gondii (Milne et al., Reference Milne, Fujimoto, Bean, Peters, Hemmington, Taylor, Fowkes, Martineau, Hamilton, Walker, Mitchell, Léger, Priestnall and Webster2020; Meyer et al., Reference Meyer, Cassidy, Stahler, Brandell, Anton, Stahler and Smith2022). Although a causal relationship has not yet been definitively established, these observations suggest that the parasite may exert broader and more complex neurobiological effects than previously recognized.
Neurological signs of N. caninum infection in dogs can involve both the CNS and PNS, with the latter being more frequently affected. Peripheral neurological disorders are typically associated with congenital infection and are most commonly observed in puppies under 6 months of age (Lyon, Reference Lyon2010). Clinical signs may be evident at birth but usually manifest within the first 5–8 weeks of life. The infection initially targets the lumbosacral plexus, resulting in unilateral pelvic limb weakness that rapidly progresses to the contralateral limb. This is followed by neurogenic atrophy and characteristic rigid hyperextension (arthrogryposis) of 1 or both hind limbs (Dubey et al., Reference Dubey, Lindsay and Lappin2009). This characteristic hind limb paresis and rigid paralysis results from limb contracture caused by a disparity between normal skeletal growth and abnormal shortening of denervated muscles due to neurogenic atrophy. Affected puppies generally retain deep pain perception and consciousness. At the same time, some may enter a chronic stable phase, medium- to large-breed dogs are often euthanized due to severe disability and management difficulties.
CNS involvement is less common in neosporosis but represents a severe and often prognostically unfavourable manifestation. Central neurological disorders are characterized by reduced consciousness, head tremors, cerebellar ataxia, hypermetria and cervical hyperesthesia, reflecting meningoencephalitis and necrotizing cerebellitis (Garosi et al., Reference Garosi, Dawson, Couturier, Matiasek, de Stefani, Davies, Jeffery and Smith2010; Calero-Bernal and Gennari, Reference Calero-Bernal and Gennari2019; Didiano et al., Reference Didiano, Monti, Taeymans and Cherubini2020; Henry et al., Reference Henry, Rees, Morrison and Woods2025).
Vector-borne protozoans
A heterogeneous group of apicomplexan and kinetoplastid parasites transmitted by hematophagous arthropods (e.g., hard ticks and sand flies) can be responsible for a wide spectrum of systemic and haematological diseases in dogs and cats and, in some cases, may also affect the nervous system through direct invasion, vascular injury or immune-mediated mechanisms. In dogs, L. infantum and B. rossi can lead to multisystemic or haemolytic disease that may give rise to secondary neurological manifestations; however, in cats, Cy. felis may induce acute to hyperacute illness with high mortality, characterized by microvascular obstruction within the CNS.
L. infantum (Order Kinetoplastida, Family Trypanosomatidae) is a vector-borne pathogen transmitted by the bite of infected sand flies of the genera Phlebotomus (Old World) and Lutzomyia (New World). Dogs represent the primary reservoir hosts, playing a central role in zoonotic transmission and parasite maintenance in endemic areas of the Mediterranean basin, the Middle East and parts of Central and South America (Dantas-Torres et al., Reference Dantas-Torres, Solano-Gallego, Baneth, Ribeiro, de Paiva-cavalcanti M and Otranto2012). Cats act as secondary reservoirs; although infections and clinical cases are increasingly reported, particularly in endemic regions, their epidemiological significance remains limited to date (Vioti et al., Reference Vioti, da Silva, Galvis-Ovallos, Alves, da Silva, Leonel, Pereira, Benassi, Spada, Maia, Galati, Starke-Buzetti and Oliveira2022). Alternative transmission routes, including transplacental, venereal and blood transfusion-associated transmission, have been documented in dogs.
Clinically, canine leishmaniosis (CanL) ranges from subclinical infection to severe multisystemic disease, with severity largely dependent on the host immune response. Much of CanL pathogenesis is linked to the deposition of soluble circulating immune complexes (sCICs) and complement activation, leading to small-vessel vasculitis affecting the kidneys, anterior uvea, dermis, nasal mucosa, synovial membranes and the nervous system, including the choroid plexus (Day, Reference Day2011). This immunopathological mechanism explains the principal systemic manifestations of CanL, including vasculitis, glomerulonephritis, uveitis, arthritis, and exfoliative or ulcerative dermatitis, particularly involving periocular, nasal and auricular regions. In cats, infection is frequently subclinical or self-limiting, although both visceral and dermatological lesions have been described.
Involvement of the CNS and PNS is uncommon in CanL and has been primarily reported in case reports and small series of cases (Garcia-Alonso et al., Reference Garcia-Alonso, Nieto, Blanco, Requena, Alonso and Navarrete1996; Prasad and Sen, Reference Prasad and Sen1996). Neurological signs are observed in approximately 5–10% of infected dogs and are generally associated with advanced systemic disease (Giannuzzi et al., Reference Giannuzzi, Ricciardi, De Simone and Gernone2017; Gernone et al., Reference Gernone, Otranto, Mendoza-Roldan, Uva, Matiasek and Zatelli2025). However, recent evidence has demonstrated multifocal meningoencephalitis and detectable parasite DNA in the brain of dogs without neurological signs and living in an endemic region for leishmaniosis, confirming that CNS involvement can occur even in the absence of overt systemic signs (da Rosa et al., Reference da Rosa, Ries, Cargnelutti, Masuda and Vogel2024).
CNS lesions may result from direct parasitic invasion, with amastigotes occasionally detected in meningeal macrophages, endothelial cells or choroid plexus epithelium cells, supporting a ‘Trojan horse’ hypothesis, whereby infected monocytes traverse the BBB (Oliveira et al., Reference Oliveira, Boechat, Mendes Junior, Madeira, Ferreira, Figueiredo, Campos, de Carvalho Rodrigues, Carvalhaes de Oliveira, Amendoeira and Menezes2017). More frequently, CNS lesions arise from immune-mediated and vascular mechanisms, including deposition of sCIC, complement activation and small-vessel vasculitis associated with perivascular inflammatory infiltrates, endothelial swelling, microthrombosis and focal ischaemic necrosis (Gernone et al., Reference Gernone, Otranto, Mendoza-Roldan, Uva, Matiasek and Zatelli2025). Systemic inflammation further increases BBB permeability through metalloproteinase activation and vascular damage, facilitating the infiltration of immune mediators and autoreactive lymphocytes, thereby amplifying neuroinflammation and gliosis.
Peripheral neuropathies in CanL may arise from sCIC deposition on the endoneurial vascular endothelium, disruption of the blood–nerve barrier or an aberrant host immune response leading to demyelination and/or axonal degeneration (Ikeda et al., Reference Ikeda, Laurenti, Corbett, Feitosa, Machado and Perri2007; Gernone et al., Reference Gernone, Otranto, Mendoza-Roldan, Uva, Matiasek and Zatelli2025). Clinically, symmetric polyneuropathy characterized by paresis, decreased postural reactions, reduced withdrawal reflexes and neurogenic pelvic limb muscle atrophy has been reported (Márquez et al., Reference Márquez, Pedregosa, López, Marco-Salazar, Fondevila and Pumarola2013). Additionally, infected macrophages may occasionally form tumour-like masses or inflammatory granulomas at paravertebral, intramedullary or intracranial sites, contributing to neurological damage manifested by ataxia, paralysis and tremors (Cauduro et al., Reference Cauduro, Favole, Lorenzo, Simonetto, Barda, Cantile and Asperio2011; José-López et al., Reference José-López, de la Fuente, Pumarola and Añor2014).
While neurological leishmaniosis is rare in cats, cases associated with systemic disease have been reported, presenting as lethargy, disorientation, anisocoria and proprioceptive deficits (Fernández Gallego et al., Reference Fernández Gallego, Feo Bernabé, Dalmau, Esteban Saltiveri, Font, Leiva, Ortuñez Navarro, Peña, Tabar, Real Sampietro, Saló, Lloret and Bardagí2020).
Cy. felis (Order Piroplasmida, Family Theileriidae) is a highly pathogenic tick-borne protozoan that infects domestic cats in North America, where wild felids such as bobcats (Lynx rufus) and pumas (Puma concolor) act as asymptomatic reservoir hosts. In Europe, a closely related species, currently referred to as Cy. sp. has been described; it recognizes the European wildcat (Felis silvestris silvestris) and Iberian lynx (Lynx pardinus) as the main reservoir, and exhibits low pathogenicity (Lloret et al., Reference Lloret, Addie, Boucraut-Baralon, Egberink, Frymus, Gruffydd-Jones, Hartmann, Horzinek, Hosie, Lutz, Marsilio, Pennisi, Radford, Thiry, Truyen and Möstl2015; Veronesi et al., Reference Veronesi, Ravagnan, Cerquetella, Carli, Olivieri, Santoro, Pesaro, Berardi, Rossi, Ragni, Beraldo and Capelli2016).
In wild felids, infection is subclinical, whereas domestic cats develop an acute or hyperacute disease with a high mortality (about 100% of cases). However, in domestic cats, which serve as accidental dead-end hosts, the parasite, before erythrocyte invasion, undergoes an asexual phase of the life cycle within mononuclear phagocytes, producing large schizonts that occlude the microvasculature (Kocan et al., Reference Kocan, Kocan, Blouin and Mukolwe1992). The combined intra-monocytic and intra-erythrocytic proliferation results in extensive microvascular obstruction, ischaemic injury and multiorgan severe dysfunction, particularly affecting the liver, lungs, heart, kidneys and CNS. Clinical manifestations reflect the widespread microvascular ischaemia induced by schizonts within the monocytes and include severe lethargy, hyperthermia, anorexia, tachypnea or dyspnea, and icterus and hematological changes (i.e., hemolytic anaemia and thrombocytopenia) (Meinkoth et al., Reference Meinkoth, Kocan, Whitworth, Murphy, Fox and Woods2000). Neurological manifestations are secondary to cerebral microvascular occlusion by schizont-laden monocytes, leading to ischaemia, neuronal necrosis, microhaemorrhages and a neuroinflammatory response. They are characterized by depression, ataxia, tremors or seizures, correlating with ischaemic and inflammatory lesions of the CNS.
