



Introduction
The epidemiology of babesiosis
Global epidemiological status of babesiosis
Babesiosis has a global distribution. Since Babesia bigemina was first identified in cattle erythrocytes by Romanian scientist Babes in 1888, more than 250 species of Babesia have been formally reported worldwide. However, only a few species are known to infect humans, including Babesia microti, B. divergens, B. venatorum, B. duncani, B. bigemina, B. bovis, B. odocoilei, B. crassa, B. motasi, B. caballi, B. ovis, Babesia sp. XXB/Hangzhou and Babesia TianZhu-like (Chen et al., Reference Chen, Xue, Ai, Song, Cai and Chen2022; Calchi et al., Reference Calchi, Moore, Bartone, Kingston, André, Breitschwerdt and Maggi2024). The first human case of babesiosis was reported in Yugoslavia in 1957 (Skrabalo and Deanovic, Reference Skrabalo and Deanovic1957), and subsequent cases have been documented across the Americas, Europe, Asia, Africa and Australia, with the highest numbers reported in the Americas and Europe (Hildebrandt et al., Reference Hildebrandt, Gray and Hunfeld2013; Krause, Reference Krause2019; Kumar et al., Reference Kumar, O’bryan and Krause2021). Since babesiosis became a nationally notifiable disease in the USA in 2011, the disease has been endemic in the Northeastern and upper Midwestern US, with the annual number of reported cases increasing to approximately 2000–3000. In contrast, human babesiosis is only sporadically reported in Europe, with fewer than 10 confirmed cases documented annually, with B. microti being the predominant species responsible for the majority of cases (National Health and Family Planning Commission, 2017; Gray and Herwaldt, Reference Gray and Herwaldt2019; Kumar et al., Reference Kumar, O’bryan and Krause2021). In Europe, reported cases are relatively fewer, with B. divergens, B. venatorum and B. microti being the main species involved (Vannier and Krause, Reference Vannier and Krause2012; Hildebrandt et al., Reference Hildebrandt, Gray and Hunfeld2013; Krause, Reference Krause2019). In recent years, human babesiosis cases have also been reported in South Africa, Japan, Colombia, South Korea, France, Egypt, Australia, Ecuador, Mexico, Singapore and Tanzania (Bush et al., Reference Bush, Isaäcson, Mohamed, Potgieter and de Waal1990; Saito-Ito et al., Reference Saito-Ito, Tsuji, Wei, He, Matsui, Kohsaki, Arai, Kamiyama, Hioki and Ishihara2000; Ríos et al., Reference Ríos, Alvarez and Blair2003; Kim et al., Reference Kim, Cho, Joo, Tsuji, Cho, Park, Chung, Ju, Cheun, Lee, Lee and Kim2007; El-Bahnasawy et al., Reference El-Bahnasawy, Khalil and Morsy2011; Martinot et al., Reference Martinot, Zadeh and De Briel2012; Senanayake et al., Reference Senanayake, Paparini, Latimer, Andriolo, Dasilva, Wilson, Xayavong, Collignon, Jeans and Irwin2012; Peniche-Lara et al., Reference Peniche-Lara, Balmaceda, Perez-osorio and Munoz-zanzi2018; Bloch et al., Reference Bloch, Kasubi, Levin, Mrango, Weaver, Munoz and West2018a; Lim et al., Reference Lim, Chavatte, Vasoo and Yang2020; Calvopiña et al., Reference Calvopiña, Montesdeoca-andrade, Bastidas-caldes, Enriquez, Rodríguez-hidalgo, Aguilar-rodríguez and Cooper2023).
Epidemiological status of human babesiosis in China
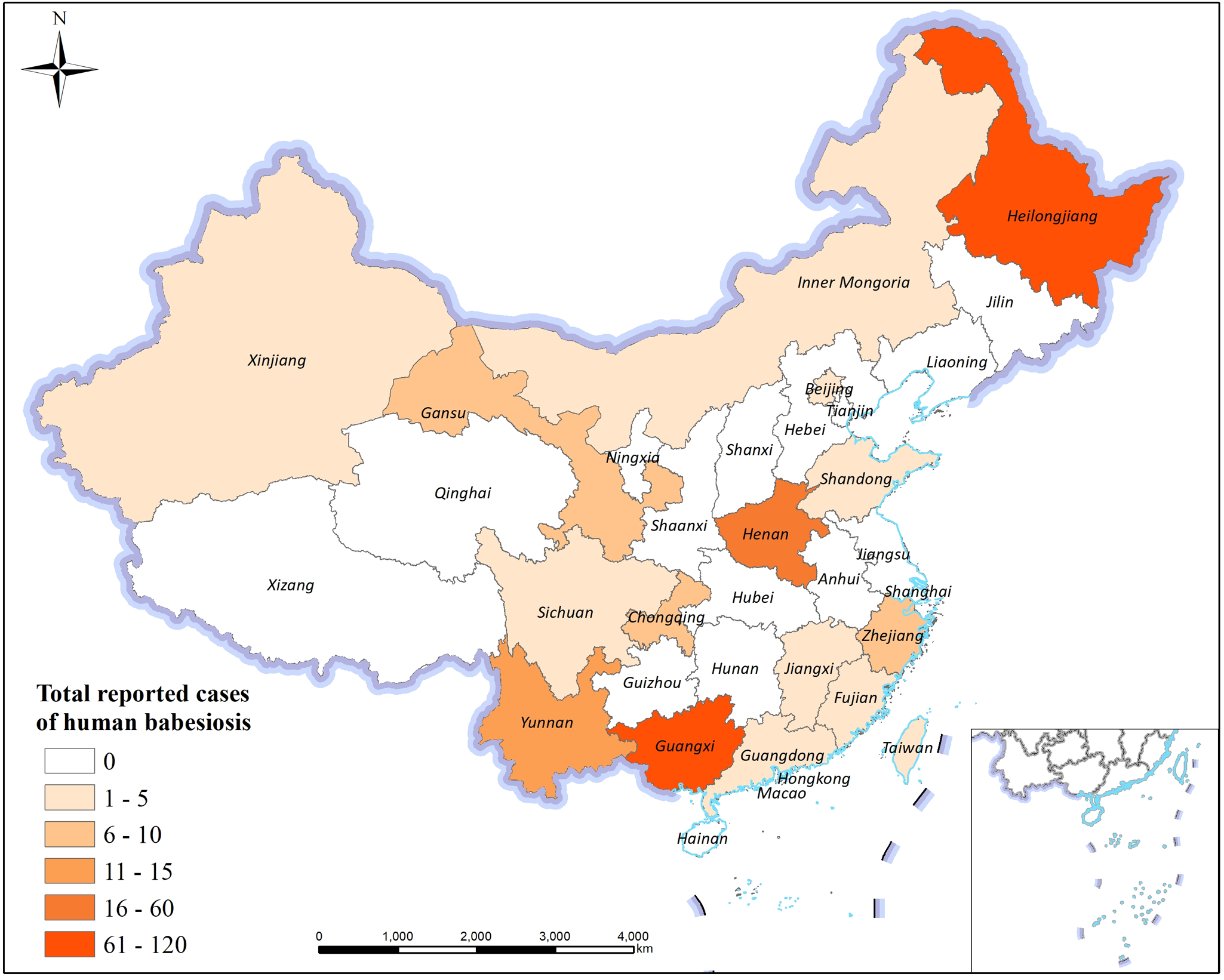
Since Hong Shilü first reported nine cases of human babesiosis-like infections in Beibei, Chongqing, China, in 1943 (Hong, Reference Hong1943a,b), which were later identified as the earliest documented human babesiosis cases in China based on the morphological characteristics of the parasite without malarial pigment (Qu, Reference Qu2007), a total of 328 human cases and asymptomatic infections have been documented nationwide by 2025. Notably, the case reported by Skrabalo and Deanovic in Yugoslavia in 1957 is recognized as the first confirmed human babesiosis case worldwide with definitive etiological identification. The cases in China are distributed among 16 of 34 provincial-level administrative regions, including Zhejiang, Yunnan, Guangxi, Heilongjiang, Fujian, Henan, Guangdong, Jiangxi, Beijing, Xinjiang and Inner Mongolia. The highest numbers of reported cases are concentrated in Heilongjiang, Guangxi, Henan and Yunnan, exhibiting a belt-like distribution pattern extending from northeastern to southwestern China (Figure 1) (Jia et al., Reference Jia, Zheng, Jiang, Jiang, Jiang, Wei, Liu, Huo, Sun, Chu, Fan, Chang, Yao, Zhang, Wang, Guo, Fu, Wang, Krause, Song and Cao2018; Zhang, Reference Zhang2022; Ji et al., Reference Ji, Jiang, He, Wang, Yue, Li, Yang, Wang, Zhang and Zhou2023; Li et al., Reference Li, Zhang, Zhou, Li, Song, Song, Li, Xiangzheng and Wu2023; Chen et al., Reference Chen, Shi, Tuerdali, Shi, Zhan, Fang and Wang2025; Shi et al., Reference Shi, Zhou, Cao, Wang, Zhang, Xia, Xu and Zhou2025; Wang et al., Reference Wang, Zhang, Shao, Li, Pu, He, Yao, Bie, Jiang and Du2025). The primary causative species identified include B. microti, B. venatorum, B. divergens, B. crassa-like organisms, B. microti-like species, the novel B. XXB/Hangzhou and several unclassified Babesia species (Chen et al., Reference Chen, Li, Gao, Bian, Yan, Kong and Liu2019, Reference Chen, Liu, Xue, Chen, Huang, Yu, Cai, Lu, Song, Zhang, Ai and Chen2020; Zhou et al., Reference Zhou, Wang, Xue, Xia and Zhou2019).
