



Introduction
The genus Udonella Johnston, 1835 (Monopisthocotylea: Udonellidae) represents a distinctive group of marine flatworms that highlights the complexity of host-parasite interactions in aquatic ecosystems. Unlike typical monopisthocotylan parasites, Udonella species have evolved a highly specialized epibiotic lifestyle, attaching to the surface of parasitic copepods and branchiurans, primarily caligids or argulids, for transport and attachment without consuming them (Kabata, Reference Kabata1973; Freeman and Ogawa, Reference Freeman and Ogawa2010; van As, Reference van As, Smit, Bruce and Hadfield2019). While hyperparasitism refers to the consumption of parasites by other parasites (Louvard et al., Reference Louvard, Hadfield, Vanhove, Sures, Smit, Smit and Sures2025), Udonella is functionally an epibiont on the copepod and directly feeds on the fish host (Kabata, Reference Kabata1973; van As, Reference van As, Smit, Bruce and Hadfield2019). This tripartite relationship challenges conventional parasitological paradigms and provides valuable insights into the transition from hyperparasitism to epibiosis in marine environments (Carvajal et al., Reference Carvajal, Ruiz and Sepúlveda2001; van As, Reference van As, Smit, Bruce and Hadfield2019).
Despite their widespread occurrence in marine environments, only 9 species of Udonella have been described globally (WoRMS, 2026) (see Table 1). Taxonomic classification of Udonella has historically been problematic due to deviation of this genus from other monopisthocotylans and the limited availability of clear diagnostic morphological characters (Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021). Specimens lack ciliated larvae and the characteristic haptoral hooks used for attachment in other monopisthocotylans (Littlewood et al., Reference Littlewood, Rohde and Clough1998). Instead, they possess a modified, hookless haptor specialized for adhering to the hard exoskeleton of parasitic crustacean hosts (Littlewood et al., Reference Littlewood, Rohde and Clough1998; Carvajal et al., Reference Carvajal, Ruiz and Sepúlveda2001; Freeman and Ogawa, Reference Freeman and Ogawa2010). As a result, genetically distinct lineages may have historically been grouped under broadly defined species based primarily on general morphology and host association (Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021). The type species Udonella caligorum Johnston, 1835 has consequently been reported from diverse regions, including the Atlantic, Indian, and Pacific coasts of America, Europe, Africa, Australia, and New Zealand (Olivier et al., Reference Olivier, Dippenaar, Khalil and Mokgalong2000; Grobler et al., Reference Grobler, Christison, Oliver and van As2003; Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021). However, molecular analyses suggest that these records likely represent a complex of cryptic species, rather than a single cosmopolitan species (Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021). Phylogenetic studies using the 18S rDNA marker have proven especially powerful for resolving species boundaries, host-specificity patterns, and biogeographical distributions in this morphologically conserved genus (Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021).
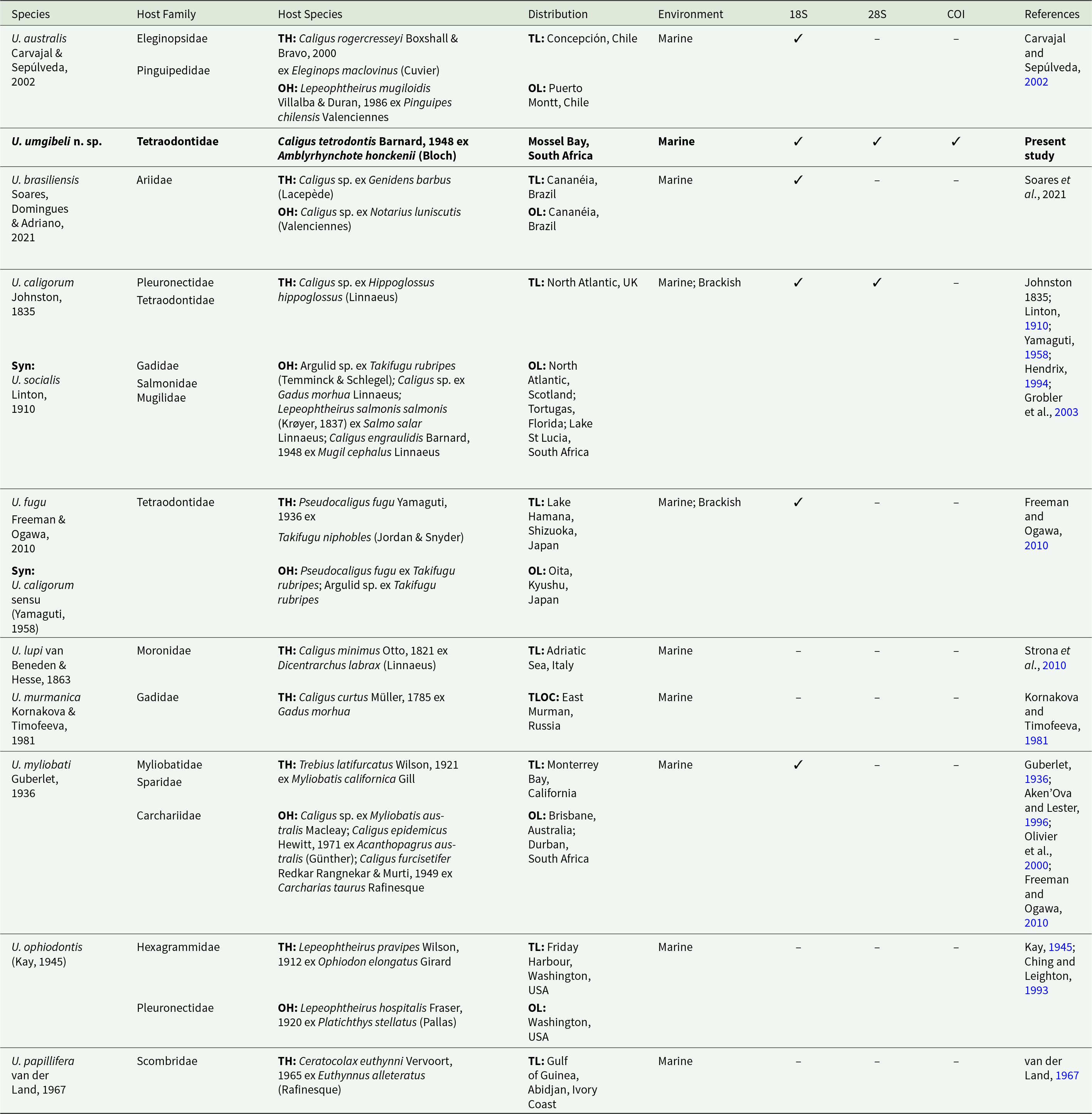
Updated information for all species of Udonella Johnston, 1835 described, with information on host species, host families, distribution and available GenBank data

Table 1 Long description
The table lists described Udonella species and summarizes their host families and host species, geographic distribution and habitat, and whether GenBank sequences are available for 18S and 28S rDNA, and COI mtDNA. Udonella umgibeli n. sp. is recorded from Caligus tetrodontis on Amblyrhynchote honckenii in Mossel Bay, South Africa, and is the only entry with all three markers available. Several species have partial molecular coverage: U. australis, U. brasiliensis, U. fugu, and U. myliobati each have 18S rDNA only, while U. caligorum has 18S and 28S rDNA, but lacks COI mtDNA. Multiple species have no sequence data reported (U. lupi, U. murmanica, U. ophiodontis, U. papillifera). Hosts are mainly parasitic copepods such as Caligus, Lepeophtheirus, Pseudocaligus, Ceratocolax, and Trebius, associated with diverse fish families, and environments are predominantly marine, with some marine and brackish records. Some rows include additional hosts and localities and a synonym note, so host range and distribution may be broader than the primary type records shown.rDNA only
Information from the present study is represented in bold.
Syn – synonym; OH – other hosts; OL – other localities; TH – type host; TL – type locality.
In South Africa, research on marine parasites is limited (Smit and Hadfield, Reference Smit and Hadfield2015; Schaeffner and Smit, Reference Schaeffner and Smit2019). Within the genus Udonella, only sporadic records have been reported. The first South African report was U. myliobati (Guberlet, Reference Guberlet1936) ex Caligus furcisetifer Redkar, Rangnekar & Murti, 1949, parasitizing the sand tiger shark, Carcharias taurus Rafinesque, off the coast of KwaZulu-Natal (Olivier et al., Reference Olivier, Dippenaar, Khalil and Mokgalong2000). Shortly thereafter, Grobler et al. (Reference Grobler, Christison, Oliver and van As2003) documented U. caligorum ex Caligus engraulidis Barnard, 1948, parasitizing the flathead grey mullet, Mugil cephalus Linnaeus, from Lake St Lucia in KwaZulu-Natal. However, none of these records included molecular characterization of the udonellids. Given the persistent taxonomic uncertainty within this genus and increasing evidence that broadly distributed species such as U. caligorum may represent complexes of cryptic species, an integrative approach is essential. The present study, therefore, combines morphological, morphometrical and molecular data (18S and 28S rDNA, and COI mtDNA) to characterize Udonella specimens collected from Caligus tetrodontis Barnard, 1948, parasitizing the evileye blaasop, Amblyrhynchote honckenii (Bloch), off the coast of South Africa. Specifically, the study aimed to provide detailed morphological descriptions of the South African specimens; to establish their phylogenetic placement through molecular analysis and to contribute to a broader understanding of Udonella biogeography and host associations. By documenting the first confirmed molecularly characterized Udonella from the southwestern Indian Ocean, this study expands knowledge of the diversity and distribution of this genus. Furthermore, it highlights the importance of integrative taxonomy in resolving cryptic species complexes.