B. rossi (Order Piroplasmida, Family Babesiidae) is a highly pathogenic intra-erythrocytic protozoan parasite of domestic dogs, endemic to southern Africa and sporadically reported in other countries (Zygner et al., Reference Zygner, Gójska-Zygner, Bartosik, Górski, Karabowicz, Kotomski and Norbury2023). Transmission occurs through the blood-meal of hard ticks. Among canine Babesia spp., B. rossi exhibits the highest virulence, inducing severe intravascular and extravascular haemolysis (Schoeman, Reference Schoeman2009; Smith et al., Reference Smith, Goddard, Boddapati, Brooks, Schoeman, Lack, Leisewitz and Ackerman2021). Erythrocyte destruction is multifactorial, resulting from direct parasite-mediated membrane injury, increased osmotic fragility, oxidative damage and immune-mediated erythrocyte clearance (Smith et al., Reference Smith, Goddard, Boddapati, Brooks, Schoeman, Lack, Leisewitz and Ackerman2021).
Haemolytic anaemia accounts for the majority of the clinical manifestations of canine babesiosis, which may range from subclinical infection to acute, subacute or chronic disease courses (Zygner et al., Reference Zygner, Gójska-Zygner, Bartosik, Górski, Karabowicz, Kotomski and Norbury2023). The most frequently observed clinical signs include pyrexia, pallor, icteric mucous membranes, splenomegaly, thrombocytopenia, hyperbilirubinemia, bilirubinuria and haemoglobinuria. Although relatively uncommon, neurological complications have been documented in severe cases. They encompass seizures, ataxia, paresis, nystagmus, anisocoria, behavioural changes and involuntary paddling with vocalization. Such neurological signs can be frequently secondary to systemic disorders, including shock, tissue hypoxia, severe haemolysis, coagulopathy and microvascular endothelial injury rather than direct parasite invasion of the CNS. Neuroglycopenia secondary to hypoglycaemia has also been proposed as a contributing mechanism, as hypoglycaemia is a recognized complication of severe canine babesiosis and may itself produce neurological signs such as seizures, collapse or coma (Keller et al., Reference Keller, Jacobson, Nel, De Clerq, Thompson and Schoeman2004). Hyperacute cerebral babesiosis is characterized by microvascular sludging of parasitized erythrocytes, rapid endothelial necrosis and perivascular oedema, and is associated with a poor prognosis.
Further the neurological complications in course of B. rossi infection, it is also important to consider the potential for treatment-associated neurologic adverse effects in other canine babesioses for instance, Babesia canis infections. Even if B. canis infection is generally milder compared to B. rossi and the preferred treatment involves imidocarb dipropionate rather than diminazene aceturate, the latter is still occasionally employed in clinical practice (Han et al., Reference Han, Yoon and Hyun2014; Lee et al., Reference Lee, Yun and Song2025). Case reports indicate that diminazene administration, even at modest or standard doses, can induce acute multifocal encephalopathy, tetraparesis and seizures in dogs (Lee et al., Reference Lee, Yun and Song2025). These effects are generally reversible following cessation of the drug and supportive therapy, highlighting both the narrow therapeutic index of diminazene and the need for caution when using it, particularly in animals with pre-existing neurological compromise.
Arthropods affecting the CNS and/or PNS
Arthropod-borne neurotoxicoses and insect larval migrations represent essential, but often under-recognized, contributors to acute neurological dysfunction in dogs and cats (Rutland et al., Reference Rutland, Byl, Hydeskov, Miniter and Johnson2017; Deng et al., Reference Deng, Fu, Elsheikha, Cao, Zhu, Wang, Zhang, Xie, Yao and Liu2024).
Tick paralysis in small companion animals is caused by salivary neurotoxins produced by certain ixodid and argasid tick species. In Australia, Ixodes holocyclus is the principal cause of severe paralysis in dogs and cats. In cats, delayed treatment leads to a particularly high fatality rate (Padula, Reference Padula2016; Leister et al., Reference Leister, Morton, Atwell and Webster2018). In North America, most confirmed cases occur in dogs infested with Dermacentor andersoni or Dermacentor variabilis (Morshed et al., Reference Morshed, Li, Lee, Fernando, Lo and Wong2017). Feline paralysis is less common, although recent reports have described cases presumably associated with Ixodes spp. or Otobius megnini (Honnas et al., Reference Honnas, Athey, Verocai, Snowden, Esteve-Gasent and Mankin2020; Daniel et al., Reference Daniel, Ramos, Luksovsky, Galindo, Saleh and Verocai2024). Reports from Europe, Asia and Africa are less frequent, but they also indicate that local tick species can induce neuromuscular dysfunction (Deng et al., Reference Deng, Fu, Elsheikha, Cao, Zhu, Wang, Zhang, Xie, Yao and Liu2024). Regional variation in tick ecology and host exposure strongly influences disease incidence and severity, highlighting the importance of locality-specific awareness, treatment and prevention strategies (Padula et al., Reference Padula, Leister and Webster2020; Roeber and Webster, Reference Roeber and Webster2021); for instance, in Australia, the administration of tick antiserum currently remains the only specific therapy available for I. holocyclus paralysis (Padula et al., Reference Padula, Leister and Webster2020; Roeber and Webster, Reference Roeber and Webster2021). The neurotoxins involved interfere with acetylcholine release at the neuromuscular junction, leading to rapidly progressive, ascending flaccid paralysis (Edlow and McGillicuddy, Reference Edlow and McGillicuddy2008). If left untreated, affected animals may develop impaired respiratory mechanics, which can progress to respiratory failure and death (Teo et al., Reference Teo, Arganda, Webster, Yappa, Barker and Barker2023). Clinical signs typically include weakness, ataxia, voice changes and increased respiratory effort. Acute cases may deteriorate rapidly within 12 h to 5 days, with initial limb paresis advancing to hindquarter or generalized paralysis and respiratory depression. Because severe cases can be fatal even with treatment, early intervention is essential (Edlow and McGillicuddy, Reference Edlow and McGillicuddy2008; Teo et al., Reference Teo, Arganda, Webster, Yappa, Barker and Barker2023).
Cuterebra spp. (Order Diptera, Family Cuterebridae) are North American rodent and lagomorph botflies that can accidentally infect cats and dogs, which explore or hunt near rodent and rabbit burrows, where female Cuterebra flies typically lay their eggs. Most infections occur during summer months (July–September), consistent with peak Cuterebra activity and were associated with outdoor exposure (Rutland et al., Reference Rutland, Byl, Hydeskov, Miniter and Johnson2017). Larvae typically develop subcutaneously, but aberrant larval migration may allow larvae to enter the cranial cavity, causing an acute, focal forebrain disease characterized by inflammation, necrosis and severe vasospasm induced by larval toxins (Williams et al., Reference Williams, Summers and de Lahunta1998; Rutland et al., Reference Rutland, Byl, Hydeskov, Miniter and Johnson2017; Park et al., Reference Park, Sobotyk, Edwards, Porter and Verocai2021). In cats, cuterebral migration is suspected to cause feline ischaemic encephalopathy (Glass et al., Reference Glass, Cornetta, de Lahunta, Center and Kent1998; Rutland et al., Reference Rutland, Byl, Hydeskov, Miniter and Johnson2017). Clinically, acute-onset neurological signs such as behaviour change, circling, unilateral or bilateral central blindness, head pressing, vestibular deficit or seizures are described (Glass et al., Reference Glass, Cornetta, de Lahunta, Center and Kent1998; James and Poma, Reference James and Poma2010; Rutland et al., Reference Rutland, Byl, Hydeskov, Miniter and Johnson2017; Park et al., Reference Park, Sobotyk, Edwards, Porter and Verocai2021), and prognosis is guarded due to the irreversible CNS damage caused by larval migration.
Fungi affecting the CNS and/or PNS
Fungal diseases of the CNS are relatively rare in small animals, apart from the more commonly observed cases of cryptococcosis. Generally, they occur in immunocompromised animals, and these infections often pose significant diagnostic and treatment difficulties. In dogs and cats, CNS mycoses may present with either rapid onset or more slowly progressive clinical signs. Neurological abnormalities can appear localized to 1 region or, more commonly, involve multiple areas of the nervous system.
Cryptococcus spp. (Phylum Basidiomycota) are encapsulated yeasts with a worldwide distribution that can cause infections in both humans and a wide range of animal species. Currently, the pathogenic members of this genus are classified within the Cr. neoformans and Cr. gattii species complexes, which comprise distinct serotypes and molecular types (Hagen et al., Reference Hagen, Khayhan, Theelen, Kolecka, Polacheck, Sionov, Falk, Parnmen, Lumbsch and Boekhout2015). While Cr. neoformans complex has a cosmopolitan distribution and is typically associated with pigeon droppings, Cr. gattii complex is more often linked to eucalyptus trees in warmer climates. However, its range has expanded into temperate regions (Oliveira et al., Reference Oliveira, Flores, Mazzanti, Fighera and Kommers2024). Among domestic animals, cats are the most frequently affected, and cryptococcosis is their most common systemic mycotic disease.
A defining virulence trait of Cryptococcus spp. is the presence of a thick mucopolysaccharide capsule, which impairs phagocytosis, restricts leukocyte migration, modulates complement activation and suppresses T-cell-mediated immune responses (Buchanan and Murphy, Reference Buchanan and Murphy1998; Bovers et al., Reference Bovers, Hagen and Boekhout2008). Additional virulence determinants include thermotolerance and the production of enzymes such as urease, laccase and phospholipase. Moreover, the pronounced neurotropism of Cryptococcus spp. has been attributed to phenoloxidase-mediated melanin synthesis, which confers protection against oxidative stress and facilitates survival within catecholamine-rich neural tissues (Baker et al., Reference Baker, Chrissian, Stark and Casadevall2022). The Cr. neoformans complex predominantly affects immunocompromised hosts, whereas the Cr. gattii species complex is more frequently associated with disease in immunocompetent individuals (Stott et al., Reference Stott, Loyse, Jarvis, Alufandika, Harrison, Mwandumba, Day, Lalloo, Bicanic, Perfect and Hope2021; Galanis et al., Reference Galanis, MacDougall, Rose, Chen, Oltean, Cieslak, DeBess, Chong, Sorrell, Baddley, Hoang, Lockhart, Pappas and Phillips2025).