Distribution map of babesiosis in China from 1943 to 2025.

Figure 1 Long description
The map of China illustrates the total reported cases of human babesiosis across various regions. The legend categorizes cases into six ranges: 0, 1-5, 6-10, 11-15, 16-60 and 61-120. Regions with 0 cases are shown in a light shade, while 1-5 cases are slightly darker. Areas with 6-10 cases include regions like Jiangxi and Fujian. Regions with 11-15 cases include Henan and Guangdong. Guangxi and Yunnan fall into the 16-60 range. Heilongjiang has the highest range of 61-120 cases. The map includes a scale bar and a compass rose for orientation.
In terms of the transmission route composition of the 328 included cases, 61 are transfusion-related asymptomatic Babesia infections, all identified through systematic nucleic acid and serological screening of blood donors in endemic areas. These include 48 cases of B. microti infection reported in a 2016 blood donor screening study in Guangxi Province (Wang et al., Reference Wang, Peng, Zhu and Xue2016) and 13 cases of B. microti infection detected in a 2018 blood donor screening program in Heilongjiang Province (Bloch et al., Reference Bloch, Yang, He, Tonnetti, Liu, Wang, Guo, Li, Leiby and Shan2018b).
From 2022 to 2025, a total of 8 cases were reported, including 2 cases from Zhejiang Province (Xu et al., Reference Xu, Li, Zhang, Yu and Ruan2023; Lu et al., Reference Lu, Zhang, Han, Yu, Ye and Zheng2024), 1 case from Inner Mongolia (Huang et al., Reference Huang, Sun, Huo, Xu, Xia, Yang, Hong, Huang, Nie, Liao, Zhang, Zhu, Li, Ma, Zhang, Li, Huang, Wang, Cao, Wang and Jiang2023), 1 case from Fujian Province (Lin et al., Reference Lin, Chen, Xie, Ouyang and Xiao2025), 2 cases from Jiangxi Province including one misdiagnosed as lupus erythematosus (Tao et al., Reference Tao, Guo and Wang2024), 1 case of China’s first imported human babesiosis in Beijing was confirmed on 14 July 2023 (Huang et al., Reference Huang, Xiang, Liao, Luan, Zhao, Yang, Luo, Huang, Xia, Zhu, Sun, Wang and Jiang2025), and 1 imported case from the USA reported in Zhuhai City, Guangdong Province on 8 September 2023 (Liu et al., Reference Liu, Ji, Huang, Huang, Li, Liu and Xia2024; Yangyang and Liang, Reference Yangyang and Liang2024). One fatal case was documented in Zhejiang Province (Xu et al., Reference Xu, Li, Zhang, Yu and Ruan2023), and 1 severe case was reported in Jiangxi Province (Xu et al., Reference Xu, Fei, Gong, Xie, Xu, Liu, Xie, Zhou and Xie2024; Luo and Xiao, Reference Luo and Xiao2025), both involving cancer patients with co-infections and severe immune deficiency. As of now, at least 328 cases have been reported in China (Table 1, Table 2 and Figure 1).
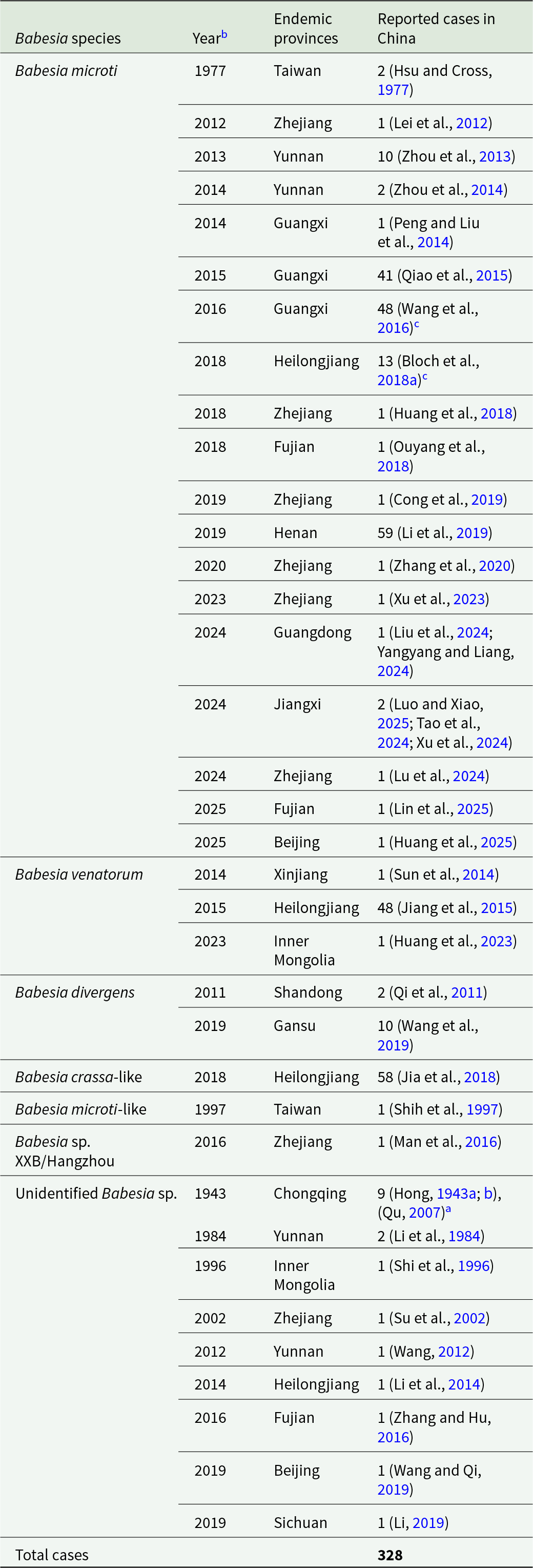
Reported human babesiosis cases in China by Babesia species and geographic region

Table 1 Long description
The table lists reported human babesiosis case counts in China by Babesia species and the province or region where cases were reported, with literature citations for each entry. Total reported cases are 328. Babesia microti accounts for most reports, with notable concentrations in Guangxi (90) and Henan (59), followed by Heilongjiang (13) and Yunnan (12), and smaller counts across several other provinces including Zhejiang (6) and multiple single-case locations. Other species contribute fewer cases overall: Babesia venatorum is mainly reported in Heilongjiang (48) with single cases in Xinjiang and Inner Mongolia; Babesia crassa-like is reported in Heilongjiang (58). Babesia divergens is reported in Gansu (10) and Shandong (2). Several entries are listed as unidentified Babesia species, led by Chongqing (9) with smaller counts in Yunnan (3) and single cases in multiple provinces. Counts reflect published reports and may be influenced by differences in surveillance, diagnostic methods, and reporting across regions and time.
a Expert review and commentary.