Materials and methods
Sample collection
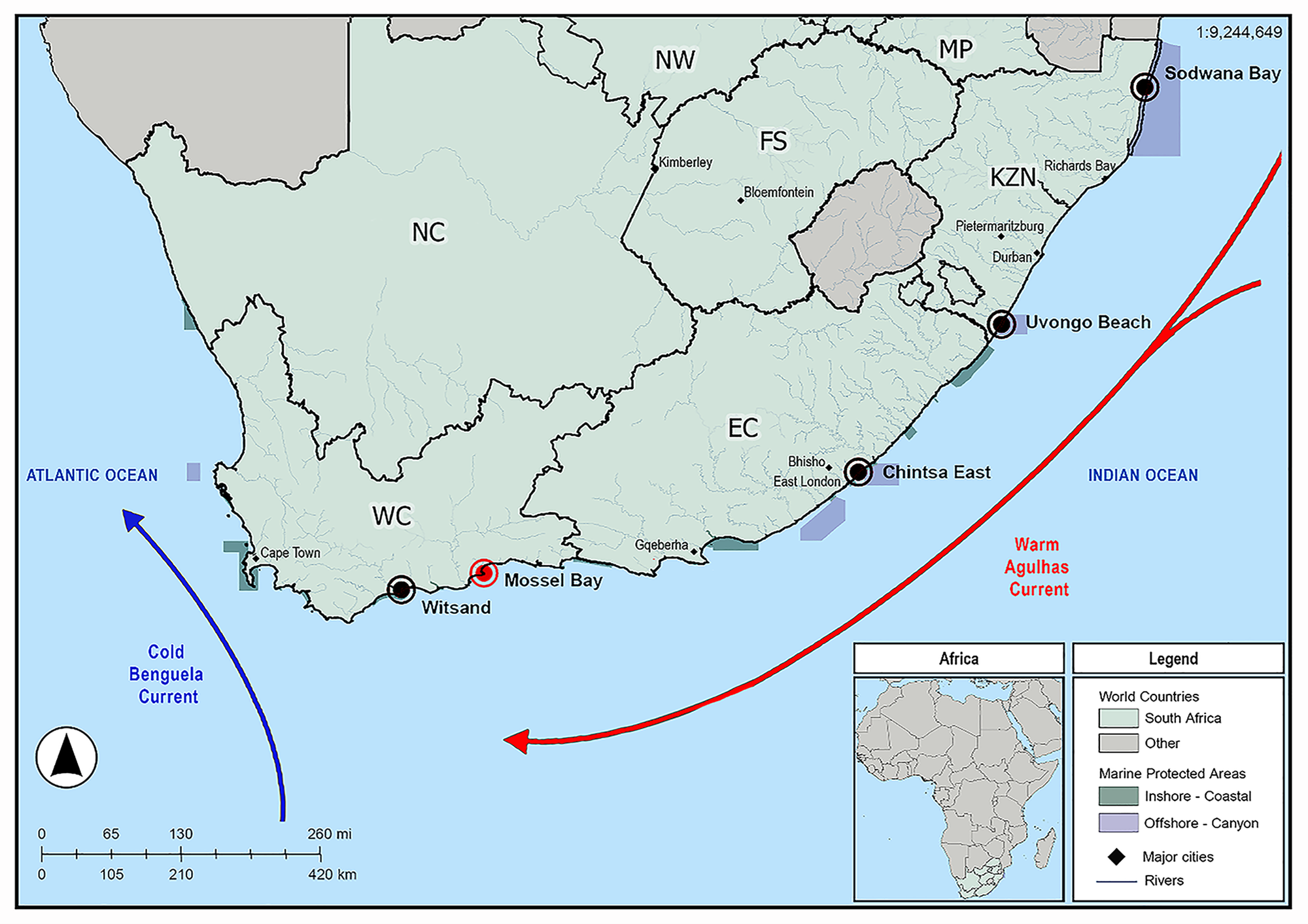
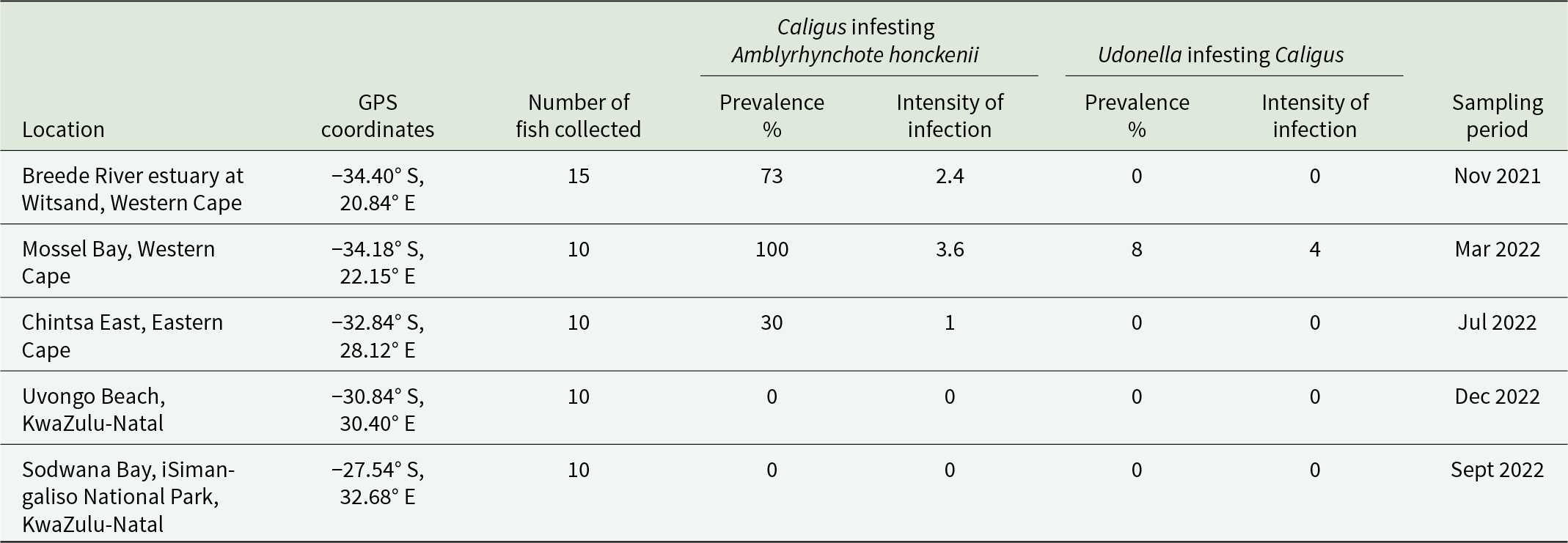
Fifty-five A. honckenii specimens were collected from 5 coastal locations along South Africa (Figure 1). Specimens were collected, identified, processed and screened for parasites as described by Van Der Spuy et al. (Reference Van Der Spuy, Erasmus, Nachev, Schaeffner, Sures, Wepener and Smit2023). Table 2 provides details on each sampling locality, including geographic coordinates, fish collection numbers, parasite infection prevalence, intensity of infection and sampling dates. Caligus Müller O.F., 1785 prevalence is shown as a percentage of infested fish, with mean intensity (parasites per infested host), while Udonella prevalence is shown as a percentage of infested Caligus, with mean intensity (parasites per infested Caligus). Ethical approval for this project was obtained from the Animal Care Research Ethics Committee of North-West University (NWU-00565-19-A5). Permits for the collection of A. honckenii specimens were issued by the South African Department of Agriculture, Forestry and Fisheries (permit nos.: RES2022-44; RES2022-86; and RES2025-16) and Cape Nature (permit no. CN44-87-18289).
Map indicating the sampling localities of specimens of Amblyrhynchote honckenii (Bloch). The red circle indicates the locality where Udonella Johnston, 1835 specimens were collected, while black circles indicate all other sampling localities.

Figure 1 Long description
The map of South Africa highlights sampling localities along the coast with specific markers. A red circle marks Mossel Bay, while black circles indicate Witsand, Chintsa East, Uvongo Beach and Sodwana Bay. The map also shows the Cold Benguela Current flowing northward along the Atlantic Ocean and the Warm Agulhas Current flowing southward along the Indian Ocean. The legend identifiesSouth Africa, marine protected areas and rivers. An inset map of Africa provides regional context. The scale bar indicates distances in kilometers and a compass rose shows orientation.
