



Introduction
Malaria remains one of the major public health challenges affecting mostly sub-Saharan Africa, with Plasmodium falciparum (P. falciparum) being the main species involved. In 2024, approximately 265 million cases of malaria and 569 000 deaths were reported worldwide, with the African region accounting for 95% of cases and 94% of deaths, with 75% of deaths occurring in children under 5 years of age (WHO, 2025). In the Republic of the Congo, where malaria is endemic with perennial transmission, 46% of consultations, 33% of hospitalizations and 22% of deaths due to malaria were reported in 2023, with P. falciparum being the main species responsible for the disease (PNLP, 2024, unpublished data).
In the absence of an effective malaria vaccine, a timely and accurate diagnosis of malaria is crucial for malaria case management and control programmes. Since 2010, the World Health Organization (WHO) has recommended that malaria be diagnosed before any treatment is administered (WHO, 2010). In 2024, in the Republic of the Congo, 62% of malaria cases were diagnosed by rapid diagnostic tests (RDTs), while 38% were diagnosed by microscopy (PNLP, 2024). The type of RDT commonly used is the P. falciparum malaria antigen RDT, with histidine-rich protein 2 (HRP2) as the targeted antigen (PNLP, 2024). Although microscopy is the widely used reference diagnostic test, RDTs play a crucial role in patient management, particularly in remote or resource-limited areas where access to microscopy is limited (WHO, 2023). HRP2-based RDTs are lateral flow immunochromatographic tests that detect HRP2 antigens, a protein specifically secreted by P. falciparum and which has a structural homology with HRP3 protein. Thus, cross-reactivity of anti-HRP2 antibodies with HRP3 may occur when using these tests (Kong et al., Reference Kong, Wilson, Ah, Nace, Rogier and Aidoo2021). Due to their operational simplicity, stability at room temperature and minimal requirements for specialized equipment or advanced technical training, RDTs are an essential tool for improving case management and contribute to strengthening malaria surveillance (Oyegoke et al., Reference Oyegoke, Akoniyon, Maharaj, Adewumi, Malgwi, Aderoju, Fatoba, Adeleke, Maharaj and Okpeku2024). HRP2-based RDT performance can be influenced by several factors, such as antigenic diversity of proteins, persistence of HRP2 in the circulation after parasite clearance and parasite density (Mayxay et al., Reference Mayxay, Pukrittayakamee, Chotivanich, Looareesuwan and White2001). In addition, the emergence of deletions in the Pfhrp2 and/or Pfhrp3 genes reported in several countries, such as Eritrea (9.4 and 41.7%) and the Democratic Republic of the Congo (6.4%), poses a serious threat to malaria control and elimination. These deletions cause false-negative RDT results (Parr et al., Reference Parr, Verity, Doctor, Janko, Carey-ewend, Turman, Keeler, Slater, Whitesell, Mwandagalirwa, Ghani, Likwela, Tshefu, Emch, Juliano and Meshnick2017; Mihreteab et al., Reference Mihreteab, Anderson, Pasay, Smith, Gatton, Cunningham, Berhane and Cheng2021; Rogier et al., Reference Rogier, Battle, Bakari, Seth, Nace, Herman, Barakoti, Madebe, Mandara, Lyimo, Giesbrecht, Popkin-Hall, Francis, Mbwambo, Garimo, Aaron, Lusasi, Molteni, Njau and Ishengoma2024). Therefore, current genomic surveillance has focused on the surveillance of deletions of the Pfhrp2 and Pfhrp3 genes in symptomatic individuals, as recommended by the WHO (WHO, 2019, 2023). In 2023, a multiregional study including P. falciparum isolates from Gabon, the Republic of the Congo, Nigeria and Benin revealed a low prevalence of deletions in the Pfhrp2 and Pfhrp3 genes in Gabon and Nigeria, with a few double deletions of Pfhrp2/3. No deletions in the Pfhrp2 and Pfhrp3 genes were found in isolates from the Republic of the Congo and Benin. However, in the Republic of the Congo, the data obtained covered only Brazzaville, the capital of the country, and Goma Tsé-Tsé, a district of the Pool department in the south of the country (Krueger et al., Reference Krueger, Ikegbunam, Lissom, Sandri, Ntabi, Djontu, Baina, Lontchi, Maloum, Ella, Agonhossou, Akoton, Djogbenou, Borrmann, Held, Ntoumi, Adegnika, Kremsner and Kreidenweiss2023). To the best of our knowledge, no information on deletions in the Pfhrp2 and Pfhrp3 genes is available for other parts of the country. The purpose of our study was to determine the prevalence of P. falciparum deletions in the Pfhrp2 and Pfhrp3 genes in isolates from symptomatic individuals in the Mossendjo locality.
Materials and methods
Study site and design
This study was conducted in 2 health centres, the Centre de Santé Intégré-Poste (CSIMSJ) and the Hôpital de Base (HBMSJ), located in Mossendjo, Niari Department.
This study is a cross-sectional investigation of Pfhrp2/3 deletions, which is part of a larger research study that aimed to evaluate the performance of routine microscopy for malaria diagnosis, to determine the prevalence of symptomatic infection and to carry out molecular characterization of P. falciparum isolates. In this remote area, details on the level of malaria transmission and RDT test pressure/treatment pressure were not documented. The study research protocol was approved by the ‘Comité d’Ethique de la Recherche en Sciences de la Santé’ (N◦275/MRSIT/IRSSA/CERSSA-2021) of the Republic of the Congo.