Reported cases of Babesia infections in China from 1943 to 2025

Table 2 Long description
The table lists reported human Babesia infections in China by species, report year, and province, with a national total of 328 cases. Babesia microti dominates the reports and spans many provinces and years, including notable clusters in Henan in 2019 with 59 cases and in Guangxi in 2015 and 2016 with 41 and 48 cases. Several entries are marked as transfusion-related, including Babesia microti reports in Guangxi in 2016 and Heilongjiang in 2018. Other species are less frequent: Babesia venatorum includes 48 cases in Heilongjiang in 2015 plus single cases in Xinjiang in 2014 and Inner Mongolia in 2023; Babesia divergens appears in Shandong in 2011 with 2 cases and Gansu in 2019 with 10 cases. Additional single-species reports include Babesia crassa-like with 58 cases in Heilongjiang in 2018, Babesia microti-like with 1 case in Taiwan in 1997, and Babesia sp.XXB/Hangzhou with 1 case in Zhejiang in 2016. Unidentified Babesia reports occur intermittently from 1943 onward, including 9 cases in Chongqing in 1943 and mostly single-case reports in later years. Counts reflect published case reports and may not represent true incidence or complete surveillance coverage.
a Expert review and commentary.
b All years mentioned in this article are case report years.
c Transfusion-related Babesia infection cases.
Transmission ecology of human babesiosis
Tick vector transmission
The primary vectors of B. microti are ticks of the genus Ixodes, such as Ixodes scapularis, I. ricinus and I. persulcatus. I. ricinus is a known vector for B. venatorum, while Dermacentor albipictus may serve as a vector for B. duncani (Swei et al., Reference Swei, O’connor, Couper, Thekkiniath, Conrad, Padgett, Burns, Yoshimizu, Gonzales, Munk, Shirkey, Konde, Ben Mamoun, Lane and Kjemtrup2019). In Europe, I. ricinus is the principal vector for B. divergens (Bonnet et al., Reference Bonnet, Jouglin, Malandrin, Becker, Agoulon, L’hostis and Chauvin2007), although B. divergens-like pathogens have also been detected in I. persulcatus and I. ovatus (Zamoto-Niikura et al., Reference Zamoto-Niikura, Tsuji, Imaoka, Kimura, Morikawa, Holman, Hirata and Ishihara2014; Jiang, Reference Jiang2016; Wang et al., Reference Wang, Teng, Wan, Wang, Yuan, Huang, Huang, Zhao, Wang, Cao, Huang, Xu, Yuan and Qin2024). Artificial infection experiments indicate that both Rhipicephalus haemaphysaloides and Hyalomma asiaticum are capable of transmitting B. microti (Li et al., Reference Li, Zhu, Zhang, Zhang and Zhou2016; Wu et al., Reference Wu, Cao, Zhou, Zhang, Gong and Zhou2017).
Ticks in China exhibit high species diversity and broad geographical distribution, yet investigations on human-infecting Babesia species carried by ticks have been limited to certain regions (Li and Zhang, Reference Li and Zhang2019) (Table 3). In Northeast China, I. persulcatus, Haemaphysalis concinna and Dermacentor silvarum are recognized vectors, with B. divergens, B. venatorum and B. crassa-like parasites identified in these tick species (Fan et al., Reference Fan, Li, Xu, Hu, Zhang and Sun2012; Jiang, Reference Jiang2016; Jia et al., Reference Jia, Zheng, Jiang, Jiang, Jiang, Wei, Liu, Huo, Sun, Chu, Fan, Chang, Yao, Zhang, Wang, Guo, Fu, Wang, Krause, Song and Cao2018). B. microti has been detected in Haemaphysalis longicornis in Henan, Shandong and Zhejiang (Chen et al., Reference Chen, Liu, Liu, Xu, Lv, Xia and Zhou2014; Zhang et al., Reference Zhang, Sun, Jiang and Huo2017; Wang et al., Reference Wang, Chen, Du, Wu, Yang, Zhao and Du2023), while Rhipicephalus sanguineus infected with B. microti has been reported in Guangxi and Zhejiang (Qiao et al., Reference Qiao, Peng, Zhu and Yan2015; Wang et al., Reference Wang, Chen, Du, Wu, Yang, Zhao and Du2023). In Inner Mongolia, Dermacentor nuttalli, H. asiaticum and Dermacentor niveus have been found infected (Hao et al., Reference Hao, Li, Sun, Ge, Qiao, Li, Tian, Shi and Yang2009). Yunnan Province reported infections in Ixodes ovatus, I. granulatus, Rhipicephalus microplus and Haemaphysalis montgomeryi (Wang, Reference Wang2021).
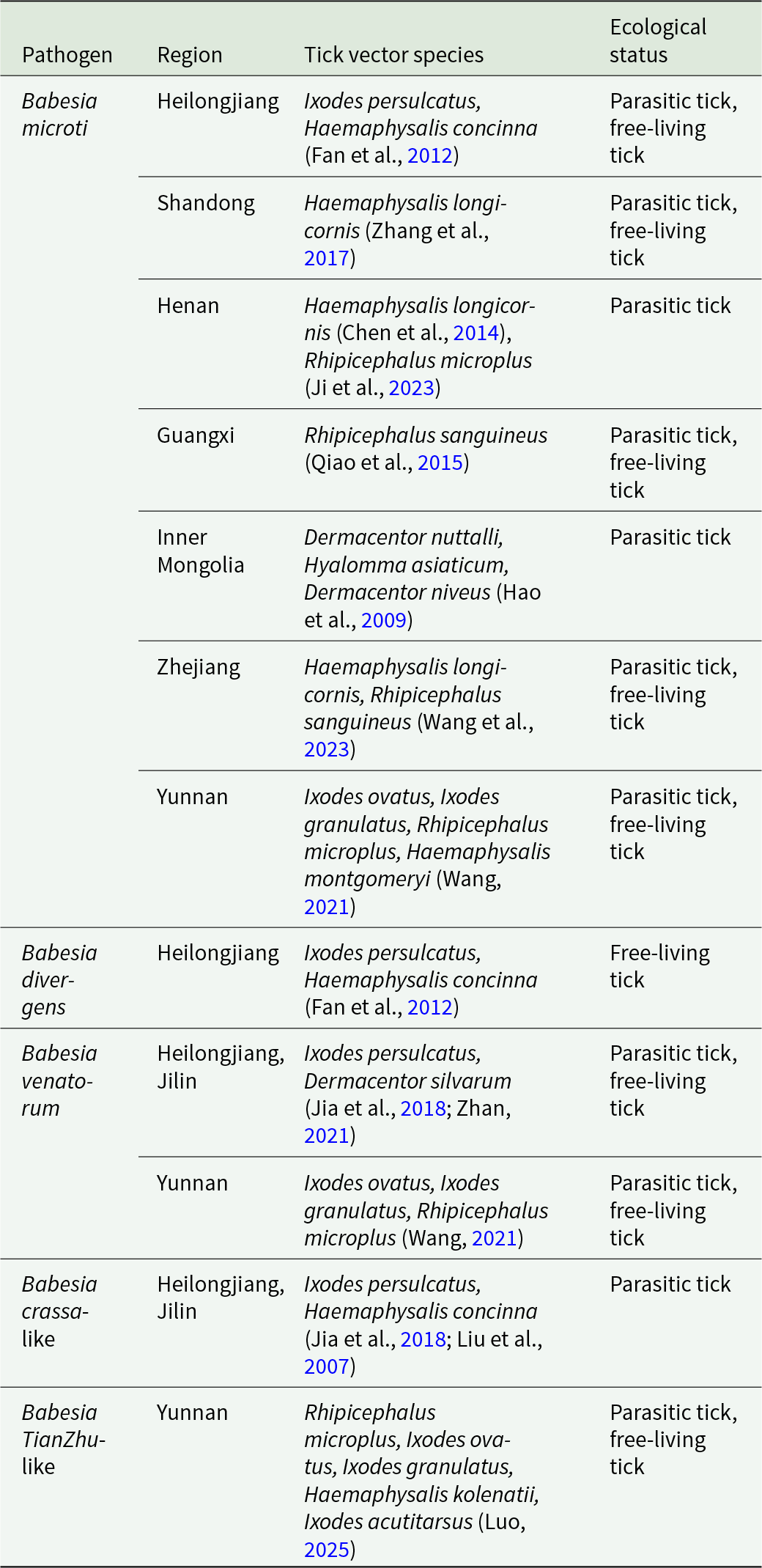
The transmission vectors of human babesiosis in China

Table 3 Long description