Infection usually begins with the inhalation of airborne basidiospores present in the environment. The pathogen typically establishes itself in the upper respiratory tract and lungs, and may subsequently disseminate via the bloodstream to the CNS, eyes, skin and other tissues. Yeasts may also access the brain through direct extension of nasal lesions, either across the cribriform plate or along the optic nerves (Malik et al., Reference Malik, Krockenberger, O’Brien, Martin, Wigney, Medleau and Greene2006). Neurological involvement occurs more frequently in dogs than in cats affected by cryptococcosis, and clinical signs often reflect multifocal disease, as the infection may affect virtually any level of the neuroaxis. Reported neurological signs might include (i) cortical and brainstem manifestations, such as blindness, seizures, behavioural or mentation changes and circling; (ii) cerebellar or vestibular abnormalities, such as ataxia and tremors; (iii) cranial nerve deficits, leading to paresis, altered pupil size or impaired facial sensation; and (iv) optic fundus abnormalities such as viz chorioretinitis, optic disc changes consistent with optic neuritis and retinal haemorrhage (Pennisi et al., Reference Pennisi, Hartmann, Lloret, Ferrer, Addie, Belák, Boucraut-Baralon, Egberink, Frymus, Gruffydd-Jones, Hosie, Lutz, Marsilio, Möstl, Radford, Thiry, Truyen and Horzinek2013; Jacobson et al., Reference Jacobson, Morton, Woerde, Thompson, Spillane, Tebb, Della Valle, Birnie and Malik2022; Oliveira et al., Reference Oliveira, Flores, Mazzanti, Fighera and Kommers2024). Hyperaesthesia and pain are also common, reflecting meningeal irritation, parenchymal inflammation or nerve root involvement. Dogs more frequently exhibit marked cervical pain, consistent with spinal or meningeal involvement. In contrast, cats tend to show more generalized discomfort or hindlimb pain, which may correspond to lumbosacral or peripheral nerve lesions (Sykes et al., Reference Sykes, Sturges, Cannon, Gericota, Higgins, Trivedi, Dickinson, Vernau, Meyer and Wisner2010).
Other than cryptococcosis, aspergillosis represents a common, cosmopolitan, opportunistic mycosis of cats and dogs, with a marked predilection for the sinonasal cavity and adjacent tissues. Aspergillus fumigatus is the species most frequently implicated, typically causing sinonasal disease; however, in its disseminated form, it may invade the CNS via haematogenous dissemination or by direct extension across the cribriform plate (Giordano et al., Reference Giordano, Gianella, Bo, Vercelli, Giudice, Della Santa, Tortorano, Peruccio and Peano2010; Cox et al., Reference Cox, Cattaneo, Russell, McGregor and Kortum2025). In cases of disseminated aspergillosis, Aspergillus terreus is more commonly identified than As. fumigatus and is associated with a condition in dogs characterized by poor clinical cure rates, frequent relapse and high case fatality (Schultz et al., Reference Schultz, Johnson, Wisner, Brown, Byrne and Sykes2008; Taylor et al., Reference Taylor, Young, Levine, Eden, Corapi, Rossmeisl and Levine2015; Lim et al., Reference Lim, Mansfield, Stevenson, Thompson, Davies, Whitney, James, Tebb, Fry, Buob, Hambrook, Boo and Dandrieux2022). More than half of reported cases occur in German Shepherd Dogs, a predisposition thought to reflect breed-related immunodeficiencies (e.g., IgA dysfunction) (Whitbread et al., Reference Whitbread, Batt and Garthwaite1984; Olsson et al., Reference Olsson, Tengvall, Frankowiack, Kierczak, Bergvall, Axelsson, Tintle, Marti, Roosje, Leeb, Hedhammar, Hammarström and Lindblad-Toh2015). Clinically, disseminated aspergillosis most often presents as discospondylitis, osteomyelitis, lymphadenopathy, nephritis, splenitis and meningoencephalitis (Schultz et al., Reference Schultz, Johnson, Wisner, Brown, Byrne and Sykes2008; Taylor et al., Reference Taylor, Young, Levine, Eden, Corapi, Rossmeisl and Levine2015; Lim et al., Reference Lim, Mansfield, Stevenson, Thompson, Davies, Whitney, James, Tebb, Fry, Buob, Hambrook, Boo and Dandrieux2022).
Cladophialophora species are pigmented fungi that can cause phaeohyphomycosis, a rare but serious infection in dogs and cats. When these fungi involve the CNS, they often produce granulomatous or necrotizing meningoencephalitis (Bouljihad et al., Reference Bouljihad, Lindeman and Hayden2002; Russell et al., Reference Russell, Gunew, Dennis and Halliday2016; Alonso et al., Reference Alonso, Fenton, Muller, Freeman, Becker, Rolph, Abramo, Rawlins, Kitson, Kessel and Thrall2024).
Histoplasma capsulatum, Coccidioides immitis and Blastomyces dermatitidis are systemic dimorphic fungi that can cause significant disease in several hosts, including dogs and cats. H. capsulatum and Bl. dermatitidis are distributed worldwide (with Bl. dermatitidis predominantly endemic to North America, whereas Co. immitis is endemic to the arid and semi-arid regions of the Americas) (Lavely and Lipsitz, Reference Lavely and Lipsitz2005). Infection typically arises following inhalation of spores from contaminated soil, leading to disseminated disease that may extend to the CNS, resulting in meningitis or granulomatous encephalitis. In all 3 infections, CNS involvement is uncommon but carries a poor prognosis, often manifesting as multifocal neurological deficits (Lavely and Lipsitz, Reference Lavely and Lipsitz2005; Reginato et al., Reference Reginato, Giannuzzi, Ricciardi, De Simone, Sanguinetti, Porcellato and Mandara2014).
E. cuniculi (Phylum Microsporidia, Family Encephalitozoonidae) is an obligate intracellular, spore-forming parasite closely related to fungi. It is characterized by its small size (1–2.5 µm), a resilient chitinous spore wall, and the presence of a distinctive invasion organelle, the polar filament, which is used to inject the infective sporoplasm into host cells (Brosson et al., Reference Brosson, Kuhn, Prensier, Vivarès and Texier2005). Although rabbits are recognized as the natural host, E. cuniculi infects a wide range of mammals, including dogs, cats, wild canids, rodents and humans (Hinney et al., Reference Hinney, Sak, Joachim and Kváč2016).
The life cycle consists of alternative proliferative and sporogonic phases within the cytoplasm of host cells. Mature spores are excreted in the urine and, possibly, in the faeces, thereby contaminating the environment (Brosson et al., Reference Brosson, Kuhn, Prensier, Vivarès and Texier2005). The principal route of infection is ingestion of contaminated food or water, although inhalation of spores and vertical transmission have also been reported (Hinney et al., Reference Hinney, Sak, Joachim and Kváč2016). In dogs, clinical disease due to E. cuniculi is most often linked to vertical transmission, whereas postnatal exposure usually results in subclinical infection; conversely, in cats, the occurrence of transplacental transmission remains debated (Santaniello et al., Reference Santaniello, Cimmino, Dipineto, Agognon, Beguinot, Formisano, Fioretti, Menna and Oriente2021). Following ingestion or vertical transmission, the parasite could disseminate haematogenously to multiple organs, particularly the kidneys, brain and eyes, causing chronic granulomatous inflammation due to parasite replication and immune-mediated tissue damage (Brosson et al., Reference Brosson, Kuhn, Prensier, Vivarès and Texier2005).
Clinical encephalitozoonosis in dogs is rare and occurs predominantly in puppies and immunocompromised animals, typically manifesting as the encephalitis–nephritis syndrome. Neurological signs may include head tremors, ataxia, seizures, tremors, circling, altered mentation, blindness and paresis (Snowden et al., Reference Snowden, Lewis, Hoffman and Mansell2009; Postma et al., Reference Postma, Pardini, Carnevale, Gregnoli, Quiroga, Venturini and Minatel2018; De Boer et al., Reference De Boer, Diaz Espineira and Mandigers2021). Neurological involvement is frequently accompanied by evidence of renal dysfunction (i.e., polyuria, polydipsia, weight loss and anorexia) and occasionally ocular lesions (Snowden et al., Reference Snowden, Lewis, Hoffman and Mansell2009; Hofmann-Wellenhof and Nell, Reference Hofmann-Wellenhof and Nell2025).
Encephalitozoonosis in cats is rare and often limited to the ocular form (i.e., bilateral cataracts and anterior uveitis) (Benz et al., Reference Benz, Maass, Csokai, Fuchs-Baumgartinger, Schwendenwein, Tichy and Nell2011; Hofmann-Wellenhof and Nell, Reference Hofmann-Wellenhof and Nell2025). Epidemiological surveys in Austria and the United States found no significant association between E. cuniculi seropositivity and feline meningoencephalitis or chronic kidney disease, indicating that systemic forms are infrequent (Hsu et al., Reference Hsu, Grant, Zajac, Witonsky and Lindsay2011; Kunzel et al., Reference Kunzel, Rebel-Bauder, Kassl, Leschnik and Url2017). A single report of generalized encephalitozoonosis with cerebellar hypoplasia was reported in a kitten (Rebel-Bauder et al., Reference Rebel-Bauder, Leschnik, Maderner and Url2011).
Diagnostic tools in parasitic diseases of the CNS and/or PNS
The diagnosis of parasitic infections affecting the CNS and PNS of dogs and cats is challenging due to the broad range of neurotropic pathogens involved, the overlapping clinical signs and the complexity of pathogenic mechanisms. To achieve an accurate diagnosis, which is essential for timely and appropriate therapeutic intervention, a comprehensive, multidisciplinary diagnostic approach integrating serological, antigenic, molecular and morphological methodologies (including histology and CSF cytology) is required. Selection of the most appropriate diagnostic tool depends on the clinical presentation, onset and duration of signs, suspected etiological agent, sample availability and the level of specificity and sensitivity required for informed clinical decision-making.
Immunoassays are widely employed as first-line screening tools, allowing the detection of parasite-specific antibodies (IgG, IgM) or circulating antigens in a variety of biological matrices (Lappin, Reference Lappin2009). Interpretation of immunological results, however, must carefully account for parasite biology and the kinetics of the host immune response. Molecular diagnostic techniques, predominantly Polymerase Chain Reaction (PCR)-based methods, provide high analytical sensitivity and specificity by directly detecting parasite DNA or RNA in CSF (intra vitam) or tissue samples (post-mortem). These methods are particularly valuable when serological findings are inconclusive or when distinguishing closely related species is required (Lappin, Reference Lappin2009). Morphological techniques, including histopathology, immunohistochemistry, special staining methods and cytological examination, enable direct visualization of parasites and/or characteristic lesions and constitute the definitive diagnostic approach in many cases.