Parasite infestation data for Amblyrhynchote honckenii (Bloch) collected from 5 South African coastal localities (listed from west to east)

Table 2 Long description
The table reports parasite infestation in Amblyrhynchote honckenii sampled at five South African coastal sites, listing coordinates, fish sample sizes, and sampling months. For Caligus on fish, prevalence was 73 % at Breede River estuary (15 fish, mean intensity 2.4), 100 % at Mossel Bay (10 fish, mean intensity 3.6), and 30 % at Chintsa East (10 fish, mean intensity 1). No Caligus were found at Uvongo Beach or Sodwana Bay (10 fish each). For Udonella on Caligus, prevalence was zero at all sites except Mossel Bay, where 8 % of Caligus were infested with a mean intensity of 4. Sampling occurred from November 2021 to December 2022, with sites ordered west to east.
The table provides details on each sampling locality, including geographic coordinates, fish collection numbers, parasite infection prevalence, intensity of infection and the sampling dates. Caligus Müller O.F., 1785 prevalence is shown as a percentage of infested fish, with mean intensity (parasites per infested host), while Udonella Johnston, 1835 prevalence is shown as a percentage of infested Caligus, with mean intensity (parasites per infested Caligus).
Morphological analyses
Specimens of Caligus and Udonella were removed from the body surface of A. honckenii using fine brushes and fixed in 80% ethanol for morphological analysis and in 96% molecular grade ethanol for molecular characterization. Caligus specimens collected from the fins and external body surfaces were rehydrated in distilled water, cleared in lactophenol, and temporarily mounted onto slides for microscopic observation. Udonella specimens were processed following Freeman and Ogawa (Reference Freeman and Ogawa2010), with minor adaptations. Specimens were stained with aceto-carmine, destained with 1% hydrochloric acid, dehydrated in a graded ethanol series (70%, 80%, 90% and 100%), cleared in methyl salicylate and permanently mounted in Canada balsam. Images and measurements of characteristic body structures were acquired using a Nikon Y-TV55 video camera mounted on a Nikon ECLIPSE Ni microscope (Nikon, Tokyo, Japan). Image analysis software Image Pro Express (Nikon, Japan) was used to obtain all necessary measurements for descriptive analyses. All measurements were taken according to specifications given by Freeman and Ogawa (Reference Freeman and Ogawa2010). Measurements are given both in text descriptions, presented as the mean, and in tabular form, presented as the range, followed by the number (n) of specimens measured in parentheses. Measurements are all presented in micrometres. Drawings were made using a drawing attachment tube. Drawings were scanned and digitized using Adobe Illustrator© software 2025 v. 28.5 (Adobe Inc., San Jose, CA, USA). Definitions of prevalence and mean intensity followed Bush et al. (Reference Bush, Lafferty, Lotz and Shostak1997).
Molecular analyses
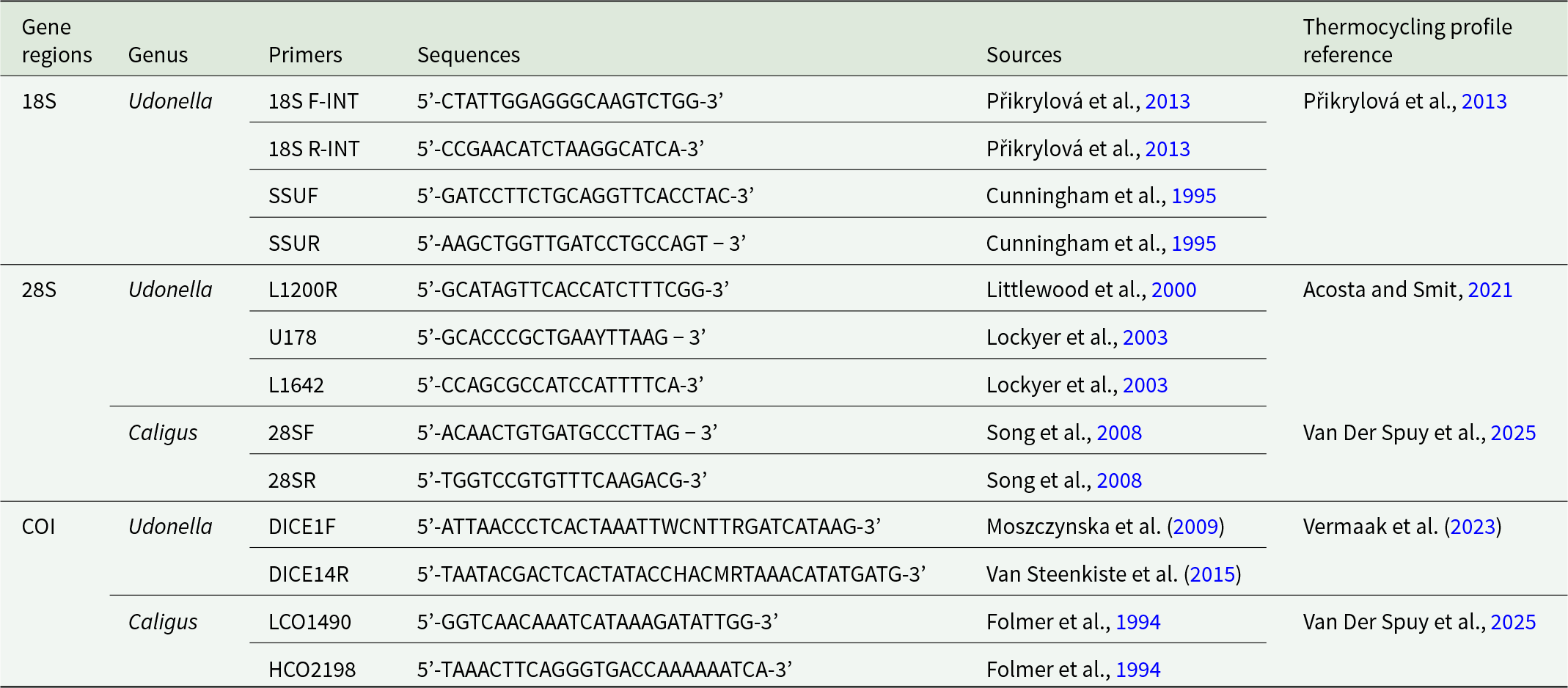
Four adult Caligus specimens and 3 Udonella specimens were selected for molecular analysis. For caligid specimens, genomic DNA was extracted specifically from egg strings, following the method described by Van Der Spuy et al. (Reference Van Der Spuy, Narciso, Hadfield, Wepener and Smit2025). This protocol followed the Macherey-Nagel NucleoSpin® Tissue Kit (Macherey-Nagel GmbH & Co KG, Sandton, South Africa), with 2 modifications: a 4-h pre-lysis incubation step and the addition of 50% more buffer BE, as recommended in Van Der Spuy et al. (Reference Van Der Spuy, Narciso, Hadfield, Wepener and Smit2025). DNA extractions for the udonellids were performed using the PCRBiosystems Rapid DNA Extraction Kit, following the manufacturer’s instructions with slight modifications. Specifically, 10 μL of lysis buffer and 5 μL of proteinase K-containing buffer were used, and the final extracted DNA was diluted to a volume of 300 μL. Subsequent gene amplification targeted the partial 18S and 28S regions of ribosomal DNA (rDNA), and the partial COI mtDNA gene (primers used for amplification are listed in Table 3). Successful PCR amplifications were confirmed by electrophoresis on 1% agarose gels. Verified products were submitted to Inqaba Biotechnical Industries (Pty) Ltd in Pretoria, South Africa, for purification and sequencing. Sequences were assembled, aligned, edited and trimmed using Geneious Prime version 2024.0.7 (Biomatters, Auckland, New Zealand). Additionally, the nucleotide Basic Local Alignment Search Tool (BLAST) was used to support identification of sequence affiliations and facilitate outgroup selection (Table 4). Partial 18S rDNA sequences were used to confirm the species-level identity of the sequenced specimens. Although partial 28S rDNA sequences were also generated during the study, not enough 28S rDNA sequences for udonellids are available on GenBank for a comparative analysis. Additionally, a partial COI mtDNA sequence was also obtained for the new species. However, as no COI mtDNA sequences are currently available on GenBank for any member of the genus, comparative or phylogenetic analyses using this marker could not be performed.