The table lists human babesiosis pathogens in China by region, the tick species reported as vectors, and whether those ticks are described as parasitic, free-living, or both. Babesia microti has the widest regional coverage, appearing in Heilongjiang, Shandong, Henan, Guangxi, Inner Mongolia, Zhejiang, and Yunnan, with vectors including Ixodes persulcatus, Haemaphysalis longicornis, and Rhipicephalus species. Several regions report more than one vector for Babesia microti, such as Henan and Yunnan. Babesia divergens is only reported in Heilongjiang and is linked to Ixodes persulcatus and Haemaphysalis concinna, with ecological status noted as free-living tick. Babesia venatorum is reported in Heilongjiang and Jilin with Ixodes persulcatus and Dermacentor silvarum, and also in Yunnan with Ixodes and Rhipicephalus microplus. Babesia crassa-like is limited to Heilongjiang and Jilin and is associated with Ixodes persulcatus and Haemaphysalis concinna, listed as parasitic tick. Babesia TianZhu-like is reported in Yunnan with multiple vectors including Rhipicephalus microplus and several Ixodes species, and includes both parasitic and free-living ecological statuses. Vector and ecological-status entries reflect cited reports and may vary by study methods and local sampling rather than indicating nationwide distributions.
Animal reservoir hosts
The reservoir hosts of different Babesia species vary significantly: B. microti primarily infects rodents; the main host of B. divergens is cattle, while cervids can also serve as reservoirs; B. venatorum predominantly utilizes cervids as its reservoir hosts; and the host of B. duncani remains unclear (Hao et al., Reference Hao, Li, Sun, Ge, Qiao, Li, Tian, Shi and Yang2009). In China, the primary reservoir hosts for B. microti are rodents, such as Apodemus agrarius, Rattus norvegicus, Microtus spp. and other wild rodents (Saito-Ito et al., Reference Saito-Ito, Takada, Ishiguro, Fujita, Yano, Ma and Chen2008; Fan et al., Reference Fan, Li, Xu, Hu, Zhang and Sun2012; Zhao et al., Reference Zhao, Li, Sun, Zhang, Jiang, Liu and Cao2013; Cao et al., Reference Cao, Ni, Zhang and Ni2016; Chen et al., Reference Chen, Ye, Fan, Li, Tang, Liu, Ren and Bai2017; Xiao et al., Reference Xiao, Peng, Xu, Chen, Lin and Deng2017; Wang et al., Reference Wang, Bie, Zhang, Song, Duan, Xiong, Wang, Jiang, Gao, Dong and Du2021; Ren et al., Reference Ren, Liu, Niu, Song, Cheng, Zhao, Cui, Chen, Bai, Rao and Yu2025), with infection rates reaching 28.0% in Heilongjiang (Fan et al., Reference Fan, Li, Xu, Hu, Zhang and Sun2012) and 28.9% in Fujian (Xiao et al., Reference Xiao, Peng, Xu, Chen, Lin and Deng2017). B. microti has also been detected in crab-eating monkeys in Guangxi (Lu et al., Reference Lu, Peng, Zhu, Li and Xue2016). B. divergens is mainly found in rodent-like animals (Fan et al., Reference Fan, Li, Xu, Hu, Zhang and Sun2012), and goats can be experimentally infected; B. venatorum has been detected in sheep (Jia et al., Reference Jia, Zheng, Jiang, Jiang, Jiang, Wei, Liu, Huo, Sun, Chu, Fan, Chang, Yao, Zhang, Wang, Guo, Fu, Wang, Krause, Song and Cao2018; Wang, Reference Wang2021), while B. crassa-like parasites have only been reported in sheep (Jia et al., Reference Jia, Zheng, Jiang, Jiang, Jiang, Wei, Liu, Huo, Sun, Chu, Fan, Chang, Yao, Zhang, Wang, Guo, Fu, Wang, Krause, Song and Cao2018) (Table 4). The newly identified human-infecting species, Babesia TianZhu-like, has been detected in both sheep and cattle (Luo, Reference Luo2025).
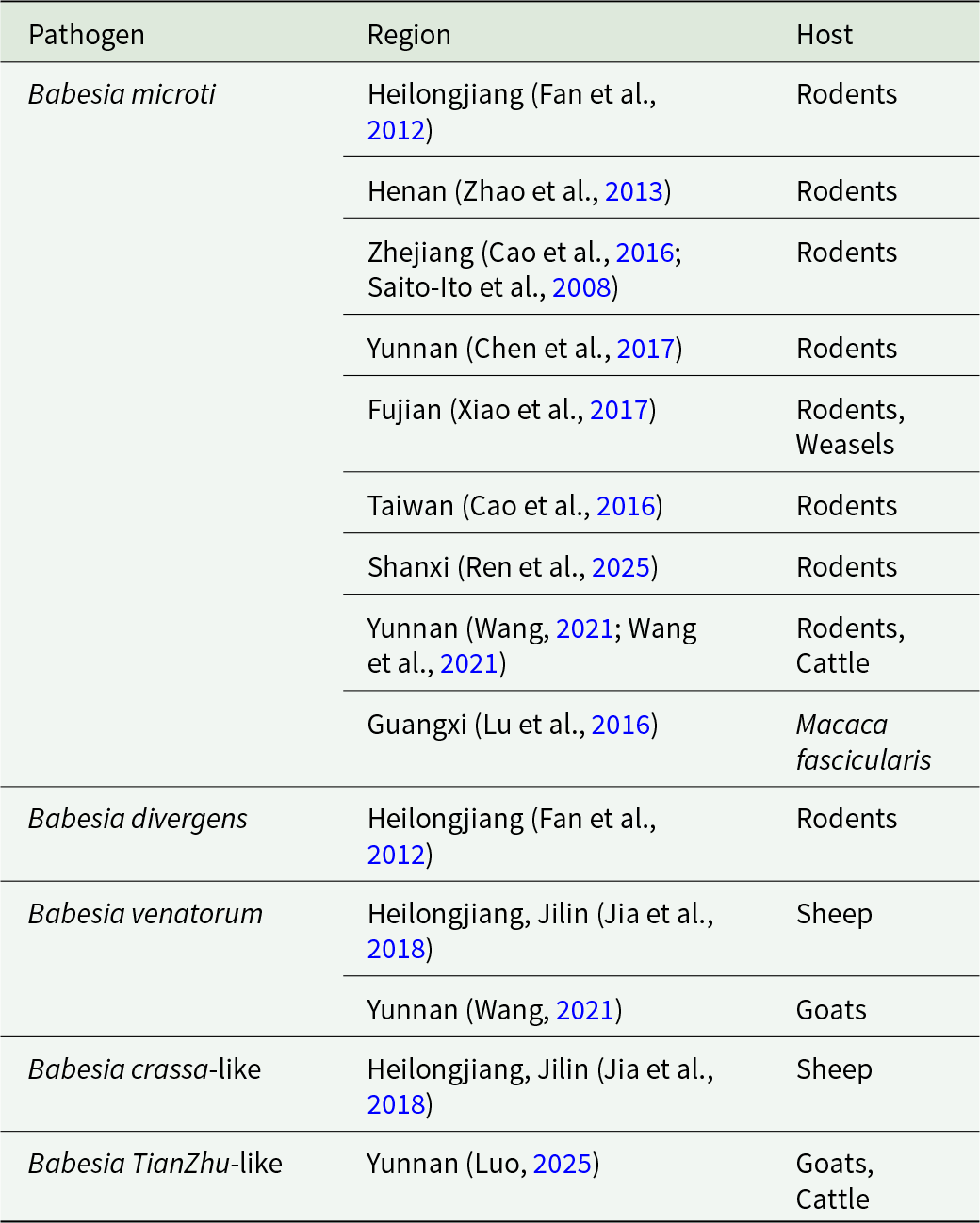
Animal hosts of Babesia in humans in China

Table 4 Long description
The table lists Babesia pathogens reported in humans in China and the animal hosts associated with each report by region. Babesia microti appears in the most locations, spanning Heilongjiang, Henan, Zhejiang, Yunnan, Fujian, Taiwan, Shanxi, and Guangxi, and is most often linked to rodents. For Babesia microti, additional hosts are noted in some regions, including weasels in Fujian, cattle in Yunnan, and Macaca fascicularis in Guangxi. Babesia divergens is reported only in Heilongjiang and is associated with rodents. Babesia venatorum and Babesia crassa-like are reported in Heilongjiang and Jilin with sheep, and Babesia venatorum is also reported in Yunnan with goats. Babesia TianZhu-like is reported in Yunnan with goats and cattle. Host listings reflect reported associations in the cited regional studies and do not indicate how common each host is or prove transmission direction.