Antigen and/or antibody diagnostics
Serological diagnosis of parasitic infections affecting the CNS and PNS of dogs and cats relies on the detection of parasite-specific antibodies and/or circulating antigens in different biological matrices. Assessment of the host systemic humoral immune response is typically performed on serum or plasma. It is particularly relevant for immunogenic and systemically distributed parasites such as T. gondii, N. caninum, E. cuniculi, L. infantum and D. immitis (in cats) (Dubey and Lindsay, Reference Dubey and Lindsay1996; Simón et al., Reference Simón, Kramer, Román, Blasini, Morchón, Marcos-Atxutegi, Grandi and Genchi2007; Dubey, Reference Dubey2010; Deshmukh et al., Reference Deshmukh, Gurupwar, Mitra, Aswale, Shinde and Chaudhari2021; Priolo et al., Reference Priolo, Martínez-Orellana, Pennisi, Raya-Bermúdez, Jurado-Tarifa, Masucci, Donato, Bruno, Castelli and Solano-Gallego2022).
Nevertheless, important limitations may exist in the interpretation of antibody-based assays, as multiple factors must be taken into account, including the timing of infection, potential serological cross-reactivity (e.g., within the Apicomplexa and with Aspergillus spp.), parasite biology and interference from maternally derived passive immunity during the first months of life (Bertlich et al., Reference Bertlich, Freytag, Huber, Dombrowski, Oppel and Gröger2024; Mendoza et al., Reference Mendoza, Pérez-Écija, Kappmeyer, Suarez and Bastos2024). Indeed, chronic or persistent infections, such as those caused by T. gondii, N. caninum and E. cuniculi, may elicit low humoral response due to reduced antigenic stimulation, immune-evasion strategies (e.g., development of minimally immunogenic stages such as tissue cysts) or localization within poorly immunogenic CNS compartments (Dubey and Lindsay, Reference Dubey and Lindsay1996; Dubey, Reference Dubey2010; Hofmann-Wellenhof and Nell, Reference Hofmann-Wellenhof and Nell2025).
CSF or aqueous humor can serve as additional diagnostic matrices for assessing in situ antibody production, particularly useful in organ-active infection or reactivation (e.g., T. gondii and N. caninum). However, their use is limited by technical constraints, such as the difficulty of CSF collection in small puppies or cats. Moreover, this approach requires consideration of the fact that intrathecal immunoglobulin synthesis may coexist with passive diffusion of immunoglobulins from the systemic circulation across the BBB. Consequently, antibody concentrations in CSF do not mirror peripheral immunokinetic patterns and cannot be reliably interpreted without accounting for blood–CSF barrier permeability (Tipold et al., Reference Tipold, Pfister, Zurbriggen and Vandevelde1994; Thomas, Reference Thomas1998; Reiber and Peter, Reference Reiber and Peter2001).
Dedicated quantitative approaches are therefore required to discriminate true intrathecal antibody production from passive transfer. Calculation of the specific antibody quotient (QIgSpec), defined as the ratio between parasite-specific antibodies measured in CSF (LAbSpec) and serum (SAbSpec), provides an initial estimate of antibody distribution between compartments. To determine whether CSF antibody levels exceed those expected from passive diffusion alone, the antibody index (AI) is calculated by normalizing QIgSpec to the total immunoglobulin quotient (QIgTot). AI values exceeding established physiological thresholds (typically >1·5) are indicative of active intrathecal antibody synthesis (Thomas, Reference Thomas1998; Reiber and Peter, Reference Reiber and Peter2001). Given that CSF immunoglobulin concentrations are generally low, highly sensitive analytical methods are required, most commonly adapted from serum-standardized enzyme-immunosorbent antibody assay (ELISA)-based techniques.
A broad spectrum of commercially available serological assays is employed for antibody detection (Table 2) in dogs and cats and can be categorized into qualitative and quantitative methods, each showing distinct diagnostic applications and accuracy. Qualitative assays, including immunochromatographic tests and rapid-ELISA formats such as SNAP tests (Idexx Laboratories, USA), that provide rapid results in a few minutes, are widely adopted in clinical practice due to their ease of use, minimal sample processing and short turnaround time. They provide binary presence/absence results, without distinguishing immunoglobulin subclasses (i.e., IgG versus IgM). Moreover, they are often not validated for matrices beyond serum or plasma (Velayudhan and Naikare, Reference Velayudhan and Naikare2022). In contrast, quantitative assays, such as indirect immunofluorescence antibody tests (IFAT), ELISA and direct agglutination test, allow a precise measurement of antibody titers, which correlate to the antigenic stimulation intensity, and can be very useful for clinical framing, monitoring of disease progression and assessment of therapeutic response in the medium-term period (Solano-Gallego et al., Reference Solano-Gallego, Miró, Koutinas, Cardoso, Pennisi, Ferrer, Bourdeau, Oliva and Baneth2011; Morganti et al., Reference Morganti, Rigamonti, Brustenga, Calgaro, Angeli, Moretta, Diaferia and Veronesi2024a).
Antibody- and antigen-based assays commonly used for the diagnosis of major parasitic infections that may affect the nervous system of small companion animals

Table 2 Long description
The table lists parasitic agents affecting small companion animals and summarizes, for each, the antibody-detection method, antigen-detection method, clinical sample type, and supporting references. Serum is the most common sample across agents, with cerebrospinal fluid also used for Toxoplasma gondii, Neospora caninum, Encephalitozoon cuniculi, Cryptococcus neoformans, and Aspergillus spp.; urine is additionally listed for E. cuniculi, and faeces for Toxocara canis. Antibody detection most often relies on ELISA IgG or serologic methods such as IFAT and DAT, with rapid tests noted for several agents (e.g., T. gondii, N. caninum, Leishmania infantum). Antigen detection is reported for fewer organisms: ELISA or rapid antigen tests are used for Angiostrongylus vasorum and Dirofilaria immitis, ELISA is listed for T. canis and E. cuniculi, and Cryptococcus uses ELISA or lateral flow assays. Some agents have no antigen test listed (e.g., Aelurostrongylus abstrusus, T. gondii, N. caninum, L. infantum), and Cryptococcus has no antibody test listed. Aspergillus antigen testing includes enzyme immunoassay and rapid chemiluminescence formats marked as not commercially available or not validated for the target animal species.
NA, not commercially available; CFS: cerebrospinal fluid; IFAT: immunofluoresce antibody test; ELISA: enzyme-linked immunosorbent assay; DAT, direct agglutination test; LFA, lateral flow assay; EIA, enzyme-immuno assay; CLIA, rapid chemiluminescence immunoassay.
a Commercially available but validated in human species.
Complementary or in addition to antibody detection, antigen assays targeting circulating parasitic components in serum, plasma, CSF, aqueous humor or urine enhance diagnostic sensitivity, particularly for parasites that induce low immunogenicity in early stages of infection or impaired antibody production (Table 2).
Antigen detection has proven to be valuable, particularly for pathogens, such as A. vasorum, D. immitis (in dogs) and Cr. neoformans, where active infections are associated with measurable circulating antigens (Velayudhan and Naikare, Reference Velayudhan and Naikare2022). Commercially available assays for parasitic antigen detection are predominantly based on immunoassay platforms originally developed and validated on serum, plasma or whole blood; several of them can also be applied to CSF or urine when supported by adequate analytical sensitivity and appropriate validation. The majority of routinely used tests consisted of ELISA or immunochromatographic formats, including rapid in-clinic SNAP® assays, that employ monoclonal antibodies targeting circulating parasite-derived antigens and allow a rapid and specific detection even in cases where humoral responses are weak, delayed or absent. ELISA-based antigen tests offer high analytical sensitivity and quantitative or semi-quantitative outputs, making them suitable for laboratory settings and, in some cases (e.g., A. vasorum infection), for monitoring infection dynamics over time (Velayudhan and Naikare, Reference Velayudhan and Naikare2022).
Molecular diagnostics
Molecular techniques have become an indispensable component of the diagnostic workup for parasitic infections affecting the CNS and PNS of dogs and cats. These methods should be integrated with serological and antigen-based assays in cases characterized by low parasitic burden, early phase of infection or compartmentalized disease, and they allow accurate species discrimination and monitoring of infection dynamics over time, which is crucial for prognosis assessment and therapeutic planning.
In general, quantitative real-time PCR (qPCR), nested PCR and isothermal amplification techniques, such as loop-mediated isothermal amplification (LAMP) or recombinase polymerase amplification, represent the main molecular approaches applied in clinical and research settings (Table 3). The selection of the most appropriate technique depends on the biological characteristics, the type and quality of the clinical sample and the required diagnostic performance.
Molecular targets and corresponding techniques commonly employed for the diagnosis of major parasitic infections that may affect the nervous system of small companion animals

Table 3 Long description
The table links each causative agent to the clinical sample type, the molecular target used for detection, and the biomolecular technique reported in the cited studies. qPCR is the predominant method across agents, applied to blood (e.g., Angiostrongylus vasorum, Cytauxzoon felis, Babesia rossi), CSF (e.g., Angiostrongylus cantonensis, Toxoplasma gondii, Neospora caninum, Cryptococcus neoformans), and necropsied brain tissue including fresh or FFPE material (e.g., Baylisascaris procyonis, T. gondii, Neospora caninum, Leishmania infantum, Cryptococcus neoformans). Molecular targets vary by organism: ITS regions are used for A. vasorum, C. felis, B. rossi and Encephalitozoon cuniculi; mitochondrial cox genes are used for A. cantonensis (cox1) and B. procyonis (cox2); B. rossi uses 18S rRNA ; T. gondii uses the 529-REP element; N. caninum uses the NC5 region; and C. neoformans uses multicopy rDNA. Different methods appear for some agents, including LAMP for A. cantonensis, T. gondii, and N. caninum; RPA for B. rossi; and conventional or nested PCR for E. cuniculi. Several pathogens list multiple acceptable sample types (notably CSF plus brain tissue), indicating flexibility depending on clinical versus post-mortem availability. Reference entries include multiple citations per organism.
CSF, cerebrospinal fluid; FFPE, formalin-fixed paraffin-Embedded; BAL, bronco-alveolar lavage; LAMP, loop-mediated isothermal amplification; rDNA, ribosomal DNA; RPA, recombinase polymerase amplification.