List of primers used for 18S and 28S rDNA, and COI mtDNA amplification of udonellids with sequences and references, used to amplify partial 18S and 28S rDNA and COI mtDNA gene regions in this study

Table 3 Long description
The table lists PCR primers used to amplify three gene regions, 18S and 28S rDNA, and COI mtDNA, for the genera Udonella and Caligus. For 18S rDNA in Udonella, four primers are provided: 18S F-INT and 18S R-INT, plus SSUF and SSUR, with sequences sourced from Přikrylová et al. 2013 and Cunningham et al. 1995, and the thermocycling profile referenced to Přikrylová et al. 2013. For 28S rDNA in Udonella, three primers are listed (L1200R, U178, L1642) with sequences from Littlewood et al. 2000 and Lockyer et al. 2003, and thermocycling referenced to Acosta and Smit 2021. For 28S rDNA in Caligus, two primers are given (28SF and 28SR) from Song et al. 2008, with thermocycling referenced to Van Der Spuy et al. 2025. For COI mtDNA, Udonella uses DICE1F from Moszczynska et al. 2009 and DICE14R from Van Steenkiste et al. 2015, with thermocycling referenced to Vermaak et al. 2023, while Caligus uses the Folmer primers LCO1490 and HCO2198 from Folmer et al. 1994 with thermocycling referenced to Van Der Spuy et al. 2025. Sequences are presented in the table as primer nucleotide strings.
List of GenBank udonellid sequences included in the phylogenetic analyses

Table 4 Long description
The table lists udonellid parasite taxa and one outgroup, giving each taxon’s host association, collection locality, and GenBank accession numbers for three markers: 18S rDNA, 28S rDNA, and COI mtDNA, plus the literature source. Udonella umgibeli n. sp. from South Africa on Caligus tetrodontis from Amblyrhynchote honckenii provides multiple accessions for all three markers, including two entries each for 18S and 28S rDNA and one COI mtDNA accession. Most other Udonella taxa include only an 18S rDNA accession and have no 28S rDNA or COI mtDNA listed, including Udonella australis from Chile, Udonella fugu from Japan, and Udonella myliobati from Australia. Udonella brasiliensis from Brazil includes two 18S rDNA accessions tied to different Caligus hosts and lacks 28S rDNA and COI mtDNA. Udonella caligorum from the UK includes both 18S and 28S rDNA accessions and is associated with two copepod hosts, one from Gadus morhua and one from Salmo salar. The outgroup, Gyrodactylus salmonis from Mexico on Oncorhynchus mykiss, includes an 18S rDNA accession only. Dashes indicate missing sequence data for a marker rather than confirmed absence in nature.
The taxa in bold font are sequences generated from the present study. Gyrodactylus salmonis Yin & Sproston, 1948 (in the grey shade) was used as the outgroup.
Results
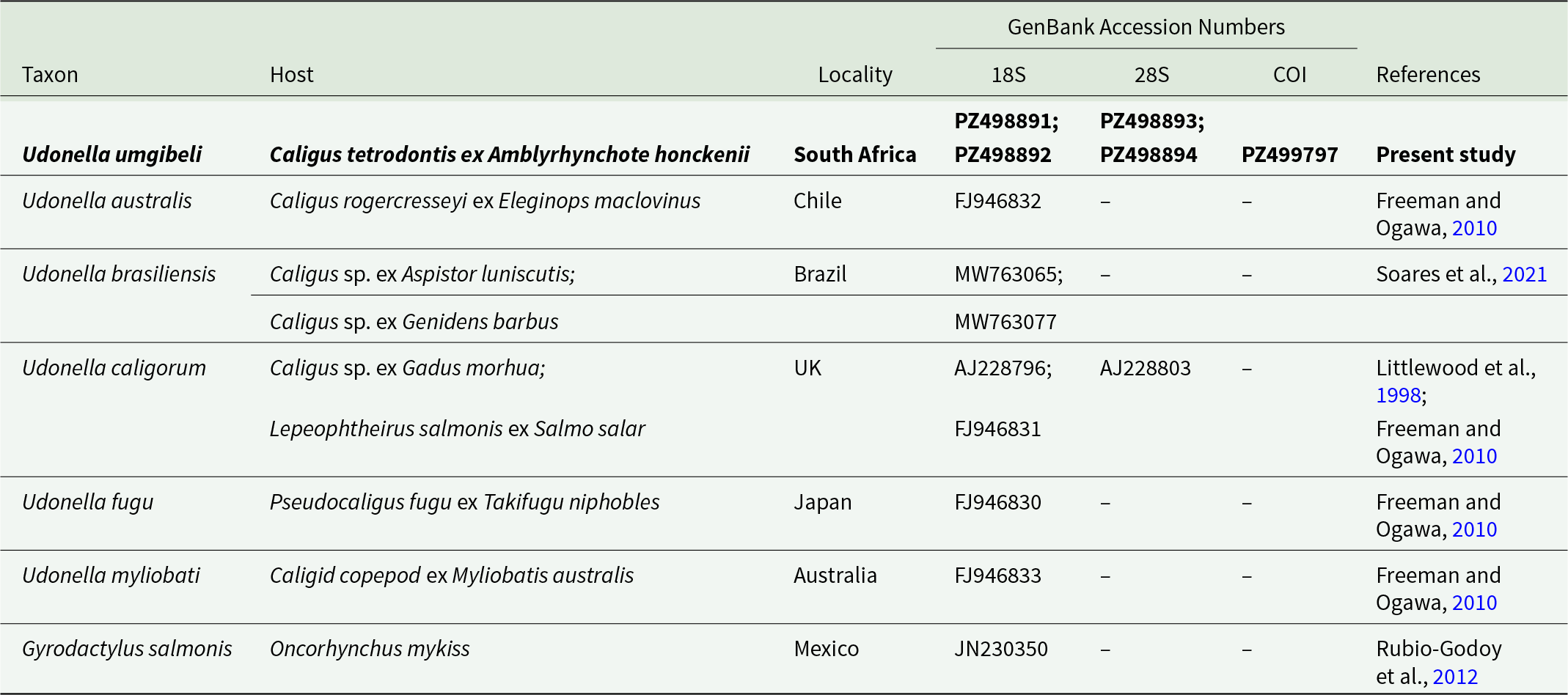
A total of 66 C. tetrodontis specimens were collected from 3 localities (Table 2). Additionally, 12 udonellids were collected from 3 Caligus specimens infesting the body surface of a single A. honckenii specimen (Figure 2).
Light microscopy images of Udonella umgibeli n. sp. ex Caligus tetrodontis Barnard, 1948, parasitizing Amblyrhynchote honckenii (Bloch) off the coast of South Africa: A – Udonella umgibeli n. sp. ex C. tetrodontis, scale bar 2 mm; B – Specimens on A. honckenii; C – Specimens on A. honckenii; D – Eggs, scale bar 500 µm.

Caligus specimens were identified based on a combination of morphological traits and molecular evidence, matching the published morphometrics of C. tetrodontis reported by Hayes et al. (Reference Hayes, Christison, Vaughan, Smit and Boxshall2021), including a subcircular cephalothoracic shield, a rounded genital complex, a single-segmented abdomen, and distinctive fourth-leg spines. Additionally, molecular analysis of the mitochondrial COI mtDNA gene confirmed the morphological identification, showing 100% sequence identity to published C. tetrodontis sequences (Hayes et al., Reference Hayes, Christison, Vaughan, Smit and Boxshall2021).
Although 2 species of Udonella have previously been reported from South Africa, this is the first study to include genetic data (18S and 28S rDNA, and COI mtDNA) for a South African representative of the genus. An integrative approach combining detailed morphometric comparisons and molecular phylogenetic analyses with known congeners demonstrates that the South African material represents a species new to science, herein described as Udonella umgibeli n. sp.