Clinical manifestations of human babesiosis
The clinical manifestations of babesiosis are associated with the host’s immune status and the extent of Babesia-induced erythrocyte destruction, with an incubation period typically ranging from 1 to 4 weeks (China, 2017). In immunocompetent individuals, most infections present with mild or flu-like symptoms and follow a self-limiting course lasting approximately 2–4 weeks. Common symptoms include chills, fever, nausea, loss of appetite, excessive sweating, fatigue, myalgia, arthralgia, headache, abdominal discomfort and anaemia. In severe cases, which often occur abruptly in splenectomized, elderly, debilitated or immunocompromised patients, manifestations may include high fever, marked anaemia, jaundice, hemoglobinuria, dyspnea, renal insufficiency, cardiac complications, altered mental status and potentially life-threatening complications (Vannier and Krause, Reference Vannier and Krause2012; China, 2017; Chen et al., Reference Chen, Liu, Xue, Chen, Huang, Yu, Cai, Lu, Song, Zhang, Ai and Chen2020). Disease severity varies distinctly among Babesia species in China: B. microti causes moderate-severe infection with recurrent fever, progressive cytopenia, hemolytic anaemia and even hemophagocytic syndrome; B. venatorum induces moderate-severe acute infection with persistent high fever, severe anaemia and systemic inflammation; B. divergens shows obvious heterogeneity, with most asymptomatic/self-limiting and a few severe cases presenting with high fever, severe hemolysis and liver/renal injury; B. crassa-like leads to mild-moderate infection with non-specific symptoms (headache, low-grade fever) and mild liver dysfunction; B. sp. XXB/Hangzhou only causes chronic mild infection with recurrent low-grade fever and fatigue for up to 10 years (Man et al., Reference Man, Qiao, Cui, Feng, Fu and Cheng2016).
Laboratory diagnostic methods for human babesiosis
Blood smear microscopy
Blood smear microscopy is a classic and effective method for diagnosing human babesiosis. After Giemsa staining, the Babesia cytoplasm appears blue and the nucleus stains purplish-red (Xu et al., Reference Xu, Li, Zhang, Yu and Ruan2023, Reference Xu, Fei, Gong, Xie, Xu, Liu, Xie, Zhou and Xie2024; Yangyang and Liang, Reference Yangyang and Liang2024; Lin et al., Reference Lin, Chen, Xie, Ouyang and Xiao2025). The morphology of Babesia within red blood cells is diverse, with common forms including ring-shaped, round, rod-shaped, dot-like, pear-shaped and amoeboid forms. Notably, the ring forms of Babesia highly resemble those of Plasmodium species, but Babesia lacks malarial pigment and often forms characteristic tetrads, which are key morphological differences. This similarity can lead to misdiagnosis, especially in southwestern China, where malaria was historically endemic and co-infection or misidentification frequently occurs. In case reports from Wenzhou, Wright staining revealed purplish-red nuclei and blue cytoplasm, presenting as ring-shaped or teardrop-shaped structures (Xu et al., Reference Xu, Fei, Gong, Xie, Xu, Liu, Xie, Zhou and Xie2024). In the first reported imported case of B. microti infection in Zhuhai, Giemsa staining showed ring or teardrop-shaped parasites (Yangyang and Liang, Reference Yangyang and Liang2024). In cases from Fujian, ring, round and rod-shaped forms were identified, some resembling ring-like structures (Lin et al., Reference Lin, Chen, Xie, Ouyang and Xiao2025).
Nucleic acid detection
Nucleic acid detection techniques such as polymerase chain reaction (PCR) and nucleic acid amplification test (NAT) for Babesia DNA and RNA can specifically amplify Babesia-specific gene fragments with high sensitivity and strong specificity. Common techniques include nested PCR, recombinase polymerase amplification, quantitative real-time PCR (qPCR), reverse line blot hybridization, genechip, loop-mediated isothermal amplification, metagenomics, genechip (Chen et al., Reference Chen, Ai, Chen, Feng, Chen, Cai, Lu, Zhou, Chen and Hu2016) and multiplex PCR (Calchi et al., Reference Calchi, Moore, Bartone, Kingston, André, Breitschwerdt and Maggi2024; Mittal et al., Reference Mittal, Chakravarti, Kundu, Tripathi and Batra2025).
Animal inoculation
Anticoagulated patient samples can be intraperitoneally inoculated into BALB/c mice, SCID mice, NOD-SCID mice or hamsters. Approximately 1 week post-inoculation, blood smears are prepared from tail blood and examined microscopically, with the observation of Babesia parasites within erythrocytes considered positive (Pruthi et al., Reference Pruthi, Marshall, Wiltsie and Persing1995; China, 2017). Gerbils, hamsters, SCID mice and BALB/c mice can serve as animal models for Babesia inoculation and are used in the diagnosis of species such as B. duncani, B. divergens and B. microti. For instance, C3H/HeJ or SCID mice were infected by injection with 1 × 10⁶ B. duncani WA-1 strain or 1 × 10⁷ B. microti LabS1 strain obtained from previously infected mouse blood (Chand et al., Reference Chand, Vydyam, Pal, Thekkiniath, Darif, Li, Choi, Magni, Luchini, Tonnetti, Horn, Tufts and Ben Mamoun2024). B. microti was isolated using NOD-SCID mice and subsequently passaged into BALB/c mice to successfully establish an infection model (Song et al., Reference Song, Cai, Lu, Ai, Chen, Chen and Chen2022).