Multiplex qPCR platforms, often targeting multicopy ribosomal DNA sequences, can be employed for the simultaneous detection of multiple pathogens (e.g., T. gondii, N. caninum, Cr. neoformans) in a single analytical reaction (Han et al., Reference Han, Chang and Na2015). Typical clinical samples include CSF, brain tissue (fresh necropsy tissue or formalin-fixed paraffin-embedded tissue), blood, urine or bronchoalveolar lavage (BAL).
For protozoa such as T. gondii and N. caninum, multiplex qPCR assays targeting the 529-REP element and the NC5 region, respectively, enable highly sensitive detection from CSF or brain tissue, even when parasite burdens are low (Truong and Šlapeta, Reference Truong and Šlapeta2023). Haemoparasites, such as Cy. felis and B. rossi, are generally identified by qPCR assays targeting the ITS or 18S rRNA regions from blood samples (Lloret et al., Reference Lloret, Addie, Boucraut-Baralon, Egberink, Frymus, Gruffydd-Jones, Hartmann, Horzinek, Hosie, Lutz, Marsilio, Pennisi, Radford, Thiry, Truyen and Möstl2015; Zygner et al., Reference Zygner, Gójska-Zygner, Bartosik, Górski, Karabowicz, Kotomski and Norbury2023). In contrast, microsporidia, such as E. cuniculi, are commonly detected using nested or conventional PCR targeting the rRNA ITS region in blood, urine or tissue samples (Santaniello et al., Reference Santaniello, Cimmino, Dipineto, Agognon, Beguinot, Formisano, Fioretti, Menna and Oriente2021). Nematodes with neuroinvasive potential, such as A. cantonensis and A. vasorum, are detected via qPCR, targeting mitochondrial or ribosomal gene regions in CSF, blood or BAL, with LAMP providing potential point-of-care application (Jefferies et al., Reference Jefferies, Morgan, Helm, Robinson and Shaw2011; Mahalingam et al., Reference Mallaiyaraj Mahalingam, Calvani, Lee, Malik and Šlapeta2021; Baláž et al., Reference Baláž, Rivory, Hayward, Jaensch, Malik, Lee, Modrý and Šlapeta2023). More recently, highly sensitive assays such as the AcanR3990 qPCR have been developed for the detection of A. cantonensis, improving diagnostic sensitivity compared with earlier ITS1-based methods (Lee et al., Reference Lee, Pai, Churcher, Davies, Braddock, Linton, Yu, Bell, Wimpole, Dengate, Collins, Brown, Reppas, Jaensch, Wun, Martin, Sears, Šlapeta and Malik2020; Sears et al., Reference Sears, Qvarnstrom, Dahlstrom, Snook, Kaluna, Baláž, Feckova, Šlapeta, Modry, Jarvi and Nutman2021). L. infantum can be detected in CSF or brain tissue via qPCR targeting kDNA or genomic sequences, allowing assessment of CNS involvement and coinfections with other pathogens (Solano-Gallego et al., Reference Solano-Gallego, Miró, Koutinas, Cardoso, Pennisi, Ferrer, Bourdeau, Oliva and Baneth2011).
CSF cytology
CSF analysis represents a valuable diagnostic tool in the diagnosis of parasitic inflammatory diseases affecting the CNS (Table 4). A marked increase in total nucleated cell count is typically observed when inflammatory lesions involve or border the ventricular system or the subarachnoid space. Therefore, pleocytosis (>5–8/mL in the cisterna magna; >2–5/mL in the lumbar cistern) is commonly detected, with a predominance of monocytes, lymphocytes or polymorphonucleated cells, depending on the specific inflammatory pattern expressed by the causative parasitic (Bernardi et al., Reference Bernardi, Brunati, Biagioli, Buoro, Cataldo, Ciusani, Corsini, Dessì, Fanelli, Muratore, Passerini and Previtarli2014; De Lorenzi and Mandara, Reference De Lorenzi, Mandara, Raskin and Meyer2016). More commonly, parasite inflammation of the CNS is associated with lymphocytic and monocytic pleocytosis, with polymorphonuclear cells being less frequently observed. In fact, contrary to expectations, parasitic infections are only rarely associated with marked eosinophilic pleocytosis. This is observed in certain helmintic migrations, such as those caused by A. cantonensis, S. lupi and Baylascaris procyonis (Wise et al., Reference Wise, Sorvillo, Shafir, Ash and Berlin2005; Wang et al., Reference Wang, Lai, Zhu, Chen and Lun2008; Chai et al., Reference Chai, Yas, Brenner, Rojas, Konstantin, Klainbart and Shamir2018). Neverthless, it cannot be excluded that the use of antihelminthic drugs, by inducing larval death, may increase parasite-derived eosinophilic inflammation. This may occur due to the sudden release of metazoan antigens and cessation of parasite-derived immunosuppressive molecule production (Lunn et al., Reference Lunn, Lee, Smaller, MacKay, King, Hunt, Martin, Krockenberger, Spielman and Malik2012). Although parasitic diseases should be included in the differential diagnosis when eosinophilic pleocytosis is present in the CSF, idiopathic eosinophilic meningoencephalitis should also be considered. As for neutrophilic pleocytosis, it is characteristic of dogs affected by aspergillosis (Taylor et al., Reference Taylor, Young, Levine, Eden, Corapi, Rossmeisl and Levine2015).
Prevalent localization, main histopathological patterns and CSF cell prevalence associated with parasitic infections affecting the nervous system of small companion animals

Table 4 Long description
The table links specific parasitic agents affecting the nervous system of small companion animals with their most common CNS localization, dominant histopathology, typical CSF cell pattern, and supporting references. Brain involvement is frequent and includes haemorrhages with Angiostrongylus vasorum (monocytic CSF) and Angiostrongylus cantonensis (eosinophilic CSF), eosinophilic abscesses with Baylisascaris procyonis (eosinophilic CSF), and necrotizing or granulomatous inflammation with agents such as Toxoplasma gondii (lymphocytic CSF) and Encephalitozoon cuniculi (monocytic CSF). Spinal cord–predominant entries include Spirocerca lupi (eosinophilic meningomyelitis with eosinophilic CSF) and Gurltia paralysans (suppurative meningitis with neutrophilic CSF, and also non-suppurative meningitis with lymphocytic CSF). Several agents are associated with non-suppurative inflammation and lymphocyte-rich CSF, including Taenia serialis (coenurosis) and Cytauxzoon felis (brain and spinal cord). Lymphomonocytic CSF patterns are reported with Capillaria boehmi, Dirofilaria immitis, Neospora caninum (spinal nerves/cerebellum with polyradiculoneuritis and necrotizing cerebellitis), and Leishmania infantum (leptomeningitis, including granulomatous patterns. Neutrophilic CSF is highlighted with suppurative processes such as Cuterebra spp. (suppurative meningitis and territorial infarcts). Some rows list multiple localizations or multiple histologic patterns for the same agent, so the table summarizes prevalent associations rather than exclusive findings.
MP, monocytic pleocytosis; EP, eosinophilic pleocytosis; LP, lymphocytic pleocytosis; NP, neutrophilic pleocytosis; LMP, lymphomonocytic pleocytosis.
Similarly, a specific increase in CSF protein concentration (>25 mg/dl in the cisterna magna; >45 mg/dl in the lumbar cistern) can be found and is generally associated with nervous tissue damage or the presence of parasitic organisms. Moreover, ELISA or IFAT, can be applied to detect specific-antibodies, supporting evidence of intrathecal antiparasitic immune responses and, consequently, CNS involvement. Furthermore, PCR-based assays performed on CSF samples are recommended to directly identify the presence of parasite DNA in the CNS. Finally, cytological examination of CSF may allow direct visualization of parasitic organisms in the CNS.
Diagnostic histopathology
Helminths
Migration of helminths within the CNS is typically associated with granulomatous inflammatory reactions. However, different patterns of lesions can be observed in different infections. Young dogs are occasionally been reported to develop severe cerebral eosinophilic meningoencephalomyelitis, characterized by extensive necrosis and micro-abscesses formation, following accidental ingestion of Baylascaris procyonis eggs (Hazlett et al., Reference Hazlett, Cai, Sparling and You2018; Heller et al., Reference Heller, Arnold and Dreyfus2019). Similarly, eosinophilic meningoencephalomyelitis is reported for aberrant migration of A. cantonensis and S. lupi (Martins et al., Reference Martins, Tanowitz and Kazacos2015; Casoy et al., Reference Casoy, Srugo, Barnoon, Segal, Zafrany, Segev and Shipov2025). Otherwise, well-preserved larvae can be observed in the nervous tissue in the absence of a marked inflammatory response.
Neurological syndromes caused by A. vasorum are generally associated with severe intracerebral haemorrhages that replace normal brain tissue, resulting from parasite-induced severe hemostatic dysfunction. In cases where nematode larvae are present within the cerebral neuroparenchyma, foci of granulomatous inflammation may develop with macrophages, lymphocytes, plasma cells, eosinophils and multinucleated giant cells (Denk et al., Reference Denk, Matiasek, Just, Hermanns, Baiker, Herbach, Steinberg and Fischer2009) (Fig. 1C, D).
Toxoplasmosis. Dog (A and B). (A) Transverse brain section. The cerebral cortex shows multifocal areas of softening (arrows) with irregular cortical profile. (B) A tissue cyst rich in bradyzoites. Lymphocytes and plasma cells infiltrate the necrotic nervous tissue. Reactive hypertrophic astrocytes (gemistocytes) are recognizable (arrow) (H&E, 40×). Angiostrongylosis. Dog (C and D). (C) Transverse cerebellum section. Diffuse hemorrhages alongside the leptomeninges. (D) Reactive endothelial cells and gliosis associated with Angiostrogylus vasorum larvae (arrows) (H&E, ×20). Cryptococcosis. Cat (E and F). (E) Transverse brain section. A number of gelatinous pseudocysts and malacic cysts rich in yeasts are present in the parietal cortex and in the caudate nuclei, respectively. (F) Typical yeasts of Cryptococcus are scattered in lymphomonocytic and neutrophilic infiltration (H&E, 40×).

Figure 1 Long description
The image A showing a coronal brain section, with three white arrows pointing to different cerebral cortex areas of softening . The image B showing a H&E micrograph with leucocytic infiltration and an eosinophilic parasitic cyst ; one black arrow points to a gemistocytic cell; The image C showing a coronal cerebellar section with diffuse hemorrhages alongside the leptomeinges; The image D showing a H&E stained micrograph with reactive blood vessels and gliosis; ttwo black arrows point the parasitic larvae; The image E showing a coronal brain section with gelatinous pseudocysts and malacic cysts containing yeasts in the left parietal cortex and in both the caudate nuclei; The image F showing a H&E stained micrograph with a number of yeasts of Cryptococcus scattered in lymphomonocytic and neutrophilic infiltration.