Study population, blood samples and data collection
All patients attending one of the study sites with signs of uncomplicated malaria from January to December 2022 were invited to participate in this study. Those who showed signs of severe malaria or any other pathologies, according to the doctor, as well as pregnant women, were not enrolled. Whole blood (2–5 mL) was collected in Vacutainer tubes containing ethylenediaminetetraacetic acid. For each patient, a few drops were placed on Whatman 3 MM filter paper, which was then placed in separate zip-lock bags with a desiccant at room temperature. As previously described, the sample size was estimated using the Schwartz method based on the malaria proportion of 18.8%, corresponding to the proportion of suspected malaria case confirmed by microscopy observed 1 year prior to study implementation at the participating health facilities, with a 95% confidence level and a 5% margin of error. The minimum required sample size was then allocated proportionally between the 2 study sites based on the number of suspected malaria cases, which was 2.78 times higher at CSIMSJ than at HBMSJ, resulting in a minimum sample size of 650 and 234 (Fila-Fila et al., Reference Fila-Fila, Koukouikila-Koussounda, Niama, Bissombolo Madingou, Demboux, Mandiangou, Vembe Mahounga, Doniama, Dossou-Yovo, Casimiro and Issamou Mayengue2024).
Microscopic analysis
Blood smears were prepared according to the WHO protocol for blood smear preparation and subsequent Giemsa staining (WHO, 2016). The slides were stained with 10% Giemsa working solution for 15 min, gently rinsed with tap water and then dried before examination. The slide was considered negative when approximately 200 microscopic fields were scanned and no parasite was found. Slides with detected parasites were evaluated until 200 leukocytes had been counted. The parasite density was defined as the number of parasites/μL of blood, considering the leukocyte count of 8000 μL−1 of blood according to the WHO method. The slides were read by 2 experienced microscopists. In the event of conflicting results, a third reader was called in to reread the slides. The microscopic results were immediately reported for possible patient care.
Identification of P. falciparum positive samples by PCR
Genomic DNA was extracted from the dried blood spots using the Zymo Quick kit (Zymo Research, Irvine, CA, USA) according to the manufacturer’s instructions. The identification of P. falciparum positive samples was performed using a nested PCR technique using primers targeting polymorphic genes of msp1 and msp2, as described by Fila-Fila et al. (Reference Fila-Fila, Koukouikila-Koussounda, Niama, Bissombolo Madingou, Demboux, Mandiangou, Vembe Mahounga, Doniama, Dossou-Yovo, Casimiro and Issamou Mayengue2024). The laboratory strains 3D7 and Dd2 were used as positive PCR controls, and DNA was extracted from 200 µL of erythrocyte suspension from 5% parasitaemia of either 3D7 or Dd2 culture. Deionized water was used as a negative control.
Genotyping of the Pfhrp2 and Pfhrp3 genes
P. falciparum positive samples were used for genotyping of Pfhrp2 and Pfhrp3 genes. We performed nested PCRs using primers that covered exon 1–2 of each gene (Supplementary Table S1) as previously described by Abdallah et al. (Reference Abdallah, Okoth, Fontecha, Torres, Banegas, Matute, Bucheli, Goldman, de Oliveira, Barnwell and Udhayakumar2015). Briefly, for each gene, the primary PCR was prepared in a total volume of 25 μL, containing 12.5 μL of Platinum™ Hot Start Green PCR Master Mix, 400 nM of the forward and reverse primers, 5.5 μL of deionized water and 5 μL of DNA template. The DNA reference strain 3D7 was used as a positive wild-type control for both genes. Furthermore, the Dd2 strain served as a positive wild-type control for Pfhrp3 gene but was mutant-type for Pfhrp2. Conversely, HB3 strain served as a positive wild-type control for Pfhrp2 and mutant-type for Pfhrp3. The parasite density of each reference strain was 5%. The PCR programme was followed: 95 °C/5 min − [95 °C/30 s; X/30 s; 68 °C/30 s] × 30 cycles − 68 °C/5 min, with X being the annealing temperature of 55 °C and 53 °C for Pfhrp2 and Pfhrp3, respectively. Both nested PCR reactions were conducted using the primary protocol, with an annealing temperature of 62 °C. For all nested PCR products, 5 μL were loaded onto a 2% agarose gel (PeqLab, Erlangen, Germany), stained with ethidium bromide, separated by electrophoresis and visualized under ultraviolet transillumination.
Nested PCRs were repeated twice for all negative samples. The persisting negative samples were then amplified using the ‘4-plex qPCR’ protocol, which was described by Kreidenweiss et al. (Reference Kreidenweiss, Trauner, Rodi, Koehne, Held, Wyndorps, Manouana, McCall, Adegnika, Lalremruata, Kremsner, Fendel and Sandri2019). This PCR method simultaneously amplifies 4 genes: P. falciparum ß tubulin (Pfbtub), P. falciparum cytochrome b (Pfcytb), Pfhrp2 and Pfhrp3. The Pfbtub and Pfcytb genes are considered internal controls. The Pfbtub is a single-copy gene, which was used to assess DNA extraction quality, while Pfcytb, which is a mitochondrial gene, was used to confirm the presence of parasite. When both controls and the genes of interest (Pfhrp2 and/or Pfhrp3) were amplified, the result was interpreted as the absence of deletion. In contrast, the absence of amplification of the Pfhrp2 and/or Pfhrp3 gene while both controls were amplified suggested the presence of Pfhrp2 and/or Pfhrp3 deletion, as described by Krueger et al. (Reference Krueger, Ikegbunam, Lissom, Sandri, Ntabi, Djontu, Baina, Lontchi, Maloum, Ella, Agonhossou, Akoton, Djogbenou, Borrmann, Held, Ntoumi, Adegnika, Kremsner and Kreidenweiss2023).