In vitro culture
In vitro culture, which enables the propagation of Babesia parasites under strictly controlled laboratory conditions, is a valuable tool for the diagnosis and research of Babesia infections. Continuous in vitro culture systems have been successfully established for several Babesia species, including B. duncani (Guo et al., Reference Guo, He, Zhao, Zhu and Mamoun2019) in human serum, as well as Babesia sp. Xinjiang in small ruminants erythrocytes (Guan et al., Reference Guan, Ma, Liu, Du, Ren, Li, Wang, Liu, Yin and Luo2012). Recent advances in culture techniques have enabled the long-term maintenance of B. duncani in human or animal red blood cells via optimized culture systems, such as HL-1 medium supplemented with serum, paired with host erythrocytes. These optimized culture systems further facilitate drug sensitivity testing, antigen production and in-depth studies on parasite biology and host-parasite interactions (Abraham et al., Reference Abraham, Brasov, Thekkiniath, Kilian, Lawres, Gao, DeBus, He, Yu, Zhu, Graham, Liu, Molestina and Ben Mamoun2018). In China, this technique has been widely applied to isolate and propagate B. microti strains from clinical cases and animal hosts, providing essential biological materials for molecular characterization and vaccine development. Despite its broad utility, in vitro culture remains technically demanding and is not suitable for routine primary diagnosis. Currently, it is mainly used as a confirmatory method in reference laboratories, and remains an indispensable tool for basic and translational research on Babesia.
Metagenomic next-generation sequencing
Metagenomic next-generation sequencing (mNGS) is a high-throughput pathogen detection method based on next-generation sequencing technology that enables unbiased detection of nucleic acid sequences from all microorganisms, including bacteria, viruses, fungi and parasites, directly from clinical samples such as blood, cerebrospinal fluid and tissues without prior culture or presumptions about specific pathogens. This technique has gained prominence in recent years for diagnosing infectious diseases, especially for identifying pathogens in complex, severe and/or immunocompromised cases. Clinically, mNGS is commonly used to assist in the diagnosis of Babesia infections, where it has successfully identified B. microti in sputum, bronchoalveolar lavage fluid and blood samples, with confirmation by peripheral blood smears providing critical support for rapid identification of rare pathogens (Lu et al., Reference Lu, Zhang, Han, Yu, Ye and Zheng2024). In Zhuhai’s first imported case of human B. microti infection, mNGS provided substantial sequence data serving as a key diagnostic reference (Yangyang and Liang, Reference Yangyang and Liang2024). In China’s first infantile babesiosis case, both the infant and mother were confirmed to be infected with Babesia microti via combined PCR and mNGS detection (Liu et al., Reference Liu, Li, Wang, Gao, Xu, Zhao, Liu, Chen, Zhou, Cai and He2026).
Immunological detection
As of now, the only standardized serological technique for diagnosing human babesiosis caused by B. microti is the indirect fluorescent antibody test (Sun et al., Reference Sun, Han, Feng and Hu2013), which is crucial for transfusion-transmission studies (Herwaldt et al., Reference Herwaldt, Linden, Bosserman, Young, Olkowska and Wilson2011; Moritz et al., Reference Moritz, Winton, Tonnetti, Townsend, Berardi, Hewins, Weeks, Dodd and Stramer2016). Enzyme-linked immunosorbent assay (ELISA) and Western blot have potential promise for human B. sp. antibody detection, with Western blot as a high-specificity confirmatory assay for ELISA-positive results, though the cited ELISA/Western-blot protocols target Theileria equi and B. caballi and are not validated for human babesiosis (G. Yang et al., Reference Yang, Zhou, Chen, Hu, Guo, Wang and Du2022b). As a rapid on-site diagnostic tool, the immunochromatographic test can simultaneously detect antibodies against T. equi and B. caballi within 10–15 minutes (G. Yang et al., Reference Yang, Chen, Guo, Hu, Qi, Liu, Wang, Du and Wang2022a), with agreement rates with cELISA of 93% and 92.9%, respectively, and is currently applied for import/export certification rather than human babesiosis diagnosis (Jongejan et al., Reference Jongejan, Du, Papadopoulos, Blanda, Di Bella, Cannella, Guercio, Vicari, Tirosh-levy, Steinman, Baneth, van Keulen, Hulsebos, Berger and Wang2024).
Recent advances in technology
In recent years, significant progress has been made in diagnostic techniques for babesiosis. Antigen capture assays such as B. duncani antigen capture assays (BdACAs) target secreted biomarkers (BdV38 and BdV234) to specifically detect active infections with a sensitivity of 115 infected erythrocytes per microliter of blood, and effectively distinguish active infections from residual parasite DNA post-treatment – an ability validated by in vitro and in vivo tafenoquine treatment models, where BdACA antigen signals declined and vanished with parasitemia clearance while qPCR detected persistent parasite DNA in the same samples (Chand et al., Reference Chand, Vydyam, Pal, Thekkiniath, Darif, Li, Choi, Magni, Luchini, Tonnetti, Horn, Tufts and Ben Mamoun2024). This specificity derives from BdV38 and BdV234 being exclusively secreted by viable, replicating parasites via vesicular transport with levels linearly correlating with parasitemia; these biomarkers are no longer detectable after parasite elimination, thus avoiding PCR false positives for active infection caused by residual non-viable parasite DNA. Meanwhile, automated fluorescent flow cytometry enables rapid, automated detection and quantification of Babesia parasites within red blood cells. Although limited in species identification, its high throughput and objective quantification offer an efficient alternative for clinical use (Vanderboom et al., Reference Vanderboom, Misra, Rodino, Eberly, Greenwood, Morris, Norrie, Fernholz, Pritt and Norgan2024). Additionally, cross-priming amplification combined with vertical flow visualization strip technology serves as a rapid, accurate and nearly instrument-free molecular diagnostic method for epidemiological investigation and diagnosis of B. motasi-induced babesiosis in sheep or humans, making it suitable for resource-limited settings (Wang et al., Reference Wang, Gao, Zhang, He, Liu, Liu, Li, Liu, Luo, Guan and Yin2020). These novel technologies provide powerful tools for precise diagnosis, outbreak surveillance and blood safety screening of babesiosis.
Clinical treatment and drug resistance of babesiosis
Currently, the standard treatment for human babesiosis primarily relies on 2 classic drug combinations: atovaquone with azithromycin and quinine with clindamycin. Among these, the atovaquone-azithromycin regimen is recommended as the first-line therapy due to its high clinical efficacy and favourable safety profile (Table 5 (Krause et al., Reference Krause, Auwaerter, Bannuru, Branda, Falck-ytter, Lantos, Lavergne, Meissner, Osani, Rips, Sood, Vannier, Vaysbrot and Wormser2020)) (Weiss, Reference Weiss2002; Wormser et al., Reference Wormser, Dattwyler, Shapiro, Halperin, Steere, Klempner, Krause, Bakken, Strle, Stanek, Bockenstedt, Fish, Dumler and Nadelman2006; Chen et al., Reference Chen, Li, Gao, Bian, Yan, Kong and Liu2019). Clinical studies have demonstrated that atovaquone 40 mg kg−1 day−1 combined with azithromycin 10 mg kg−1 day−1 effectively cures neonatal babesiosis without significant adverse reactions (Aderinboye and Syed, Reference Aderinboye and Syed2010; Liu et al., Reference Liu, Li, Wang, Gao, Xu, Zhao, Liu, Chen, Zhou, Cai and He2026). Furthermore, in immunocompromised patients such as those with HIV infection, elderly individuals and IgM myeloma patients, atovaquone 750 mg twice daily plus azithromycin 500–1000 mg daily is recommended. Based on current evidence, the atovaquone-azithromycin combination is not only suitable for immunocompetent patients with mild to moderate babesiosis but also effective for severe and life-threatening disease. Atovaquone-azithromycin offers safe and effective therapy for most cases and represents the currently recommended first-line option (Krause et al., Reference Krause, Lepore, Sikand, Gadbaw, Burke, Telford, Brassard, Pearl, Azlanzadeh, Christianson, McGrath and Spielman2000; Kletsova et al., Reference Kletsova, Spitzer, Fries and Marcos2017; Chen et al., Reference Chen, Li, Gao, Bian, Yan, Kong and Liu2019).