Taxonomic summary
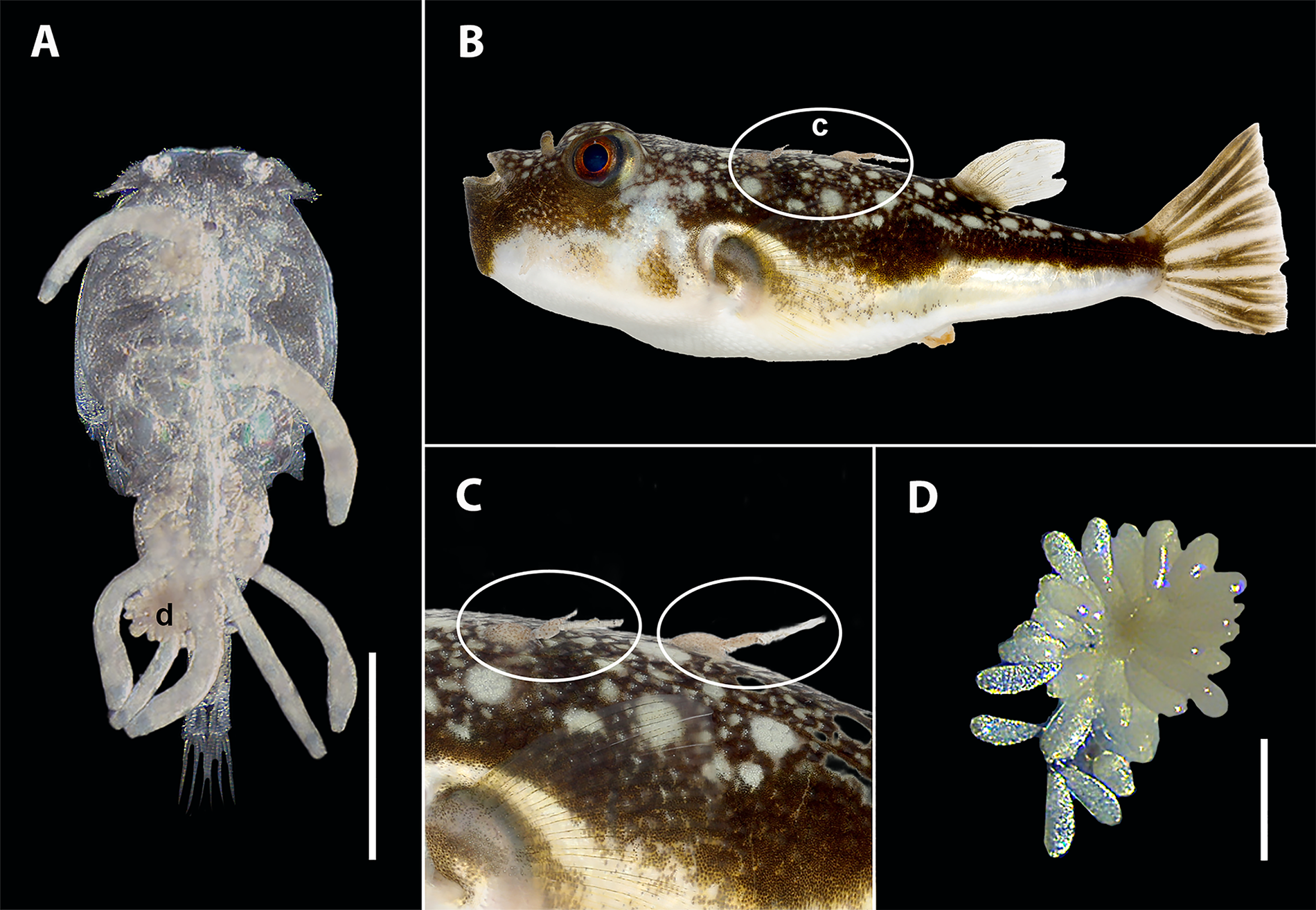
Udonella umgibeli n. sp.: Figure 3
Illustrations of an adult Udonella umgibeli n. sp. Specimen: A – Entire body of holotype (NMB P 1330), ventral view, scale bar 200 µm; B – Egg, scale bar 100 µm.

ZooBank LSID: urn:lsid:zoobank.org:act:26DAED93-0C72-4C67-904D-83F7398B6BF0.
Type host: Caligus tetrodontis Barnard, 1948 ex. A. honckenii (Bloch).
Type locality: Mossel Bay, South Africa (−34.179656, 22.152093) (Figure 1).
Site of infection: Dorsal surface of carapace of C. tetrodontis ex A. honckenii (Figure 2).
Prevalence (P): 7% on fish hosts, 8% on copepod hosts.
Intensity (MI): 12 on fish hosts, 4 on copepod hosts.
Type material: 8 morphological specimens fixed on slides (Holotype: NMB P 1330, Paratypes: NMB P 1331 – NMB P 1338) and 3 genetic vouchers fixed in ethanol (2 Udonella specimens: NMB P 1339 – NMB P 1340; and 1 Caligus specimen: NMB P 1341) ex C. tetrodontis collected from Mossel Bay, South Africa.
Representative DNA sequences: For U. umgibeli n. sp.: 3 identical 18S rDNA replicates of 969–1800 bp, 2 submitted to GenBank (PZ498891–PZ498892); 3 identical 28S rDNA replicates of 979–1481 bp, 2 submitted to GenBank (PZ498893–PZ498894); and 1 COI mtDNA sequence of 710 bp, submitted to GenBank (PZ499797), were obtained. For Caligus tetrodontis: 3 identical 28S rDNA replicates of 719–720 bp, 1 submitted to GenBank (PZ498221); and 4 identical COI replicates of 653–680 bp, 1 submitted to GenBank (PZ498225), were obtained.
Etymology: The species name ‘umgibeli’ is derived from the isiZulu word meaning ‘rider’ or ‘passenger’. The term refers to one who is carried by another, therefore reflecting the organism’s epibiotic lifestyle.
Description
Based on measurements of 6 whole-mounts of mature worms. Body flat, subcylindrical 1170 µm in total length, 326 µm in width, widest at ovarian level. Haptor unarmed, muscular, disc-shaped, 344 µm in diameter (Figure 3A). Four groups of haptoral glands, immediately posterior and slightly overlapping vitellaria (Figure 3A). Collecting ducts of adhesive glands join together in common duct on median line, leading to centre of haptor, where it opens ventrally. Pair of anterior head organs fed by single group of gland cells, extending on each side of body from level of postpharyngeal gland to anterior part of intestine (Figure 3A). Three papilla-like projections at distal end, located obliquely near anterior end of body. One pair of conical sensory papillae prominent, 10 µm in height (Figure 3A). Mouth subterminal, median. Pharynx ovate, and transversely wide, 144 µm long by 106 µm wide. No oesophagus. Intestine a wide tubular sac, running posteriorly in dorsal median field, extending to end of vitellaria (Figure 3A).
Testis single, subspherical, 191 µm long by 146 µm wide, posterior, overlapping with ovary (Figure 3A). Vas deferens narrow, running on left side of ovary and uterus. Vesicula seminalis elliptical, 27 µm long by 10 µm wide, situated alongside prostatic reservoir behind egg. Ejaculatory bulb 34 µm long by 20 µm wide. Prostatic reservoir spherical, 22 µm long by 32 µm wide. Genital atrium, 17 µm long by 10 µm wide, opening 13 µm in diameter, posterior to pharynx. Cirrus not differentiated.
Ovary single, spherical 114 µm long by 146 µm wide, located in middle of body, overlapping testis, testis: ovary width ratio 1: 1.04 µm (Figure 3A). Germiduct arising from ventral side of ovary, dilated to form fertilization chamber, with single large oocyte. Fertilization chamber situated in anterior part of ovary on its left side, rounded, 65 µm long by 49 µm wide. Uterus distended with pyriform egg (Figure 3B), 191 µm long by 57 µm wide. Egg provided with long polar filament 391 µm long. Uterus opening directly into the genital atrium. Vitellarium comprising numerous large irregular follicles, extending in each lateral field from level of genital pore to posterior end of intestine.
Remarks
Species-level delimitation in Udonella is challenging due to a scarcity of high-discrimination morphological characters (Freeman and Ogawa, Reference Freeman and Ogawa2010). As a result, only a few features reliably separate species, including body dimensions (length and width), haptor diameter, gonad proportionality (ovary-to-testis size), and genital pore position (Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021). Table 5 summarizes comparative morphometric data for all recognized species. Udonella umgibeli n. sp. can be differentiated from every congener by at least 2 diagnostic characters per comparison.
Comparative metrical information for all species of Udonella Johnston, 1835. Information is presented as the mean, followed by the standard deviation and the number of specimens examined

Table 5 Long description
The table compares body size and organ measurements across nine Udonella species, reporting ranges and, where available, mean values with specimen counts, all in micrometres. Body length varies widely, from small species such as Udonella myliobati (about 430 to 830) and Udonella brasiliensis (about 553 to 785) to very large Udonella murmanica (about 4500 to 7100). Several species are intermediate in length, including Udonella fugu (about 1900 to 3100) and Udonella ophiodontis (about 2000 to 2800), while Udonella umgibeli n. sp. is shorter (about 950 to 1469). Body width generally increases with body length, with Udonella murmanica also having the greatest width range (about 690 to 1040). Haptor diameter is reported for most species and is smallest in Udonella brasiliensis (about 46 to 81) and largest among the reported values in Udonella murmanica (about 410 to 640), while some entries are missing. Pharynx, ovary, testis, egg size, and egg filament length show species-specific ranges, with notably long egg filaments in Udonella murmanica (about 1000 to 1400) and in Udonella caligorum (about 600 to 800). Qualitative traits also differ, including caudal gland grouping patterns and genital pore location, which is commonly mid-ventral and posterior to the pharynx but varies in some species.
All measurements are in micrometres.
L- length; W- width.
The closest morphological congener is Udonella australis Carvajal and Sepúlveda, 2002 (Table 5). However, U. umgibeli n. sp. stands out with several clear morphometric differences, especially in haptor diameter and gonad proportions. Its haptor measures over 317 µm, much larger than the measurements known for U. australis (<310 µm). The testis-to-ovary width ratio is also distinct at 1:0.8–1.4 (with the ovary roughly equal to the testis), compared to 1:0.6–0.75 in U. australis. Additionally, the egg, ovary, and testes of U. umgibeli n. sp. overlap, unlike those of U. australis. While overall body lengths overlap somewhat, U. umgibeli n. sp. tends to be shorter and broader at 1170 µm × 326 µm, vs 1500 µm × 259 µm for U. australis. Finally, U. umgibeli n. sp. also has 4 clumped groups of caudal glands, whereas U. australis shows only one.