We used a TaqMan Multiplex Master Mix (Thermo Fisher Scientific, Waltham, MA, USA) at a concentration of 400 nM for forward and reverse primers of Pfcytb, Pfhrp 2 and Pfhrp 3 and 600 nM for primers of Pfbtub (Supplementary Table S2). The final probe concentrations were 300 nM for Pfbtub, 75 nM for Pfhrp2 and 150 nM for Pfhrp3 and Pfcytb, with a final volume of 10 µL including 3 µL template DNA. The amplifications were carried out under the following PCR conditions: Initial activation at 95 °C for 20 s, followed by 45 cycles of denaturation at 95 °C for 3 min, annealing at 62 °C for 50 s and a final cooling step at 40 °C. We define a case of deletion of Pfhrp2 and Pfhrp3 as a sample that was negative in the nested PCRs and the quadruplex real-time PCR of Pfhrp2/3, but positive in one or both of the msp 1 and msp 2 genes.
Results
Characteristics of the study participants
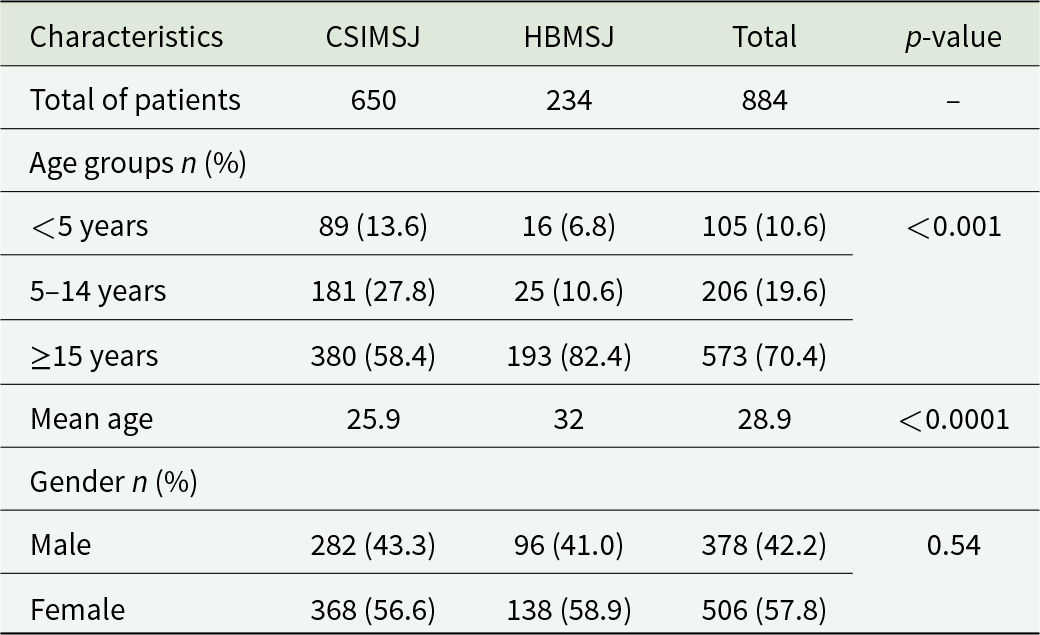
A total of 884 patients of all ages participated in this study, with 650 and 234 recruited at CSIMSJ and HBMSJ, respectively. The patients’ ages ranged from 6 months to 85 years and from 4 months to 80 years, with a mean age of 25.9 and 32.0 years in CSIMSJ and HBMSJ, respectively. Women were more represented than men at both sites, accounting for 56.6% of patients at CSIMSJ and 58.9% at HBMSJ. Children under 5 years of age were the least represented, with prevalences of 13.6 and 6.8% at CSIMSJ and HBMSJ, respectively (Table 1). Considering the 2 sites together, 41.6% (368/884) of patients tested positive by microscopy, including 44.4% (289/650) and 33.7% (79/234) at CSIMSJ and HBMSJ, respectively. The geometric mean parasite densities were 4845.5 parasites µL−1 [95% CI: 4201.8–5489.2] at CSIMSJ and 4019.8 parasites µL−1 [95% CI: 3082.9–4956.7] at HBMSJ, respectively.
Sociodemographic and clinical characteristics of the study participants

Table 1 Long description
The table summarizes participant counts, age distribution, mean age, and gender for two study sites (CSIMSJ and HBMSJ) and overall totals, with p-values for site comparisons. There are 650 patients at CSIMSJ and 234 at HBMSJ, for 884 total. Age differs notably by site: at CSIMSJ, 13.6 percent are under 5 years, 27.8 percent are 5 to 14 years, and 58.4 percent are 15 years or older; at HBMSJ, 6.8 percent are under 5 years, 10.6 percent are 5 to 14 years, and 82.4 percent are 15 years or older. Overall, 70.4 percent of participants are 15 years or older. Mean age is higher at HBMSJ (32) than at CSIMSJ (25.9), and the age comparison is statistically significant (p less than 0.001). Gender distribution is similar across sites: males are 43.3 percent at CSIMSJ and 41.0 percent at HBMSJ, while females are 56.6 percent at CSIMSJ and 58.9 percent at HBMSJ, with no statistically significant difference (p 0.54). Percentages may not sum to exactly 100 due to rounding.