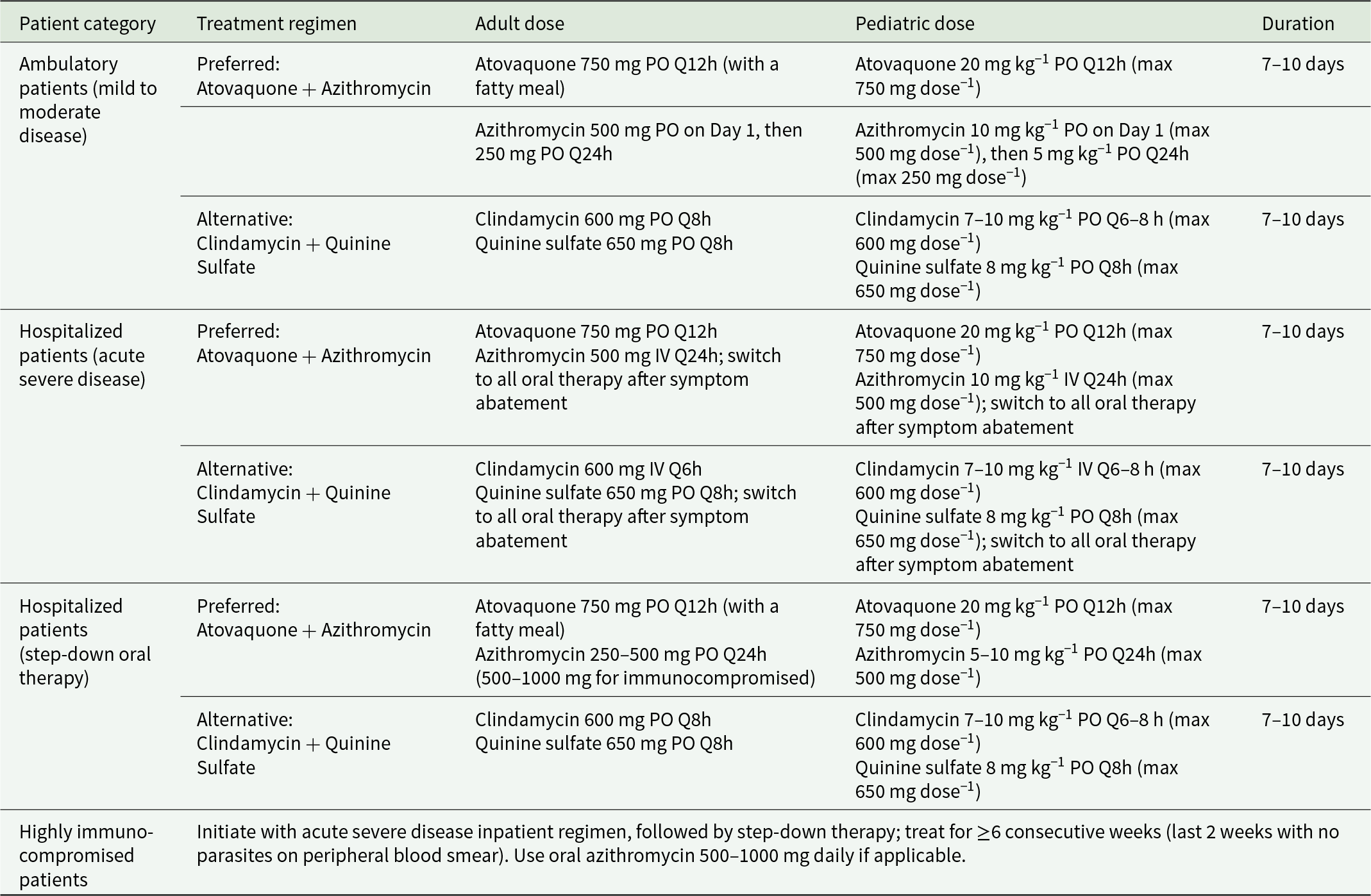
Treatment regimens for babesiosis in adults and children (Krause et al., Reference Krause, Auwaerter, Bannuru, Branda, Falck-ytter, Lantos, Lavergne, Meissner, Osani, Rips, Sood, Vannier, Vaysbrot and Wormser2020)

Table 5 Long description
The table lists recommended babesiosis drug regimens by patient category, with adult and pediatric dosing and typical treatment length. For ambulatory mild to moderate disease, the preferred regimen is atovaquone taken by mouth twice daily plus azithromycin, with a higher first-day azithromycin dose followed by a lower daily dose; an alternative is clindamycin plus quinine sulfate. For hospitalized acute severe disease, the preferred regimen keeps atovaquone by mouth twice daily but gives azithromycin intravenously once daily, then switches to all-oral therapy after symptoms improve; the alternative uses intravenous clindamycin with oral quinine sulfate, also switching to oral after improvement. Step-down oral therapy for hospitalized patients continues the same preferred or alternative drug pairs using oral dosing. Across these categories, the usual duration is seven to ten days. Pediatric doses are weight-based with stated maximum per-dose limits, while adult doses are fixed. Highly immunocompromised patients are treated longer, starting with the inpatient severe-disease regimen and then step-down therapy for at least six consecutive weeks, with the final two weeks requiring no parasites seen on blood smear.
PO = orally; IV = intravenously; Q12h = every 12 hours; Q24h = every 24 hours; Q6h = every 6 hours; Q8h = every 8 hours; max = maximum. This table refers to the Clinical Practice Guidelines by the Infectious Diseases Society of America (IDSA): 2020 Guideline on the Diagnosis and Management of Babesiosis.
The combination therapy of quinine and clindamycin was once considered the standard treatment for babesiosis (Krause et al., Reference Krause, Lepore, Sikand, Gadbaw, Burke, Telford, Brassard, Pearl, Azlanzadeh, Christianson, McGrath and Spielman2000). The standard regimen typically involves a 7- to 10-day course with quinine administered orally at 650 mg 3–4 times daily and clindamycin given orally or intravenously at 600 mg 3–4 times daily. However, in severely immunocompromised patients such as those post-splenectomy, with HIV infection, or on long-term immunosuppressive therapy, this regimen may be ineffective. Exchange transfusion is recommended as a vital adjunctive therapy administered in combination with antimicrobial agents to rapidly reduce parasite load (Krause et al., Reference Krause, Lepore, Sikand, Gadbaw, Burke, Telford, Brassard, Pearl, Azlanzadeh, Christianson, McGrath and Spielman2000; Ord and Lobo, Reference Ord and Lobo2015).
Artemisinin derivatives such as artesunate and artemether may also serve as an option for treating Babesia infections (Goo et al., Reference Goo, Terkawi, Jia, Aboge, Ooka, Nelson, Kim, Sunaga, Namikawa, Igarashi, Nishikawa and Xuan2010). Additionally, atovaquone-proguanil therapy represents another alternative (Vyas et al., Reference Vyas, Telford and Robbins2007), though its efficacy requires further validation in clinical practice.
Babesia species have developed significant resistance to commonly used therapeutic agents. In Babesia gibsoni from infected dogs, treatment with diminazene aceturate results in prolonged therapy duration and increased relapse rates, yet no resistance-associated mutations have been identified in the cytochrome b gene, suggesting that the resistance mechanism may be independent of this target (Feng et al., Reference Feng, Wu, Lin and Yao2022). Conversely, resistance to atovaquone is closely associated with specific mutations in the cytochrome b gene, such as P152L and F258Y, which occur within the drug-binding region and compromise drug efficacy (Rahman et al., Reference Rahman, Panicker, Narayanan, Pillai, Unny, Krishna and Simon2022). Additionally, B. microti can exhibit multidrug resistance to atovaquone-azithromycin in immunocompromised patients, linked to cytochrome b V141A mutations and 23S rRNA mutations (Rogers et al., Reference Rogers, Krause, Norris, Ting, Nagami, Cilley and Vannier2023; Krause et al., Reference Krause, Rogers, Shah, Kang, Parsonnet, Kodama and Vannier2024). Although drug-resistant strains maintain normal growth within mammalian erythrocytes, their fitness is reduced in tick vectors; nevertheless, they remain capable of tick-borne transmission to naive mammalian hosts, facilitating the persistence of resistance. Experimental data confirm I. scapularis can transmit drug-resistant B. microti to uninfected mice (Chiu et al., Reference Chiu, Renard, George, Pal, Alday, Narasimhan, Riscoe, Doggett and Ben Mamoun2022). Overall, the mechanisms of drug resistance in Babesia are diverse, involving target gene mutations, alterations in drug metabolic pathways and underscore the urgent need for novel therapeutics and combination therapy strategies.