Udonella umgibeli n. sp. differs markedly from U. brasiliensis Soares, Domingues & Adriano, U. caligorum and U. myliobati (Guberlet, Reference Guberlet1936), as it is larger in most measured characters (Table 5). Compared to U. brasiliensis, it exceeds the former across all key metrics. For instance, its body is longer (>950 µm vs < 785 µm) and wider (>307 µm vs < 205 µm), with a bigger haptor (>317 µm vs < 81 µm), larger pharynx, gonads, and longer egg filament length (>386 µm vs < 154 µm). These differences are substantial and non-overlapping, providing clear morphometric distinctions. Similarly, U. umgibeli n. sp. is larger than U. caligorum in body width, haptor diameter, testis width, and egg dimensions. However, U. caligorum has a longer egg filament (>600 µm vs < 395 µm) and a more pronounced gonad ratio (1:1.7 vs < 1:1.4). They also differ in genital pore position: U. umgibeli n. sp. has a mid-ventral pore posterior to the pharynx, while U. caligorum shows a sinistral/submarginal pore positioned slightly anterior to the posterior margin of the pharynx. Caudal glands add another distinction, with 4 clumped groups in U. umgibeli n. sp. versus 2 separate groups in U. caligorum. Compared to U. myliobati, U. umgibeli n. sp. is again larger in body length, haptor diameter, pharynx length, ovary length, and testis length. Its caudal glands form 4 clumped groups, unlike the single unicellular group in U. myliobati.
The remaining congeners (Udonella fugu Freeman and Ogawa, Reference Freeman and Ogawa2010; Udonella murmanica Kornakova and Timofeeva, Reference Kornakova and Timofeeva1981; Udonella ophiodontis Ching and Leighton, Reference Ching and Leighton1993 and Udonella papillifera; van der Land, Reference van der Land1967) are generally larger than U. umgibeli n. sp. in most traits (Table 5). For example, U. fugu surpasses U. umgibeli n. sp. in body size, pharynx, gonads and egg width, but U. umgibeli n. sp. has a longer egg filament (>386 µm vs < 310 µm) and 4 caudal gland groups instead of 2. Udonella murmanica is substantially larger than U. umgibeli n. sp. across nearly all characters, with body lengths over 4500 µm and egg filaments reaching 1000 µm. Udonella ophiodontis and U. papillifera follow suit, exceeding it in body, pharynx, gonadal, and egg dimensions. Genital pore position sets them further apart. Udonella ophiodontis has a submarginal pore and U. papillifera a ventral pore offset left of the midline, while U. umgibeli n. sp. possesses a mid-ventral, median pore posterior to the pharynx. Differences in caudal gland arrangement also reinforce these distinctions. These consistent morphometric, anatomical, and organizational differences clearly distinguish U. umgibeli n. sp. from all known congeners.
Molecular characterization
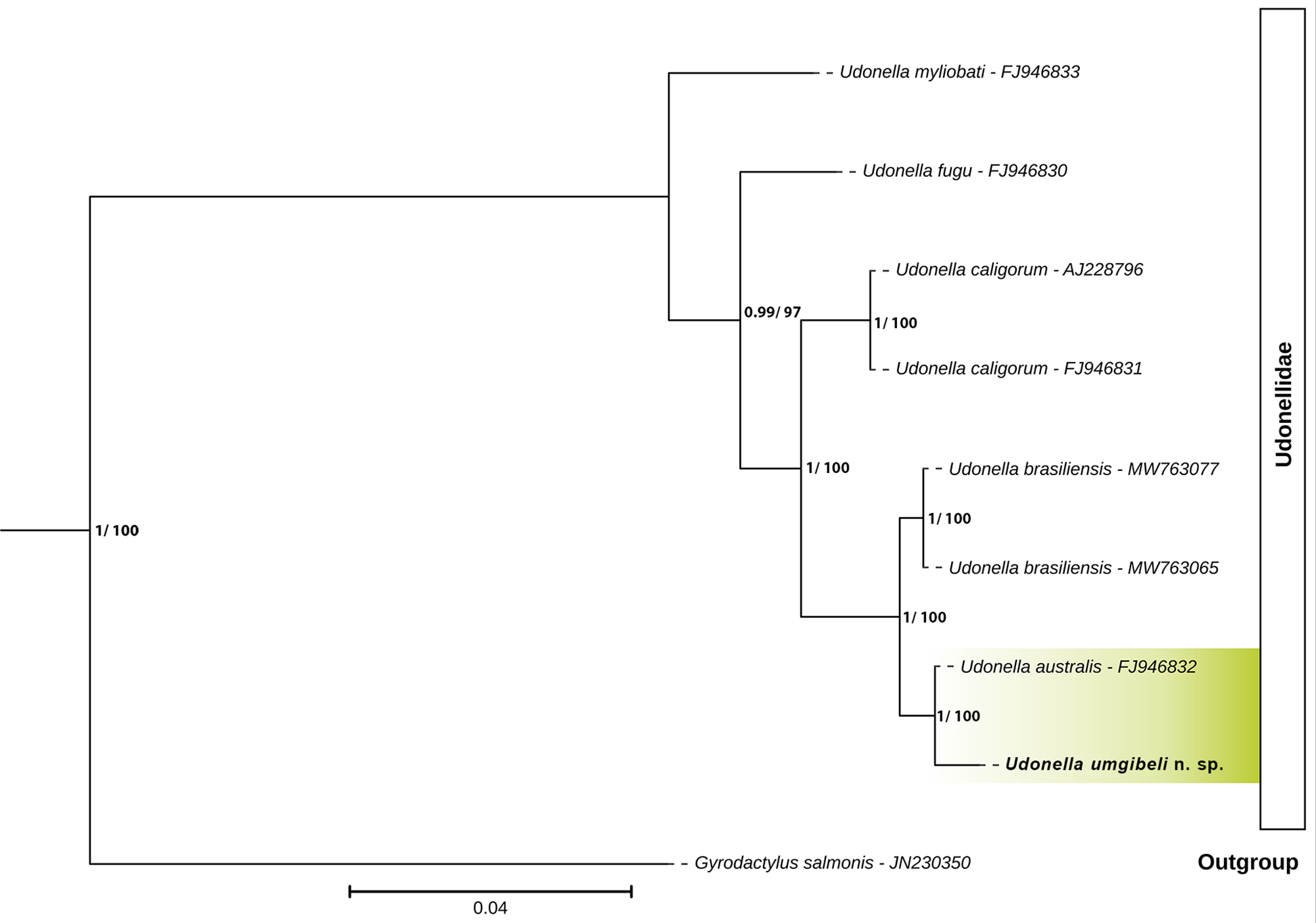
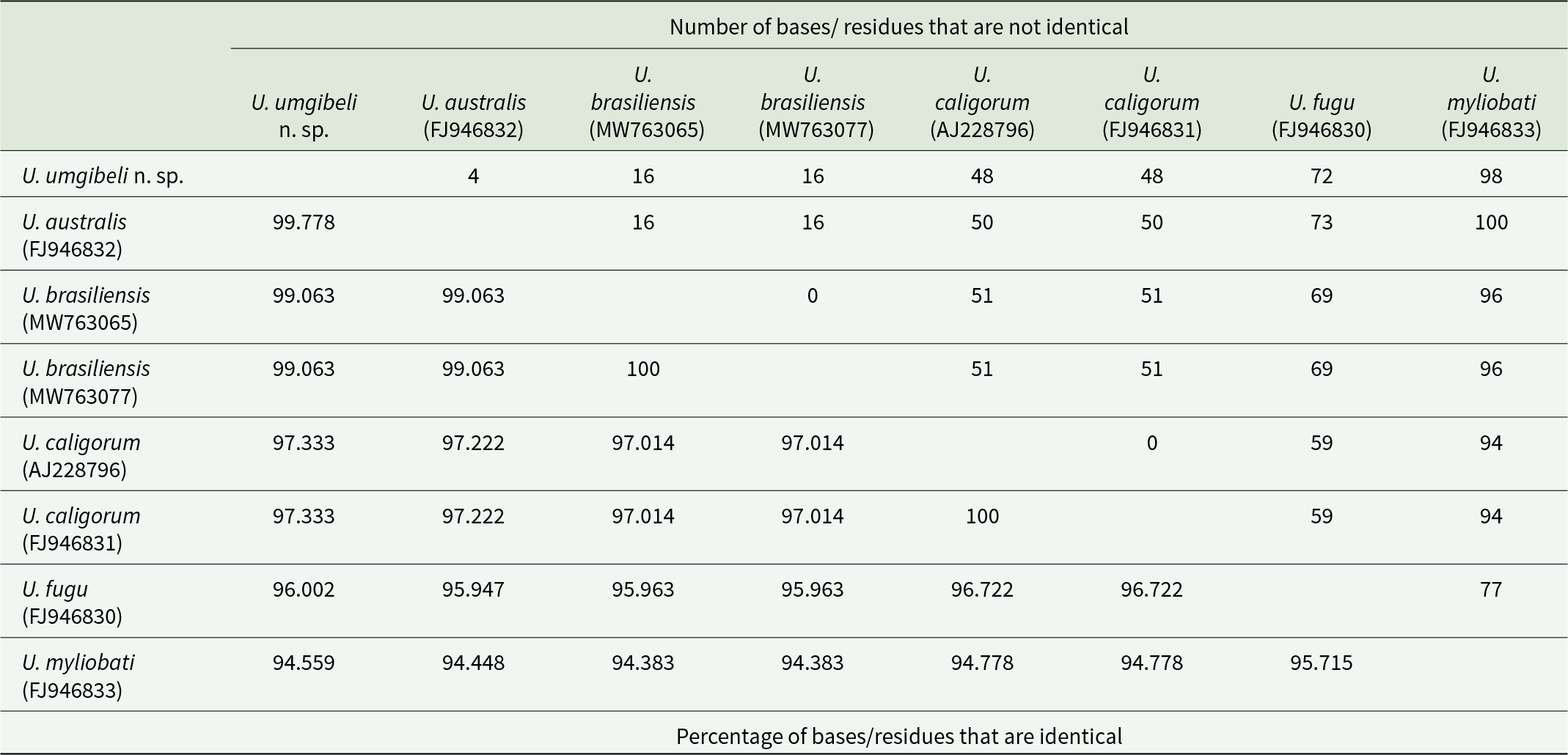
The molecular data provide convergent evidence supporting the recognition of U. umgibeli n. sp. as a new species. Three specimens of U. umgibeli n. sp. were successfully sequenced for partial 18S (969–1800 bp) and 28S (979–1481 bp) rDNA fragments. All sequences obtained were 100% identical across specimens for both markers. Phylogenetic and pairwise genetic analyses based on 18S rDNA revealed that U. umgibeli n. sp. is most closely related to U. australis, differing by only 4 bp (99.77% identity across an 1811 bp alignment) (see Table 6; Figure 4). Sequence identities to other congeners were increasingly lower, with 99.06% to U. brasiliensis (16 bp differences), 97.33% to U. caligorum (48 bp differences), 96.00% to U. fugu (72 bp differences) and 94.56% to U. myliobati (98 bp differences). Phylogenetic trees, therefore, place U. umgibeli n. sp. in a well-supported clade with U. australis, separate from other sequenced udonellids (Figure 4). The slight but consistent 18S rDNA divergence from U. australis is reinforced by multiple non-overlapping morphological differences, indicating that the 2 taxa are not conspecific.
Bayesian inference (BI) phylogenetic tree of the family Udonellidae, based on 18S rDNA sequences. Nodal support values are indicated as BI/maximum likelihood (ML); only values ≥ 0.7 (BI) and ≥ 70 (ML) are displayed. Newly sequenced isolates generated in this study are highlighted in green with bold text. Gyrodactylus salmonis Yin & Sproston, 1948 was selected as the outgroup.

Figure 4 Long description
The diagram is a Bayesian Inference phylogenetic tree of the family Udonellidae, oriented horizontally from left to right. The tree is based on 18S rDNA sequences. Nodal support values are indicated as Bayesian Inference over Maximum Likelihood, with only values greater than or equal to 0.7 for Bayesian Inference and 70 for Maximum Likelihood displayed. The tree begins with the outgroup Gyrodactylus salmonis, labeled with its sequence identifier JN230350. The main clade, labeled Udonellidae, branches off to include several species. Udonella myliobati, with sequence FJ946833, is the first branch. Following this, Udonella fugu, labeled FJ946830, branches off. Next, Udonella caligorum appears twice, with sequences AJ228796 and FJ946831. Udonella brasiliensis is shown with two sequences, MW763077 and MW763065. Udonella australis, labeled FJ946832, is closely related to Udonella umgibeli n. sp., which is highlighted in green. The scale bar at the bottom represents a genetic distance of 0.04.