The average age at HBMSJ is significantly higher than that at CSIMSJ (difference ≈ 6 years, p < 0.001, 95% CI: [6.2–5.8]). For gender, the analysis showed no statistically significant difference in gender distribution at the 2 sites (p = 0.54; OR: 1.10; 95% CI: [0.80–1.51]).
CI, confidence interval.
Genotyping of Pfhrp2 and Pfhrp3
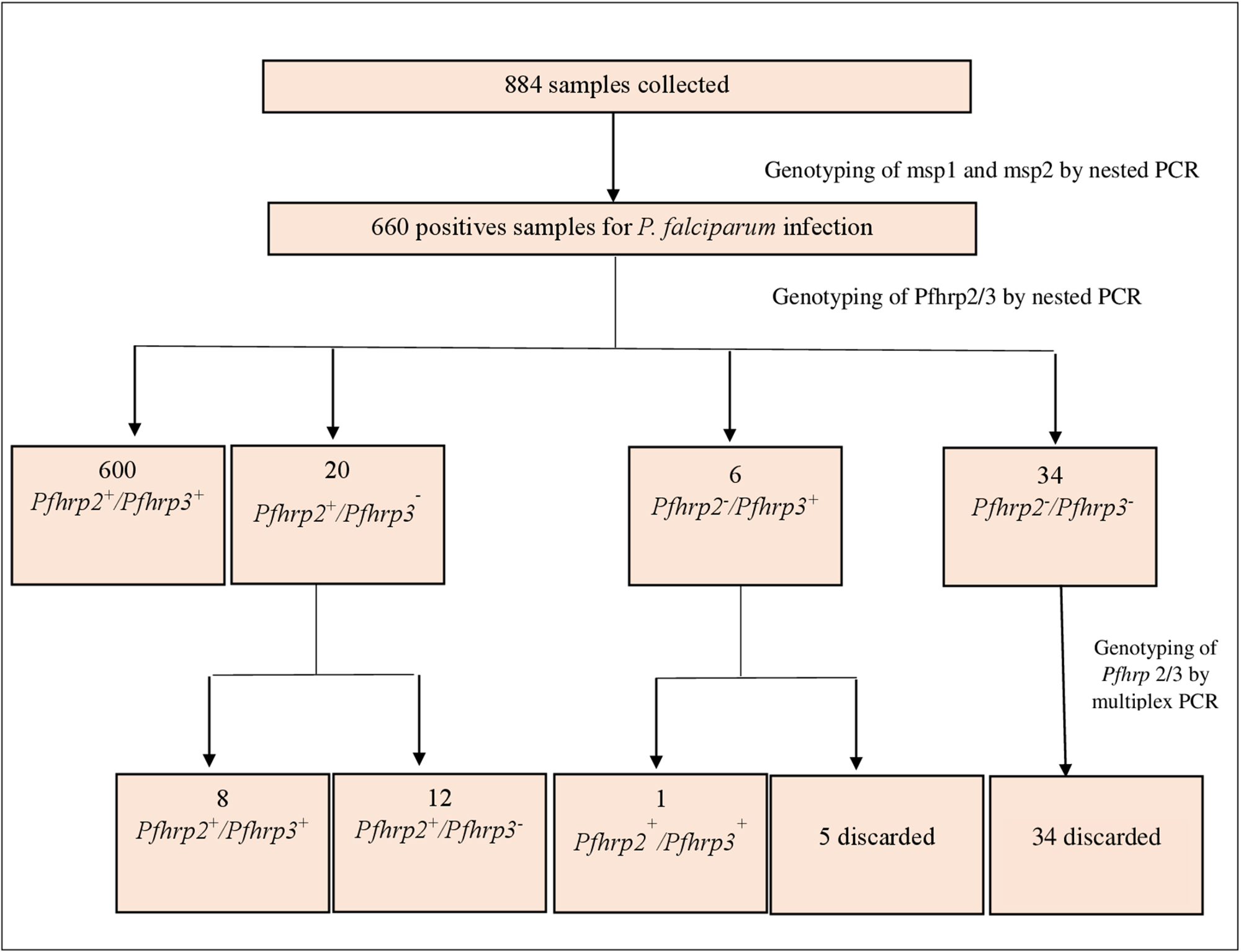
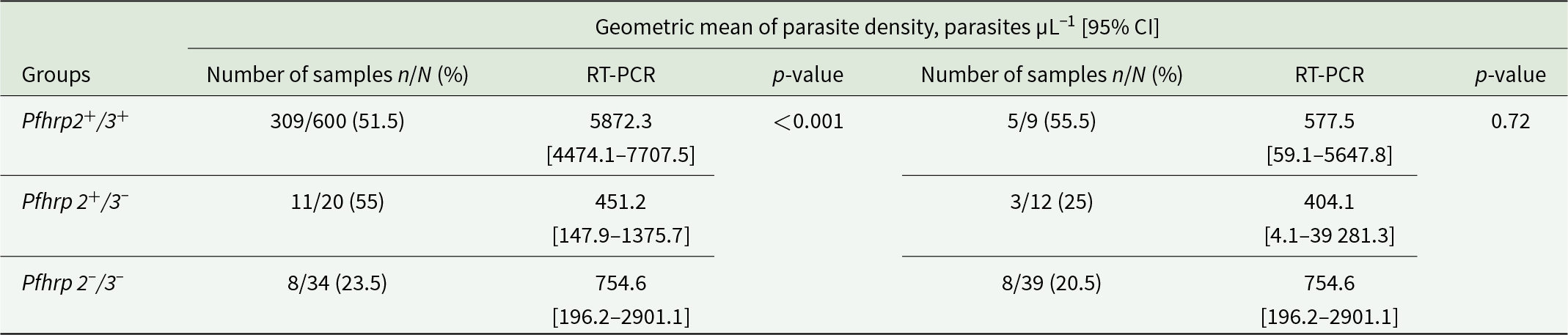
Of the 884 samples analysed by nested PCR for the msp1 and msp2 genes, 660 (74.7%) were positive, of which 316 (47.9%) corresponded to microscopy-negative infections. PCR-positive samples were then genotyped for Pfhrp2 and Pfhrp3 by nested PCR using specific primers. Of them, 600 (90.9%) tested positive for the Pfhrp2 and Pfhrp3 genes, and 60 (9.1%) were negative for at least 1 gene (Figure 1). To confirm the Pfhrp2/3 negativity of these samples, the quadruplex real-time PCR was performed using the Pfcytb, Pfbtub, Pfhrp2 and Pfhrp3 genes. Following analysis, the results from 39 samples (65%) were discarded due to DNA quality, 9 (15%) samples tested positive for the Pfhrp2 and Pfhrp3 genes, while 12 (20%) samples tested positive for the Pfhrp2 gene and negative for the Pfhrp3 gene, of which 3 samples were microscopically positive. No double deletion of Pfhrp2 and Pfhrp3 was found. The Pfhrp2+/3+ group identified by nested PCR had a higher mean geometric parasite density of 5872.3 parasites µL−1 [95% CI: 4474.1–7707.5] than Pfhrp 2+/3− group with 451.2 parasites µL−1 [95% CI: 147.9–1375.7] and Pfhrp 2−/3− group with 754.6 parasites µL−1 [95% CI: 196.2–2901.1] (p < 0.001). In contrast, no significant difference was observed between the same groups using RT-PCR (p = 0.72) (Table 2).
PCR analysis workflow.

Figure 1 Long description