Risk factors and potential public health threats of babesiosis in China
The Centers for Disease Control and Prevention USA, designated babesiosis as a nationally notifiable disease in 2011, with states asked to report both confirmed and probable cases. The American Red Cross began blood screening for Babesia-infected blood donors in 2019. In contrast, China has not yet included babesiosis in its national statutory notifiable disease system. However, China has made significant progress in diagnostic standardization through multi-province collaborative projects that have refined microscopic and nucleic acid detection standards. For instance, Zhejiang Province has established systematic surveillance of natural foci since 2020, with ongoing monitoring of reservoir hosts, tick vectors and asymptomatic infections (Wu et al., Reference Wu, Wang, Liu, Li, Luo and Gong2022). Additionally, the higher reported case counts in the USA also reflect inherent epidemiological factors: the northeastern US is a highly endemic region for Babesia, with ecological conditions including high vector density, suitable habitats and human exposure patterns that sustain high transmission intensity independent of diagnostic capacity or reporting infrastructure.
Several populations and regions in China face elevated risk. Endemic areas, including Heilongjiang, Zhejiang, Yunnan, Guangxi and Henan, form a belt-like distribution from northeast to southwest China, where suitable tick habitats and reservoir hosts exist. Farmers, forest workers and outdoor enthusiasts experience higher tick exposure. Immunocompromised individuals are particularly susceptible to severe outcomes. Blood transfusion recipients in endemic regions face additional risk due to the absence of routine nationwide screening, while patients with co-infections from other tick-borne pathogens may experience complicated diagnosis and treatment.
Babesiosis emergence in China presents several public health threats. Underdiagnosis remains pervasive without routine surveillance, resulting in substantial underestimation of true disease burden. Transfusion safety represents a critical vulnerability, as asymptomatic carriers in endemic provinces may donate blood. Climate change may further expand tick vectors into previously unaffected regions. Emerging drug resistance to first-line therapies in immunocompromised patients underscores the need for alternative treatments and resistance monitoring. Inadequate public health infrastructure, including limited laboratory capacity and insufficient healthcare worker training, hinders early detection and effective response. These challenges warrant strengthened surveillance, standardized diagnostics and integrated control strategies to address this emerging tick-borne disease threat.
Monitoring and prevention recommendations
To effectively address the threat of babesiosis, an emerging parasitic disease transmitted primarily through tick bites, China has progressively established a multi-regional surveillance system. Against this backdrop, Zhejiang Province has served as a pioneer since 2012, conducting systematic investigations and monitoring of Babesia natural foci for consecutive years, becoming one of the earliest provinces to establish provincial-level surveillance sites and contributing vital local practical experience to the improvement of the health care system in China (Ruan et al., Reference Ruan, Zhang, Chen, Lu, Zhang, Feng and Yao2017). For prevention and control, it is recommended to enhance health education by disseminating disease knowledge through community campaigns and new media platforms, targeting rural residents and frequent outdoor activity participants, while training primary healthcare workers to improve identification capabilities and avoid confusion with diseases such as malaria and Lyme disease. Livestock tick control should also be regulated to reduce transmission risks from animal hosts. To interrupt transmission pathways, 2 key measures should be strengthened: (1) Tick management and personal protection: Conduct environmental disinfection in farming and forested areas to reduce tick host density, and promote protective behaviours among high-risk groups (e.g. outdoor workers, rural residents) such as wearing long sleeve shirts and pants, tucking pants inside of socks, using tick repellents, performing tick checks after potential tick exposure and avoiding prolonged stays in tick-infested habitats; (2) blood product screening: Enforce strict screening of blood supplies in endemic regions, including mandatory epidemiological history inquiries for blood donors and laboratory testing for Babesia pathogens. Recommended laboratory methods include NAT for Babesia DNA/RNA, quantitative real-time PCR (qPCR) and nested PCR, which exhibit high sensitivity and specificity for pathogen detection. Currently, China has launched pilot blood transfusion screening programs for Babesia in endemic provinces (Ji et al., Reference Ji, Chang, Yan, Sun and Wang2024) (e.g. Xinjiang, Inner Mongolia, Heilongjiang), integrating NAT into routine blood safety testing in key regions. Although nationwide universal screening has not yet been implemented, the surveillance and screening system is continuously expanded with the support of national public health projects, aiming to mitigate transfusion-transmitted babesiosis risks. Clinically, diagnostic and treatment capacities should be elevated through the promotion of rapid diagnostic techniques, the formulation of national treatment guidelines, and the optimization of therapeutic protocols. Furthermore, surveillance and early warning systems need improvement via the establishment of a nationwide monitoring network, active surveillance in all endemic areas (16 provinces/regions: Heilongjiang, Guangxi, Henan, Yunnan, Zhejiang, Fujian, Jiangxi, Guangdong, Beijing, Xinjiang, Inner Mongolia, Chongqing, Taiwan, Shandong, Gansu, Sichuan) and potential risk areas, inclusion of babesiosis in the national notifiable disease reporting system, and enhanced molecular tracing of pathogens. Finally, increased support for basic and applied research will be essential to narrow gaps with developed countries and ensure the sustained and effective advancement of control strategies.
Conclusion
Babesiosis, an emerging tick-borne zoonotic parasitic disease, has shown a trend of global expansion in recent years. The first human babesiosis cases in China were reported by Hong Shilü in 1943. Since this first report, a total of 328 infections have been documented nationwide, involving all identified human-pathogenic Babesia species in China: B. microti, B. venatorum, B. divergens, B. crassa-like organisms, B. microti-like species and the novel B. sp. XXB/Hangzhou, with their distribution extending to 16 endemic provinces/regions across the country. Current diagnostic techniques include blood smear microscopy, serological testing, molecular methods like PCR and nested PCR and mNGS. The preferred treatment is atovaquone combined with azithromycin, although drug resistance is becoming increasingly prominent. Future efforts should focus on enhancing active surveillance of vector ticks and animal hosts, establishing rapid and accurate diagnostic systems, optimizing clinical treatment strategies and promoting public health education to address the potential threat of babesiosis to public health in China.
Data availability statement
Not applicable.
Author contributions
Y.C.: Writing – review & editing, writing – original draft, validation, methodology, investigation, resources, formal analysis, data curation, conceptualization. J.X.: Supervision, investigation, resources, data curation, conceptualization. J.S.: Investigation, resources, data curation. Y.L.: Supervision, investigation, resources, data curation. H.X.: Resources, investigation, validation. Y.C.: Writing – review & editing, supervision, validation, methodology, investigation, resources, formal analysis, data curation, conceptualization. M.C.: Writing – review & editing, supervision, validation, methodology, investigation, resources, formal analysis, data curation, conceptualization.
Financial support
This study was supported by The National Parasitic Resources Center, and the Ministry of Science and Technology fund (NPRC-2019-194-30), the Three-Year Public Health Action Plan (2023-2025) of Shanghai (No. GWVI-11.2-XD33), Shenzhen Major Project for Medical Research (B2404002), the National Science and Technology Basic Resources Survey Program (2024FY100903), the Key Research and Development Program in Hainan Province (Grant No. ZDYF2024SHFZ083), the Hainan Province Health Technology Innovation Joint Project (No. WSJK2024MS226) and Shanghai Oriental Talent Program Top-Notch Project (No. BJWS2025006).
Competing interests
None of the authors has conflicts of interest to report.
Ethical standards
Not applicable.


 Open access
Open access