Nucleotide comparison of partial 18S rDNA sequences of Udonella Johnston, 1835, based on a 1811 bp alignment

Table 6 Long description
The table compares pairwise similarity among partial 18S rDNA sequences from eight Udonella samples, reporting mismatched base counts and the corresponding percent identity. The closest matches are within species: the two U. brasiliensis sequenceshave zero differences and 100 % identity, and the two U. caligorum sequences also have zero differences and 100 % identity. Udonella umgibeli n. sp. is very similar to U. australis with 4 mismatches and 99.778 % identity, and it is also close to U. brasiliensis with 16 mismatches and 99.063 % identity. Differences increase between U. umgibeli and U. caligorum with 48 mismatches and 97.333 % identity, and further with U. fugu at 72 mismatches and 96.002 %identity. The most divergent pairing in the table is U. umgibeli n.sp. versus U. myliobati with 98 mismatches and 94.559 % identity. Overall, identities range from about 94.383 to 100 %, indicating high conservation across these sequences, with the largest separations involving U. myliobati.
Analysis of the 28S rDNA region further supports species-level distinction. The only available comparative sequence in GenBank (U. caligorum, AY228803) differs from U. umgibeli n. sp. by 65 bp (91.7% identity). This divergence exceeds typical intraspecific variation and clearly excludes U. umgibeli n. sp. from the historically broad U. caligorum complex. In addition to the rDNA markers, a partial COI mtDNA sequence was generated for U. umgibeli n. sp. However, as no COI mtDNA sequences are currently available for any member of the genus Udonella, comparative analyses using this marker were not possible. Nevertheless, this sequence represents the first COI mtDNA record for the genus and provides an important reference for future studies as additional udonellid COI mtDNA data become available.
Discussion
The description of U. umgibeli n. sp. from the southwestern Indian Ocean illustrates how integrative taxonomy can resolve species boundaries in a morphologically conservative genus. The present study shows that the South African material differs from all currently recognized congeners by a combination of diagnostic morphological traits and genetic divergence in the 18S and 28S rDNA regions. Although the 18S rDNA sequence differs from that of U. australis by only 4 bp (99.77% identity over 1811 bp), the combined morphological and molecular evidence supports the recognition of U. umgibeli n. sp. as a distinct species.
Nuclear 18S rDNA is a highly conserved, multi-copy nuclear locus that often lacks species-level resolution in Platyhelminthes because ribosomal DNA arrays are homogenized through concerted evolution (Littlewood et al., Reference Littlewood, Rohde and Clough1998; Poisot et al., Reference Poisot, Verneau and Desdevises2011; Vanhove et al., Reference Vanhove, Tessens, Schoelinck, Jondelius, Littlewood, Artois and Huyse2013; Ondračková et al., Reference Ondračková, Seifertová, Bryjová, Leis and Jurajda2020; Bargues et al., Reference Bargues, Halajian, Artigas, Luus-Powell, Valero and Mas-coma2022; Krupenko et al., Reference Krupenko, Kremnev, Gonchar, Uryadova, Miroliubov, Krapivin, Skobkina, Gubler and Knyazeva2022). Consequently, morphologically and ecologically distinct taxa may differ by only a few bases, or even share identical 18S rDNA sequences, while remaining recognizable as separate species. This limitation is well documented in monopisthocotylans. For example, Gyrodactylus melas Ondračková, Seifertová & Leis in Ondračková et al., Reference Ondračková, Seifertová, Bryjová, Leis and Jurajda2020 differs from G. nebulosus Kritsky & Mizelle, 1968 by a single base (0.2%) over a 432-bp 18S rDNA fragment, yet both are recognized as distinct species based on morphological and ecological evidence (Ondračková et al., Reference Ondračková, Seifertová, Bryjová, Leis and Jurajda2020). Likewise, Lamellodiscus fraternus Bychowsky, 1957 and L. ergensi Euzet & Oliver, 1966 differ by only 0.6% in their 18S rDNA (Poisot et al., Reference Poisot, Verneau and Desdevises2011), while 18S rDNA failed to distinguish Macrogyrodactylus karibae Douëllou & Chishawa, 1995 and M. congolensis (Prudhoe, 1957) Yamaguti, 1963, despite clear morphological differences, leading the authors to conclude that ITS rDNA and COI mtDNA are more reliable markers for closely related taxa (Maduenyane et al., Reference Maduenyane, Dos Santos and Avenant-oldewage2022). Beyond monopisthocotylans, biodiversity and phylogenetic studies have repeatedly shown that 18S rDNA underestimates species-level diversity relative to COI mtDNA, whereas 28S rDNA and mitochondrial markers generally provide greater resolution (Vanhove et al., Reference Vanhove, Tessens, Schoelinck, Jondelius, Littlewood, Artois and Huyse2013; Krupenko et al., Reference Krupenko, Kremnev, Gonchar, Uryadova, Miroliubov, Krapivin, Skobkina, Gubler and Knyazeva2022).