The flowchart illustrates the genotyping process of 884 collected samples. Initially, genotyping of msp1 and msp2 is performed by nested PCR, resulting in 660 positive samples for P. falciparum infection. These samples undergo further genotyping for Pfhrp2 and Pfhrp3 by nested PCR. The results are divided into four groups: 600 samples are Pfhrp2 plus/Pfhrp3 plus, 20 samples are Pfhrp2 plus/Pfhrp3 minus, 6 samples are Pfhrp2 minus/Pfhrp3 plus and 34 samples are Pfhrp2 minus/Pfhrp3 minus. Further genotyping of Pfhrp2/3 by multiplex PCR is conducted. From the 600 samples, 8 are Pfhrp2 plus/Pfhrp3 plus and 12 are Pfhrp2 plus/Pfhrp3 minus. From the 6 samples, 1 is Pfhrp2 minus/Pfhrp3 plus and 5 are discarded. From the 34 samples, all are discarded.
Relationship between parasite density, PCR type and deletions in the Pfhrp 2 and Pfhrp 3 genes

Table 2 Long description
The table compares geometric mean parasite density measured by RT-PCR across three Pfhrp2 and Pfhrp3 deletion groups, reported with 95 percent confidence intervals and sample counts. For Pfhrp2 and Pfhrp3 intact, microscopy-positive samples are 309 of 600 and the RT-PCR geometric mean is 5872.3 parasites per microliter with a confidence interval from 4474.1 to 7707.5; the paired smaller sample set is 5 of 9 with a mean of 577.5 and a confidence interval from 59.1 to 5647.8. For Pfhrp2 intact and Pfhrp3 deleted, microscopy-positive samples are 11 of 20 with a mean of 451.2 and a confidence interval from 147.9 to 1375.7; the paired set is 3 of 12 with a mean of 404.1 and a confidence interval from 4.1 to 39281.3. For Pfhrp2 and Pfhrp3 deleted, microscopy-positive samples are 8 of 34 with a mean of 754.6 and a confidence interval from 196.2 to 2901.1; the paired set is 8 of 39 with the same mean and confidence interval. The only reported statistical test shows a significant difference for the Pfhrp2 and Pfhrp3 intact group in the first comparison, while the paired comparison for that group is not significant. Confidence intervals are wide in the smaller groups, so differences should be interpreted cautiously due to limited sample sizes and imprecision.
n, microscopy positive samples in each group; N, total number of samples in each group.