On the other hand, A. cantonensis causes mechanical peripheral nerve injury and haemorrhages secondary to the retrograde migration process to the CNS. Thrombosis and aneurysms can also occur secondary to larval migration (Chotmongkol and Sawanyawisuth, Reference Chotmongkol and Sawanyawisuth2002). When it arrives in the brain or spinal cord, severe eosinophilic meningoencephalomyelitis is activated by larval antigens, such as metabolic, excretory or moulting products (Lunn et al., Reference Lunn, Lee, Smaller, MacKay, King, Hunt, Martin, Krockenberger, Spielman and Malik2012). Occasionally, a granulomatous reaction can be found around the nematode (Lunn et al., Reference Lunn, Lee, Smaller, MacKay, King, Hunt, Martin, Krockenberger, Spielman and Malik2012).
Aberrant migration of S. lupi tends to primarly affect the spinal cord along the extradural or intradural subarachnoid spaces. The severity of clinical signs and neuroanatomical localization may vary throughout disease progression, likely reflecting ongoing spinal cord damage associated with migration through the parenchyma (Casoy et al., Reference Casoy, Srugo, Barnoon, Segal, Zafrany, Segev and Shipov2025).
G. paralysans parasitize the meningeal vessels, producing severe congestion of the meningeal veins in the spinal cord, which progresses to perivenous fibrosis. Suppurative or lymphoplasma histiocytic meningitis associated with vasculitis of the meningeal vessels mainly affects the spinal cord (Hartung et al., Reference Hartung, Weyrich, Moroni, Gòmez and Harden2022). Axonal degeneration and gliosis, myelomalacia and inflammatory infiltrates have also been reported in the spinal cord, in proximity to parasites.
Aberrant localizations of adult Ae. abstrusus are an infrequent event. Fatal necrotic and haemorrhagic multifocal meningoencephalitis has been reported in a cat with gross lesions in the cerebral cortex, cerebellum and brain stem. Subarachnoid haemorrhages were also observed in the cervical and lumbar region of the spinal cord (Tinoco et al., Reference Tinoco, Morelli, de Farias Brito, Oliveira Pereira, Correia Oliveira, Diakou, Colombo, Frangipane Di Regalbono and Traversa2022).
Aberrant migration of C. boehmi from the nasal cavities in the CNS can be associated with an inflammatory response, characterized by lymphocytes, plasma cells and macrophages, with few neutrophils and eosinophils (Clark et al., Reference Clark, López, Levine, Cooper, Craig, Voges, Johnson and Porter2013). The inflammatory reaction may also extend to involve the meninges, resulting in the formation of mass-like lesions that mimic extra-axial masses.
Historically, aberrant migration of D. immitis in the CNS is reported in the canine and feline literature associated with encephalitis (Bozeman and Phillips, Reference Bozeman and Phillips1972). Multifocal foci of malacia can also be observed; it is supposed that they are caused by toxic metabolic products of the parasite or the physical pressure exerted by it (Molnar et al., Reference Molnar, Pazàr, Rigòt, Mhàté, Fok, Glàvits, Vajdovich, Jacsò, Balogh and Sòs2010).
Feline cerebral coenurosis caused by T. serialis is characterized by encephalitis with mixed-cellular inflammatory infiltrate, composed predominantly of lymphocytes, neutrophils and eosinophils (Jull et al., Reference Jull, Browne, Boufana, Schöniger and Davies2012). The histological recognition of metacestodes supports the diagnosis.
Protozoan
T. gondii and N. caninum produce very similar lesions in the CNS. Therefore, to specifically attribute the lesions to each of them requires an exact protozoan identification. The association between the lesions and the neurological syndrome needs specific identification of parasitic cysts or of tachyzoites widespread in the affected area (Fig. 1B). Immunohistochemistry is the diagnostic gold standard for this purpose.
Dogs congenitally infected by N. caninum can be severely affected during the first few months of life, presenting with myositis/polyradiculoneuritis (Cantile and Youssef, Reference Cantile and Youssef2016). Less commonly, young dogs develop encephalomyelitis (Mandara et al., Reference Mandara, Cantile, Baroni and Bernardini2010). Lesions in peripheral nerves or radicles are characterized by infiltration of lymphocytes and monocytes, occasionally accompanied by eosinophils and neutrophils, and are associated with secondary degeneration and necrosis in both muscles and nerves (Cantile and Arispici, Reference Cantile and Arispici2002).
In the CNS, which is predominantly affected in adult dogs, lesions are typified by perivascular lymphohistiocytic cuffing, gliosis and capillary proliferation, resulting in necrotizing and granulomatous multifocal meningoencephalomyelitis (Vandevelde et al., Reference Vandevelde, Higgins and Oevermann2012) (Fig. 1A). Sometimes, mass lesions simulating neoplasms may be observed (Falzone et al., Reference Falzone, Baroni, De Lorenzi and Mandara2008). In the brain, the most severe lesions generally involve the grey matter, whereas in the spinal cord, the white matter is predominantly affected (Mandara et al., Reference Mandara, Cantile, Baroni and Bernardini2010). In adult dogs with N. caninum infection, the cerebellum can be dramatically involved, with necrotizing cerebellitis leading to gross cerebellar cortex atrophy (Cantile and Arispici, Reference Cantile and Arispici2002). The association between these lesions and the observed neurological syndrome requires specific identification of parasitic cysts or widespread tachyzoites within the affected regions (Fig. 1B).
In neurological leishmaniosis, meningitis is the most common inflammatory pattern; however, choroiditis, encephalitis and myelitis have also been reported (Oliveira et al., Reference Oliveira, Boechat, Mendes Junior, Madeira, Ferreira, Figueiredo, Campos, de Carvalho Rodrigues, Carvalhaes de Oliveira, Amendoeira and Menezes2017). Inflammation can occur as a granulomatous non-suppurative pattern, a non-granulomatous (lymphoplasmacytic) infiltration or a granulomatous suppurative infiltration (Oliveira et al., Reference Oliveira, Boechat, Mendes Junior, Madeira, Ferreira, Figueiredo, Campos, de Carvalho Rodrigues, Carvalhaes de Oliveira, Amendoeira and Menezes2017). Leishmaniosis has also been found in association with epidural granulomas and macrophagic meningitis in the spinal cord (Cauduro et al., Reference Cauduro, Favole, Lorenzo, Simonetto, Barda, Cantile and Asperio2011). Occasionally, amastigote forms can be observed within the cytoplasm of macrophages located in the perivascular space (Oliveira et al., Reference Oliveira, Boechat, Mendes Junior, Madeira, Ferreira, Figueiredo, Campos, de Carvalho Rodrigues, Carvalhaes de Oliveira, Amendoeira and Menezes2017). Radiculoneuritis may also be present, reflecting peripheral nerve involvement (Márquez et al., Reference Márquez, Pedregosa, López, Marco-Salazar, Fondevila and Pumarola2013).
The most prominent lesions associated with CNS invasion by Cy. felis consist of intraluminal, schizont-laden macrophages within arterioles and venules of leptomeninges and nervous parenchyma. Occasionally, these schizont-laden macrophages can be observed outside blood vessels, within the surrounding neural tissue. In these regions, histopathological examination reveals edema, microgliosis, astrocytosis, microhaemorrhages and multiple foci of necrosis. The distribution of lesions corresponds to areas of vascular occlusion by protozoal organisms (Clarke and Rissi, Reference Clarke and Rissi2015).
Arthropods
CNS lesions associated with invasion by Cuterebra spp. larvae consist of suppurative meningitis foci caused by bacterial contamination, or areas of cerebral necrosis resulting from larval migration (Rissi and Howerth, Reference Rissi and Howerth2013). However, cuterebriasis is more commonly associated with a cerebrovascular syndrome known as feline ischaemic encephalopathy, mainly affecting the median cerebral artery (De Lahunta and Glass, Reference De Lahunta and Glass2009). Gross lesions typically include large territorial infarcts involving the parietotemporal cortex. The recovery of the insect larva consistent with Cuterebra sp. next to the lesions or within the calvarium confirms the cerebrospinal cuterebriasis.
Fungi
Mycotic infections of the CNS initially produce suppurative inflammatory responses. As the disease progresses, extensive necrotic lesions may develop, often associated with vasculitis or a pronounced granulomatous reaction, particularly in chronic infections. Cryptococcus spp. can produce multifocal or diffuse meningitis and submeningeal encephalitis, sometimes ependymitis, subependymal encephalitis and choroiditis (Sykes et al., Reference Sykes, Sturges, Cannon, Gericota, Higgins, Trivedi, Dickinson, Vernau, Meyer and Wisner2010).
When fungal organisms are present in large numbers, the meninges may become coated with abundant gelatinous proteinaceous material, leading to expansion of the Virchow–Robin spaces and the formation of gelatinous pseudocysts (Sykes et al., Reference Sykes, Sturges, Cannon, Gericota, Higgins, Trivedi, Dickinson, Vernau, Meyer and Wisner2010) (Fig. 1E, F). Occasionally, tumour-like intraparenchymal granulomas containing characteristic yeast cells with a prominent clear capsule are observed. These mass lesions, referred to as cryptococcomas, are more commonly reported in immunocompetent human patients (Mandrioli et al., Reference Mandrioli, Bettini, Marcata, Benazzi, Della Salda, Krockenberger and Jensen2002; Klock et al., Reference Klock, Cerski and Goldani2009). Periodic Acid-Schiff and Grocott stains are valuable tools for histological diagnosis of the infection.
E. cuniculi selectively parasitizes vascular endotheliocytes, leading to arteriolar fibrinoid necrosis and subsequent secondary lesions (Cantile and Youssef, Reference Cantile and Youssef2016). In the CNS of dogs, foxes and minks, as in rabbits, this microsporidium induces characteristic nodular microgranulomas composed of epithelioid histiocytes, forming the so-called ‘hepatoid’ foci that are widely distributed throughout the brain. The parasites can be identified in histological sections stained with Giemsa, Ziehl–Neelsen or Gram stains, appearing as oblong elements (Vandevelde et al., Reference Vandevelde, Higgins and Oevermann2012).