Among Udonella species, pairwise 18S rDNA distances range from 1% to 5.6% (Table 6), with the smallest reported distance of 1% (17 bp) between U. brasiliensis and U. australis (Soares et al., Reference Soares, Domingues and Adriano2021). In this context, the limited 18S rDNA divergence and weak resolution between U. umgibeli n. sp. and U. australis (4 bp difference/0.22% identity) are consistent with published patterns within the genus and should not be regarded as evidence against the recognition of a distinct species. Instead, these results reinforce the importance of evaluating molecular data alongside morphological and ecological evidence when delimiting species in monopisthocotylans (Littlewood et al., Reference Littlewood, Rohde and Clough1998; Freeman and Ogawa, Reference Freeman and Ogawa2010; Vanhove et al., Reference Vanhove, Tessens, Schoelinck, Jondelius, Littlewood, Artois and Huyse2013; Krupenko et al., Reference Krupenko, Kremnev, Gonchar, Uryadova, Miroliubov, Krapivin, Skobkina, Gubler and Knyazeva2022).
The available 28S rDNA comparative data reinforce species-level separation. The partial 28S rDNA sequence of U. umgibeli n. sp. differs from the only U. caligorum sequence in GenBank by 65 bp, indicating greater divergence than observed in the corresponding 18S rDNA comparison. This level of divergence exceeds typical intraspecific variation reported for other monopisthocotylan and polyopisthocotylan groups, and aligns more closely with interspecific differences, particularly given that 28S rDNA is usually more variable than 18S rDNA, while remaining alignable across related taxa (Vanhove et al., Reference Vanhove, Tessens, Schoelinck, Jondelius, Littlewood, Artois and Huyse2013; Chaabane et al., Reference Chaabane, Verneau and Du Preez2019). The 28S rDNA data for U. umgibeli n. sp. thus support its exclusion from the historically broad U. caligorum complex and further support the recognition of the South African lineage as a distinct species.
In monopisthocotylan, species hypotheses remain grounded in diagnosable morphological differences, particularly in structures associated with attachment and reproduction, because these traits are functionally constrained and less prone to transient environmental or fixation artefacts (Vanhove et al., Reference Vanhove, Tessens, Schoelinck, Jondelius, Littlewood, Artois and Huyse2013; Soares et al., Reference Soares, Domingues and Adriano2021). The epibiotic lifestyle of Udonella is likely to impose strong selection on these structures (Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021) and several of the characters described in the present study, including haptoral morphology, gland arrangement and reproductive organ proportions (Table 4), may reflect adaptation to life on caligid copepods (Kabata, Reference Kabata1973; Freeman and Ogawa, Reference Freeman and Ogawa2010; van As, Reference van As, Smit, Bruce and Hadfield2019; Soares et al., Reference Soares, Domingues and Adriano2021). Regardless of their functional significance, these characters consistently distinguish U. umgibeli n. sp. from its congeners and provide multiple independent diagnostic traits supporting its recognition as a distinct species.
Furthermore, host specificity and ecology provide an independent, biologically meaningful line of evidence for species delimitation, particularly in parasites. South African records of Udonella include U. myliobati from a caligid on the ragged-tooth shark and U. caligorum from caligids in estuarine/ marine contexts, but the genus remains poorly sampled relative to the diversity of caligids and marine fishes in the region. In the present study, U. umgibeli n. sp. was collected from C. tetrodontis parasitizing A. honckenii off Mossel Bay, South Africa, representing the first molecularly characterized Udonella from the southwestern Indian Ocean. The species shows a prevalence of 7% in fish hosts and 8% in copepod hosts, with a mean intensity of 4 udonellids per infected copepod, consistent with the patchy occurrence reported for other udonellids (Olivier et al., Reference Olivier, Dippenaar, Khalil and Mokgalong2000; Grobler et al., Reference Grobler, Christison, Oliver and van As2003; Freeman and Ogawa, Reference Freeman and Ogawa2010). Although A. honckenii and its caligid copepod were collected from several localities, U. umgibeli n. sp. was detected only at Mossel Bay, a pattern that may reflect either limited sampling or a more restricted distribution and will require broader geographic sampling to resolve. Alternatively, it may indicate a more localized association, possibly linked to cooler Atlantic-influenced waters rather than the warmer western Indian Ocean sites (see Figure 1). Furthermore, monopisthocotylans are generally regarded as highly host-specific parasites (Whittington et al., Reference Whittington, Cribb, Hamwood and Halliday2000), and, although udonellids attach to copepods rather than directly to fishes, they depend on the fish host as a trophic resource. Because A. honckenii is not the type host of C. tetrodontis, the association reported here may represent only part of the copepod’s host range, which could also influence the observed narrow distribution range of U. umgibeli n. sp.
The association with C. tetrodontis on A. honckenii contrasts with the host records of other South African species, as well as with U. australis, which is known from South American fish-copepod systems, and with global records of U. caligorum that span multiple host-copepod combinations across the Atlantic, Indian, and Pacific Oceans (Freeman and Ogawa, Reference Freeman and Ogawa2010; Soares et al., Reference Soares, Domingues and Adriano2021; WoRMS, 2026). Biogeographic separation between the southwestern Indian Ocean and the South or Southeast Atlantic regions, together with differences in host-copepod associations, further supports the distinction of the Mossel Bay material from U. australis and the broadly defined U. caligorum complex (Lavikainen et al., Reference Lavikainen, Iwaki, Haukisalmi, Konyaev, Casiraghi, Dokuchaev, Galimberti, Halajian, Henttonen, Ichikawa-seki, Itagaki, Krivopalov, Meri, Morand, Näreaho, Olsson, Ribas, Terefe and Nakao2016; Krupenko et al., Reference Krupenko, Kremnev, Gonchar, Uryadova, Miroliubov, Krapivin, Skobkina, Gubler and Knyazeva2022).
Further refinement of species boundaries within the Udonella will require additional molecular and ecological data. Sequencing faster-evolving markers from multiple individuals, particularly 28S rDNA and COI mtDNA, would provide an improved resolution for species-level and population-level analyses (Vanhove et al., Reference Vanhove, Tessens, Schoelinck, Jondelius, Littlewood, Artois and Huyse2013; Chaabane et al., Reference Chaabane, Verneau and Du Preez2019; Ondračková et al., Reference Ondračková, Seifertová, Bryjová, Leis and Jurajda2020). Expanding geographic and host sampling across the known distribution of A. honckenii-related tetraodontiform fishes and their associated caligid copepod species would further clarify host specificity, biogeographic patterns, and the extent to which U. umgibeli n. sp. is restricted to its current known host-copepod system.
Conclusion
The discovery of U. umgibeli n. sp. provides a new record of the genus from the southwestern Indian Ocean and contributes to a sparse African record. By combining detailed morphometrics with molecular data, this study demonstrates that integrative taxonomy is essential for resolving species boundaries in morphologically conservative epibiotic parasites and for understanding their evolution and distribution in marine ecosystems. Udonella umgibeli n. sp. is morphologically distinct, supported by current separation in geography and host-copepod association. Additional loci and broader sampling will refine future udonellid phylogenetic analyses and will be especially valuable for testing whether U. umgibeli represents a locally restricted endemic or part of a wider southwestern Indian Ocean radiation of Udonella associated with regional caligid diversity.
Data availability statement
The sequence data generated in this study have been made available on the NCBI GenBank database, voucher specimens have been accessioned in the Parasite Collection at the National Museum (NMB) in Bloemfontein, South Africa, and all other data are included in the manuscript.
Acknowledgements
We would like to thank the Unit for Environmental Sciences and Management, North-West University (NWU), for the use of field equipment and laboratory facilities. Additional thanks go to our colleagues Hannes Erasmus, Anja Erasmus and Anja Vermaak from the NWU, who assisted with the collection of samples. This is contribution no. 1064 from the NWU-Water Research Group.
Author contributions
L.D.K.: Methodology, Investigation, Funding acquisition, Writing – original draft.
I.P.: Methodology, Writing – review & editing.
N.J.S.: Conceptualization, Data curation, Funding acquisition, Writing – review & editing.
Financial support
This work is based on the research supported by the National Research Foundation (NRF) of South Africa (grant 132805; L. van der Spuy; grant CPRR240430216732; N.J. Smit). Opinions, findings, conclusions and recommendations expressed in this publication are those of the authors, and the NRF accept no liability whatsoever in this regard.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical standards
Ethical approval for this project was obtained from the Animal Care Research Ethics Committee of North-West University (NWU-00565-19-A5). Permits for the collection of A. honckenii specimens were issued by the South African Department of Agriculture, Forestry and Fisheries (permit nos.: RES2022-44; RES2022-86 and RES2025-16) and Cape Nature (permit no. CN44-87-18289).

 Open access
Open access