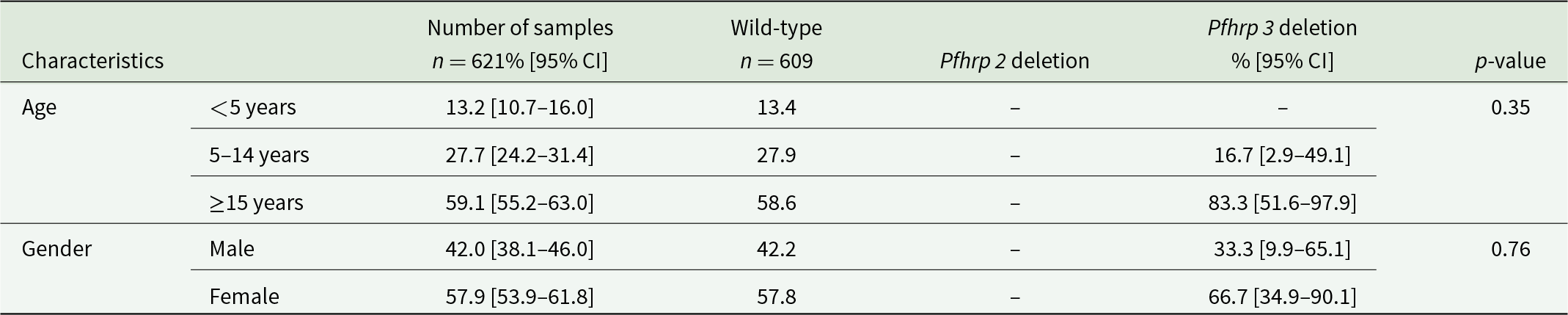
All children under the age of 5 years had wild-type profiles for the Pfhrp2 and Pfhrp3 genes. Among the 12 samples presenting the deletion of the Pfhrp3, 83.3% were from adults and 16.7% from children (p = 0.35), while 66.7% were from women compared to 33.3% from men (p = 0.76) (Table 3). In addition, 9 of the 12 samples with Pfhrp3 deletion (75%) were microscopy negative infections.
Relationship between age, gender and the deletion in the Pfhrp 2 and Pfhrp 3 genes

Table 3 Long description
The table compares the distribution of samples by age group and gender across wild-type parasites and those with Pfhrp2 or Pfhrp3 gene deletions, reporting percentages with 95 percent confidence intervals and p-values. There were 621 samples overall and 609 wild-type samples; Pfhrp2 deletion values are not reported in the table. By age, the overall sample was 13.2 percent under 5 years, 27.7 percent ages 5 to 14, and 59.1 percent ages 15 years or older; wild-type percentages were similar at 13.4, 27.9, and 58.6 percent. Pfhrp3 deletions were concentrated in the oldest group at 83.3 percent, with 16.7 percent in ages 5 to 14 and none reported under 5 years. The age comparison shows no statistically significant difference, with a p-value of 0.35. By gender, the overall sample was 42.0 percent male and 57.9 percent female, closely matching wild-type at 42.2 percent male and 57.8 percent female. Pfhrp3 deletions were 33.3 percent male and 66.7 percent female, and the gender comparison was not statistically significant with a p-value of 0.76. Confidence intervals for Pfhrp3 deletion percentages are wide, so estimates for that group are imprecise and should be interpreted cautiously.
Discussion
Studies carried out in Brazzaville, Pointe-Noire and Mossendjo on the usefulness of malaria diagnostic tools and the evaluation of microscopists have reported that (1) the diagnostic accuracy of microscopy was poor and varied from one centre to another, with sensitivities ranging from 32.9% to 94.2% and specificities ranging from 33.6% to 94.9% (Mayengue et al., Reference Mayengue, Batsimba, Dossou-Yovo, Niama, Macosso, Singana, Louzolo, Loukabou, Obili, Kobawila and Parra2018; Fila-Fila et al., Reference Fila-Fila, Koukouikila-Koussounda, Niama, Bissombolo Madingou, Demboux, Mandiangou, Vembe Mahounga, Doniama, Dossou-Yovo, Casimiro and Issamou Mayengue2024), and (2) the low use of RDTs (Ntoumi et al., Reference Ntoumi, Vouvoungui, Ibara, Landry and Sidibé2013). These observations suggest the need for targeted training of health workers in the use of diagnostic tools, regular evaluation of diagnostic performance, improved acceptability, increased availability and more widespread use of RDTs (Ntoumi et al., Reference Ntoumi, Vouvoungui, Ibara, Landry and Sidibé2013; Mayengue et al., Reference Mayengue, Batsimba, Dossou-Yovo, Niama, Macosso, Singana, Louzolo, Loukabou, Obili, Kobawila and Parra2018; Fila-Fila et al., Reference Fila-Fila, Koukouikila-Koussounda, Niama, Bissombolo Madingou, Demboux, Mandiangou, Vembe Mahounga, Doniama, Dossou-Yovo, Casimiro and Issamou Mayengue2024).
HRP2-based RDTs are commonly used with a significant impact on disease management, especially in endemic areas. However, several studies around the world have reported deletions in the Pfhrp2 and/or Pfhrp3 genes, which lead to parasites not being detected by RDT (Parr et al., Reference Parr, Verity, Doctor, Janko, Carey-ewend, Turman, Keeler, Slater, Whitesell, Mwandagalirwa, Ghani, Likwela, Tshefu, Emch, Juliano and Meshnick2017; Iriart et al., Reference Iriart, Menard, Chauvin, Mohamed, Charpentier, Mohamed, Berry and Aboubaker2020; Prosser et al., Reference Prosser, Gresty, Ellis, Meyer, Anderson, Lee and Cheng2021; Bally et al., Reference Bally, da S, Arregue, de Mattos MK and Suárez-Mutis2024; Mekonnen et al., Reference Mekonnen, Dugassa, Feleke, Taddesew, Tegegn, Adamu, Mandefro, Tasew and Golassa2024). The present study aimed to assess the prevalence of deletion in the Pfhrp2 and Pfhrp3 genes in isolates collected from patients attending 2 health facilities in Mossendjo, the Republic of the Congo, where high prevalence of microscopic and microscopy-negative infections were found (Fila-Fila et al., Reference Fila-Fila, Koukouikila-Koussounda, Niama, Bissombolo Madingou, Demboux, Mandiangou, Vembe Mahounga, Doniama, Dossou-Yovo, Casimiro and Issamou Mayengue2024).