Conclusions
Parasitic infections affecting the nervous system of dogs and cats represent a clinically significant cause of severe neurological diseases. Parasitic involvement of the CNS and/or PNS in small companion animals closely parallels what is described in other hosts, including humans; in fact, the neurological manifestations observed are highly variable and rarely pathognomonic, making differentiation from viral, bacterial or immune-mediated encephalitis particularly challenging, and are often associated with substantial morbidity and a guarded prognosis.
The neuropathological patterns described in small companion animals also closely resemble those documented in other species affected by neurotropic parasites, underscoring shared mechanisms of neuroinvasion, persistence and immune-mediated tissue damage across taxonomically different organisms. The neurological consequences of the parasitic infections in dogs and cats, ranging from seizures and vestibulocerebellar dysfunctions to paresis, paralysis and behavioural alterations, mirror the substantial burden observed in human neuroparasitic diseases such as neurocysticercosis, cerebral toxoplasmosis and other neglected tropical infections (Shahrivar et al., Reference Shahrivar, Moghimi, Hosseinzadeh, Kohansal, Mortazavi, Galeh and Ahmadpour2025). Despite this, parasitic infections of the CNS and PNS in small companion animals remain comparatively underrepresented in research and surveillance efforts, reflecting a broader trend of limited scientific and policy attention towards parasitic diseases. Progress in this field will depend on improvements in diagnostic capacity, particularly the wider availability of sensitive molecular assays and antigen-detection tools suitable for routine clinical use. In parallel, further research into host–parasite interactions within neural tissues, determinants of neurotropism and the immunopathological drivers of tissue damage will be essential for refining therapeutic and preventive strategies.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182026102108.
Acknowledgements
The authors gratefully acknowledge Dr Muhammad Furquan Ashad for his assistance in revising the English of this manuscript.
Author contributions
GM contributed to conceptualization, draft writing, review, and editing. MTM contributed to draft writing and review. CT, AV, AS, SZ, LB and MD contributed to review and editing. FV contributed to conceptualization, draft writing, supervision and review.
Financial support
This paper received no specific grant from any funding agency, commercial or not-for-profit sectors.
Competing interests
The authors declare none.
Ethical standards
No ethical approval was required for this review.
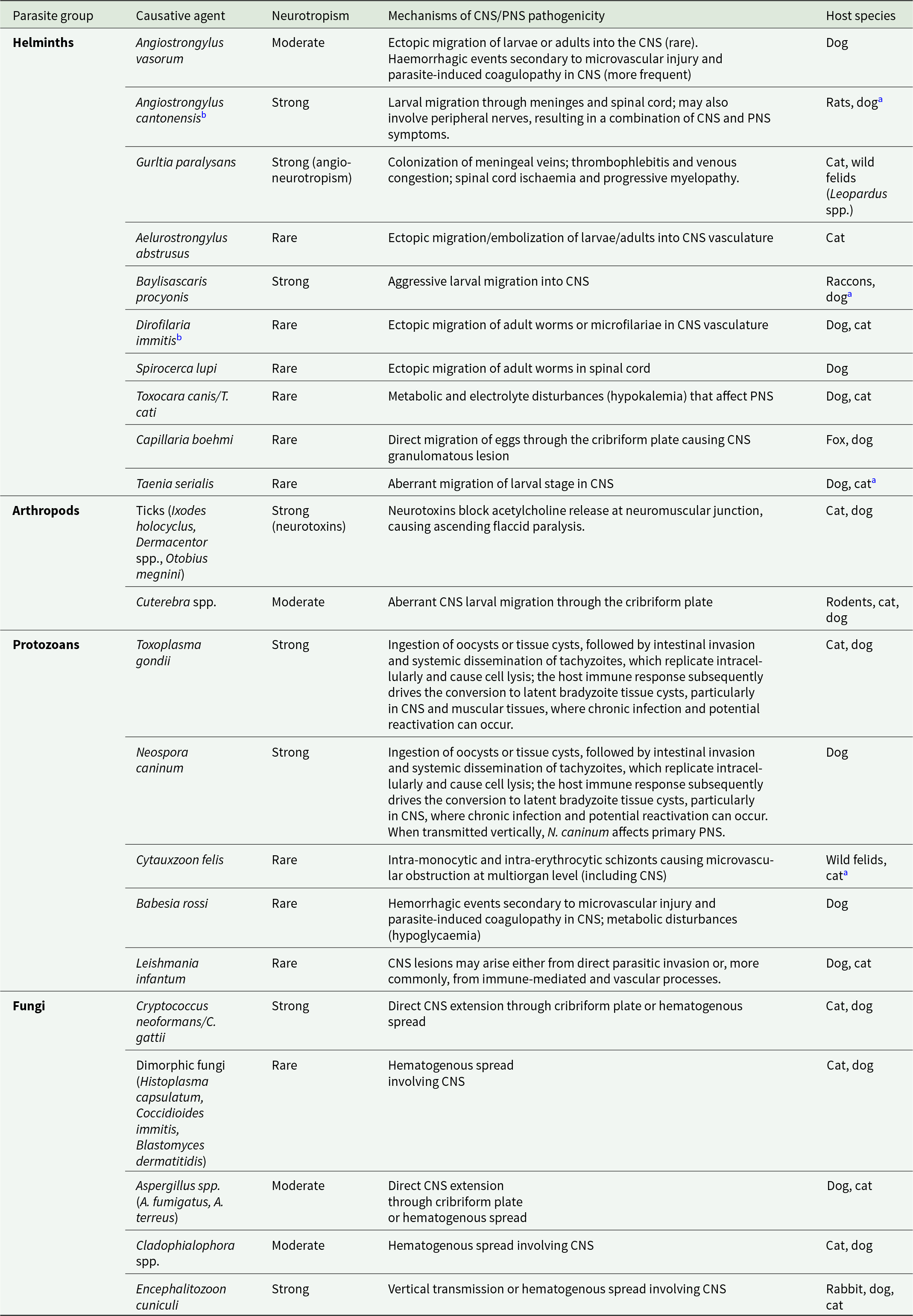
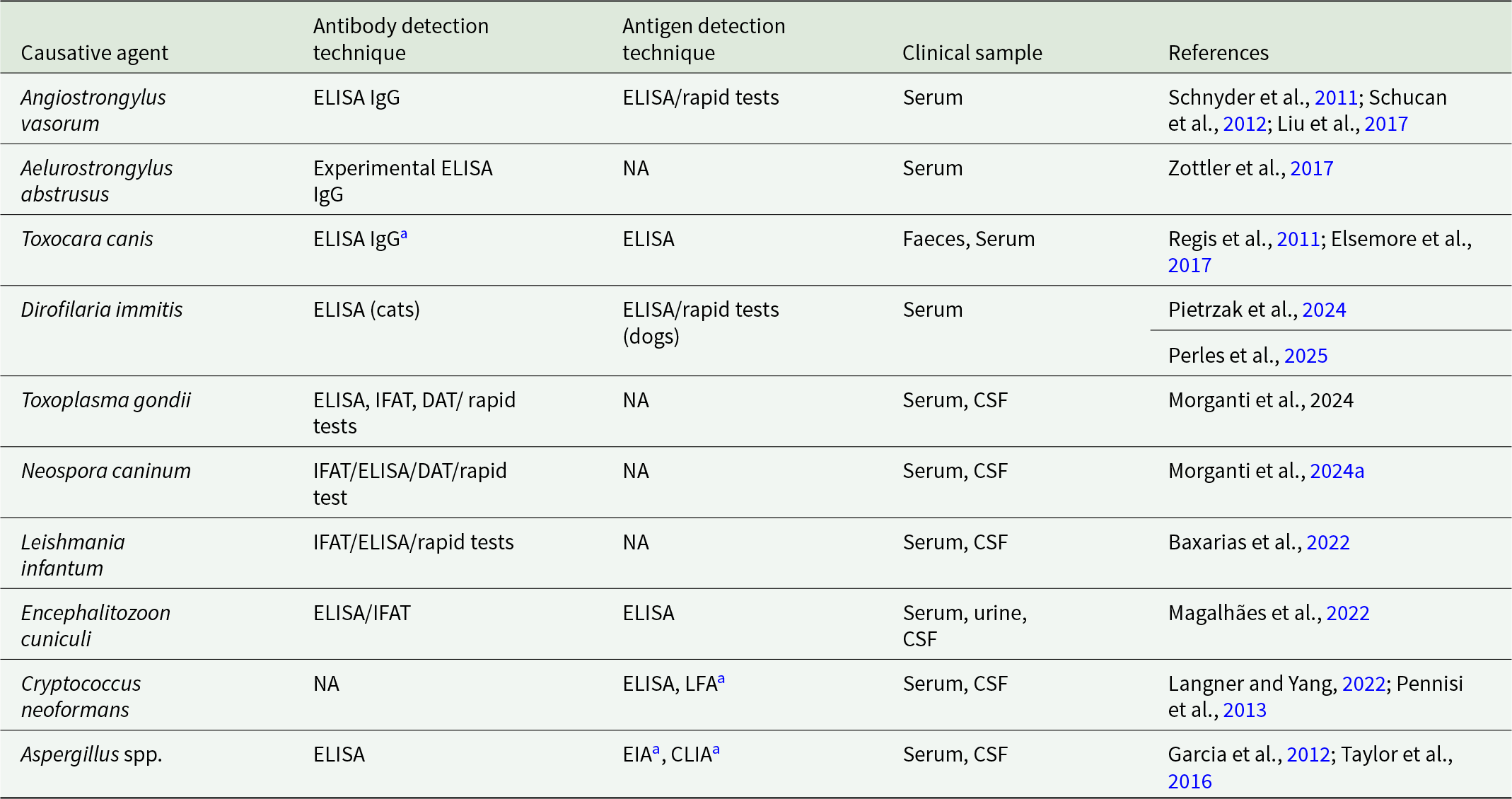
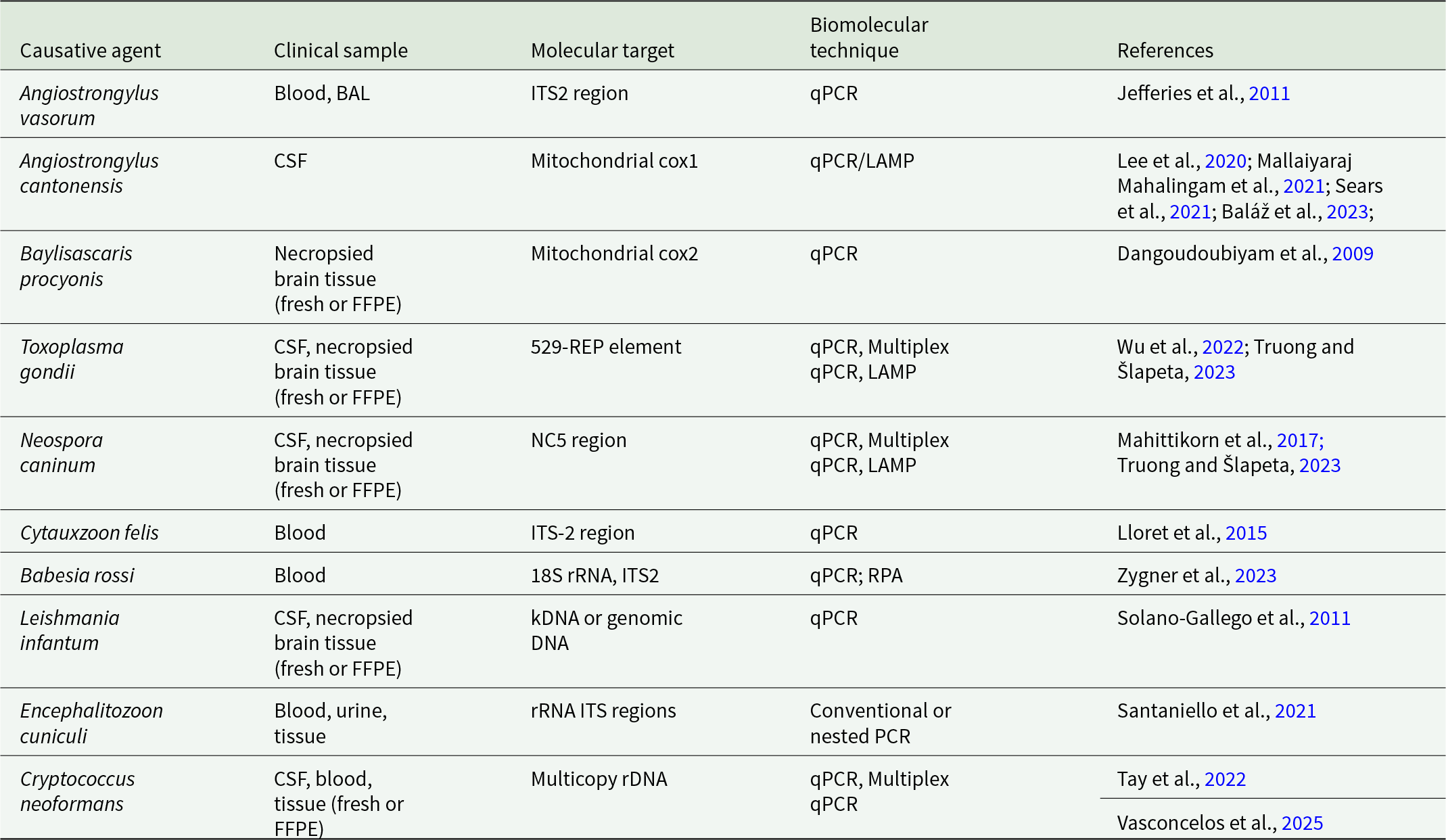
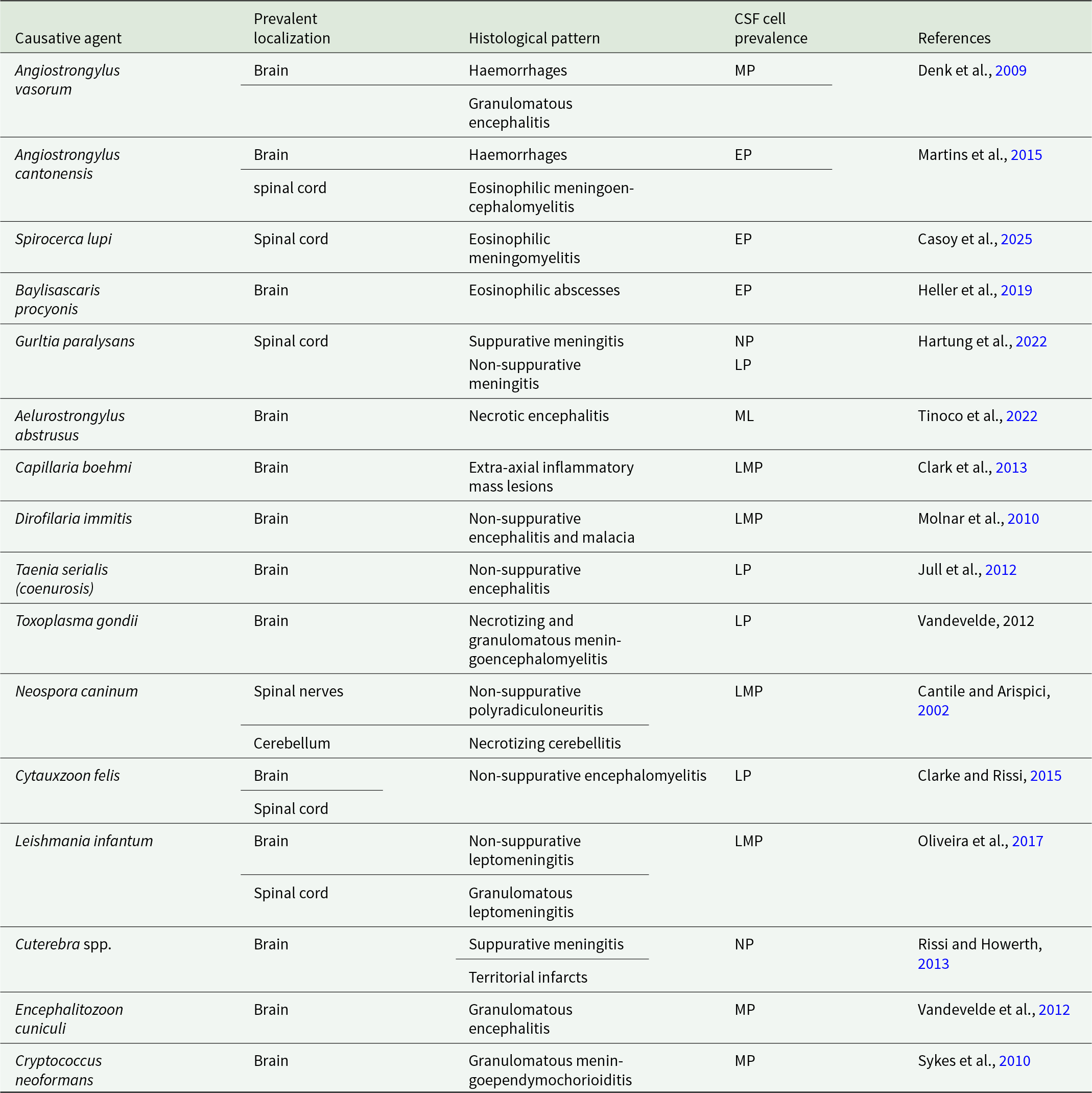
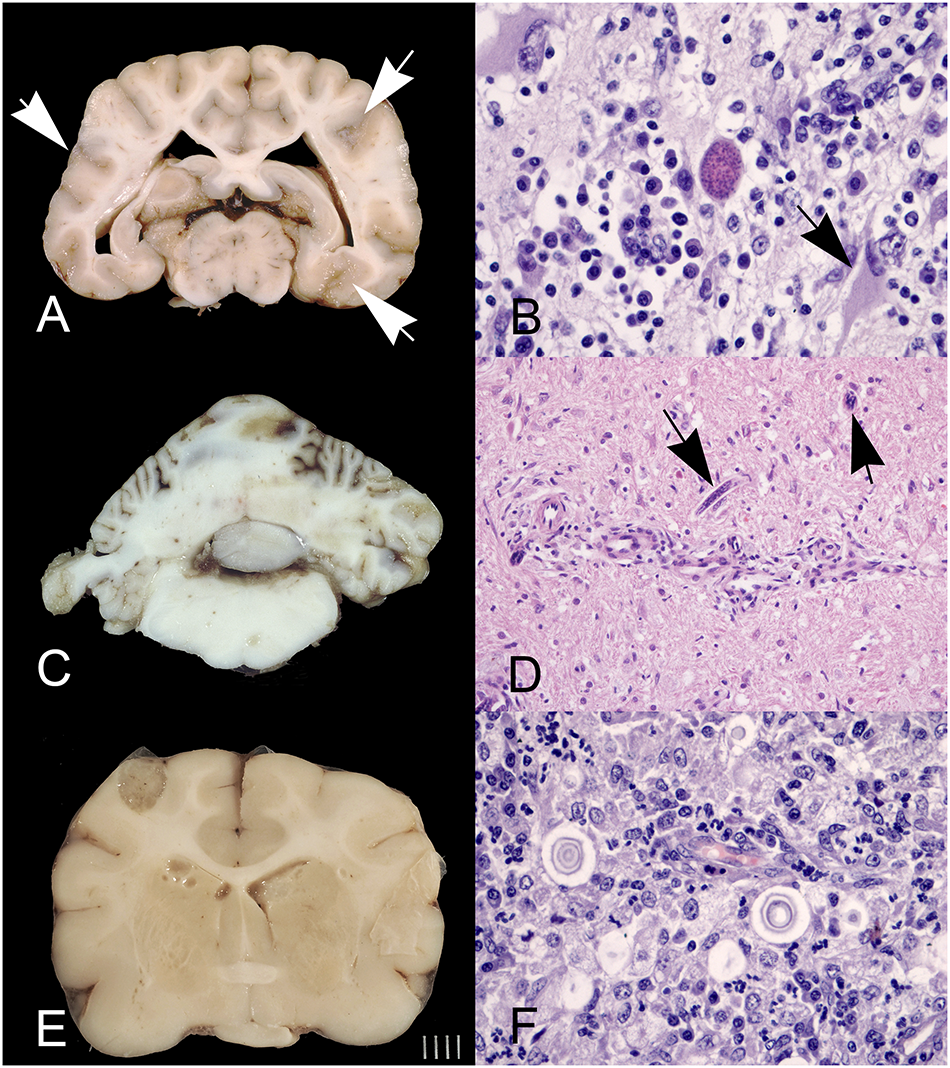



 Open access
Open access