In our study, about half of the infections were missed by microscopy in this study, Pfhrp2 deletions were absent and Pfhrp3 deletions were rare (1.9%). A multicentre study conducted in 2023 reported a low prevalence of Pfhrp2 and Pfhrp3 gene deletions in Nigeria (Pfhrp2 0.03% and Pfhrp3 0.03%), with no Pfhrp2 deletion in Gabon (Pfhrp2 1% and Pfhrp3 0%). In contrast, no Pfhrp2 or Pfhrp3 gene deletions were observed in samples from Benin, as well as from Brazzaville and its surroundings (Krueger et al., Reference Krueger, Ikegbunam, Lissom, Sandri, Ntabi, Djontu, Baina, Lontchi, Maloum, Ella, Agonhossou, Akoton, Djogbenou, Borrmann, Held, Ntoumi, Adegnika, Kremsner and Kreidenweiss2023). However, in the present study, although no Pfhrp2 deletions were detected, a 1.9% prevalence of Pfhrp3 deletions was observed. Surprisingly, the deletion of the Pfhrp2 gene was found at a prevalence higher than 5% in 2 cities located in the western part of the Democratic Republic of the Congo, a very close neighbouring country with the Republic of the Congo (Parr et al., Reference Parr, Verity, Doctor, Janko, Carey-ewend, Turman, Keeler, Slater, Whitesell, Mwandagalirwa, Ghani, Likwela, Tshefu, Emch, Juliano and Meshnick2017; Munyeku et al., Reference Munyeku, Musaka, Ernest, Smith, Mansiangi and Culleton2021). Other countries in East Africa have reported a high prevalence of deletions for the Pfhrp2 and/or Pfhrp3 genes as well as the double deletions, including Tanzania (Pfhrp2 1.6% and Pfhrp3 50%), Ethiopia (Pfhrp2 27.3%, Pfhrp3 30.5% and Pfhrp2/3 13.2%) and Eritrea (Pfhrp2 9.4%, Pfhrp3 41.7% and Pfhrp2/3 7.6%) (Mihreteab et al., Reference Mihreteab, Anderson, Pasay, Smith, Gatton, Cunningham, Berhane and Cheng2021; Kaaya et al., Reference Kaaya, Kavishe, Tenu, Matowo, Mosha, Drakeley, Sutherland and Beshir2022; Mekonnen et al., Reference Mekonnen, Dugassa, Feleke, Taddesew, Tegegn, Adamu, Mandefro, Tasew and Golassa2024; Molina-de la Fuente et al., Reference Molina-de la Fuente, Pacheco, García, González, Riloha, Oki, Benito, Escalante and Berzosa2024). Variations in the prevalence of deletions within or between countries may be due to several factors, such as the clinical status of study participants (Molina-de la Fuente et al., Reference Molina-de la Fuente, Pacheco, García, González, Riloha, Oki, Benito, Escalante and Berzosa2024), the level of disease transmission, the interaction between the deleted parasite, the host and the vector (Faik et al., Reference Faik, De carvalho and Kun2009; Mekonnen et al., Reference Mekonnen, Dugassa, Feleke, Taddesew, Tegegn, Adamu, Mandefro, Tasew and Golassa2024), as well as the frequency of use of HRP-based RDTs (Parr et al., Reference Parr, Verity, Doctor, Janko, Carey-ewend, Turman, Keeler, Slater, Whitesell, Mwandagalirwa, Ghani, Likwela, Tshefu, Emch, Juliano and Meshnick2017) and the molecular surveillance methodology adopted (Kreidenweiss et al., Reference Kreidenweiss, Trauner, Rodi, Koehne, Held, Wyndorps, Manouana, McCall, Adegnika, Lalremruata, Kremsner, Fendel and Sandri2019; Kaaya et al., Reference Kaaya, Kavishe, Tenu, Matowo, Mosha, Drakeley, Sutherland and Beshir2022). Indeed, it is expected a stronger selection pressure driven by an extended RDT use and properly implementation of ‘test-treat strategies’ (Krueger et al., Reference Krueger, Ikegbunam, Lissom, Sandri, Ntabi, Djontu, Baina, Lontchi, Maloum, Ella, Agonhossou, Akoton, Djogbenou, Borrmann, Held, Ntoumi, Adegnika, Kremsner and Kreidenweiss2023; Molina-de la Fuente et al., Reference Molina-de la Fuente, Pacheco, García, González, Riloha, Oki, Benito, Escalante and Berzosa2024). Furthermore, the exclusive use of nested PCR or conventional PCR to confirm the deletion tends to overestimate the prevalence of deletions compared to real-time PCR (Krueger et al., Reference Krueger, Ikegbunam, Lissom, Sandri, Ntabi, Djontu, Baina, Lontchi, Maloum, Ella, Agonhossou, Akoton, Djogbenou, Borrmann, Held, Ntoumi, Adegnika, Kremsner and Kreidenweiss2023; Agaba et al., Reference Agaba, Smith, Travis, Pasay, Nabatanzi, Arinaitwe, Ssewanyana, Nabadda, Cunningham, Kamya and Cheng2024). Consequently, the amplification of a single-copy gene by real-time PCR is recommended for correct diagnosis (Cheng et al., Reference Cheng, Gatton, Barnwell, Chiodini, McCarthy, Bell and Cunningham2014; Iriart et al., Reference Iriart, Menard, Chauvin, Mohamed, Charpentier, Mohamed, Berry and Aboubaker2020). Consequently, the amplification of a single-copy gene by real-time PCR is recommended for correct diagnosis (Cheng et al., Reference Cheng, Gatton, Barnwell, Chiodini, McCarthy, Bell and Cunningham2014; Iriart et al., Reference Iriart, Menard, Chauvin, Mohamed, Charpentier, Mohamed, Berry and Aboubaker2020).
The absence of deletion in the Pfhrp2 gene in this locality may suggest that the risk of obtaining false-negative RDT tests due to this deletion is low even though the non-amplification of exon 2 alone accordingly to the methodology adopted could underestimate the detection of deletions. Our data indicate that HRP2-based RDTs can still be used reliably in Mossendjo, where Pfhrp2 deletions were absent. This could improve malaria diagnosis, as microscopic accuracy has proven to be low. Further data on molecular analysis of exon 2 alone in both genes and an evaluation of the sensitivity and specificity of RDT tests in this locality are needed. Reliable RDT would allow for more effective patient care by avoiding the inappropriate use of antimalarial treatments, thereby helping to reduce drug pressure and, consequently, the emergence of Plasmodium strains resistant to antimalarials. Therefore, the potential emergence of P. falciparum strains that are undetectable by HRP2-based RDTs should be closely monitored.
Nine out of 12 samples carrying deletions of the Pfhrp3 gene were microscopy negative infections of P. falciparum, the remaining 3 samples being microscopic infections with relatively low parasite densities. A study conducted by Mekonnen et al. reported that there was no statistically significant association between parasite density and gene deletions (Mekonnen et al., Reference Mekonnen, Dugassa, Feleke, Taddesew, Tegegn, Adamu, Mandefro, Tasew and Golassa2024). This discrepancy may be due to the different molecular techniques used. Although low parasite density may reduce the sensitivity of the molecular techniques, in the current study, the RT-PCR allowing better detection of minority strains (Schindler et al., Reference Schindler, Deal, Fink, Guirou, Moser, Mwakasungula, Mihayo, Jongo, Chaki, Abdulla, Valverde, Torres, Bijeri, Silva, Hoffman, Gamboa, Tanner and Daubenberger2019) was added to the nested PCR. Pfhrp3 deletions were more prevalent in adults compared to young children between 5 and 14 years of age and absent in those under 5 years of age, probably related to the low parasite density found in the adult group (unpublished data), which may help the detection of minor strains. Indeed, low parasitaemia can facilitate the detection of minority mutant strains, which may be masked during PCR by often predominant wild strains. However, due to the low number of isolates with deletions, further studies with more isolates having deletions are needed to really assess this association.
Study limitation
The limitations of our study lie in the fact that we did not perform RDT tests. This means that the sensitivity of RDTs was not taken into account here, which would have provided additional information on the actual performance of RDTs in this context. The other limitation is that the exon 2 alone, which constitutes the main coding and antigenic region of the Pfhrp2 and Pfhrp3 genes (Baker et al., Reference Baker, Ho, Pelecanos, Gatton, Chen, Abdullah, Albertini, Ariey, Barnwell, Bell, Cunningham, Djalle, Echeverry, Gamboa, Hii, Kyaw, Luchavez, Membi, Menard and Cheng2010), was not used during amplification, while its specificity has already been demonstrated (Parr et al., Reference Parr, Anderson, Juliano and Meshnick2018). The use of exon 1–2 targeting a longer fragment could reduce PCR efficiency and increase the risk of false negatives (Woodrow and Fanello, Reference Woodrow and Fanello2017). Also, the link between low parasite density and Pfhrp3 deletions could indicate a low sensitivity of the Pfhrp3 PCRs. In addition, the PCR controls used had a relatively high parasite level (5%), which may not accurately reflect the test’s amplification performance in field samples with low parasite densities. Furthermore, the limit of detection for the in-house PCR protocol has not been formally evaluated and should be assessed in future studies.
In conclusion, this study revealed a low prevalence of Pfhrp3 deletions and no deletion in the Pfhrp2 gene. Therefore, HRP2-based RDTs can still be reliably used in Mossendjo. This could improve malaria diagnosis, as microscopic accuracy has proven to be low. However, evaluation of their performance and regular monitoring of these genes remain necessary, as recommended by the WHO.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182026102352.
Acknowledgements
We thank all the health professionals at CSIMSJ and HBMSJ for their help in collecting sociodemographic data and whole-blood venous samples. We also express our gratitude to all participants for their participation in this study and for allowing us to collect the necessary samples for this research.
Author contributions
P.I.M. and R.F.N. conceived and planned the project. P.I.M. and G.P.U.F.F. investigated and validated the methodology. P.I.M., G.P.U.F.F. and J.E.D. performed formal analysis. P.I.M. provided all resources. P.I.M., J.E.D.L. and G.P.U.F.F. conceived and analysed all data. P.I.M. and G.P.U.F.F. wrote original draft. F.R.N., F.K.K., A.F.M., J.E.D., R.G.E. and L.P.B.M. reviewed and edited the draft. P.I.M. visualized and supervised project administration. P.I.M. and F.R.N. provided funding acquisition. All authors have read and agreed to the published version of the manuscript.
Financial support
This research was supported by the Laboratoire National de Santé Publique and l’Agence Française de Développement (AFD, AMES).
Competing interests
The authors declare that they have no conflicts of interest.
Ethical standards
This study was conducted in accordance with the Declaration of Helsinki and approved by the ‘Comité d’Ethique de la Recherche en Sciences de la Santé’ (CERSSA) (N°275/MRSIT/IRSSA/CERSSA-2021).



 Open access
Open access