



Introduction
Sexual competition, which leads to sexual selection, exerts significant pressure within parasite populations. It shapes their sexual size dimorphism (Piross et al., Reference Piross, Harnos and Rózsa2019) and impacts both primary and secondary sexual traits (Tryjanowski et al., Reference Tryjanowski, Adamski, Dylewska, Bulkai and Rózsa2009; Rózsa et al., Reference Rózsa, Tryjanowski, Vas, Morand, Krasnov and Littlewood2015) and the sex ratio (SR, here defined as the proportion of males expressed on a 0–1 scale), which is a key factor influencing the strength of sexual selection pressure.
Following Düsing (Reference Düsing1884), Ronald Fisher proposed an idea that has become a fundamental hypothesis in evolutionary biology (Gardner, Reference Gardner2023). He explained that in a sexually reproducing population, the optimal strategy for parents is to allocate half of their resources into producing male offspring and half of their resources into producing females. This strategy yields a characteristic equal SR (= 0.5) in several species (Fisher, Reference Fisher1930). However, deviations from this equality do occur under specific conditions.
In free-living organisms, Local Mate Competition (LMC) is 1 factor potentially affecting primary SRs. Primary, secondary, and tertiary SRs are measured at the zygote, egg and adult stages, respectively (West, Reference West2009). LMC occurs when a metapopulation is subdivided into small subpopulations, where the offspring (brothers, sisters and other closely related kin) form small and temporarily isolated breeding communities, the members of which mate with each other before dispersing to other habitat patches to establish new subpopulations (Hamilton, Reference Hamilton1967; Charnov, Reference Charnov1982). Under such conditions, parents can maximize the number of their grandchildren by reducing the proportion of the more competitive sex – usually the male (Janicke et al., Reference Janicke, Häderer, Lajeunesse and Anthes2016) – in their offspring. Indeed, 1 son (or very few) is sufficient to fertilize all their sisters.
Although the interpretive framework of LMC was at the individual level, Hamilton (Reference Hamilton1967) added a footnote to link it to group selection. Subsequently, Bulmer and Taylor (Reference Bulmer and Taylor1980) and Frank (Reference Frank1986) interpreted the phenomenon in the context of the ‘haystack model’. The metaphor, which names a model of multilevel selection, was introduced by Maynard Smith (Reference Maynard Smith1964). Imagine a population of mice divided into subgroups inhabiting haystacks. Within-sex competition occurs at 2 levels: typically, competition between genetically related males dominates within the group, while genetically non-related females compete with each other when invading new habitat patches to found new colonies. In this model, intrasexual competition occurs in both sexes, but only male-male competition happens among close kin. Therefore, LMC will select parents to decrease the proportion of males in their litters.
Note that Gardner and Hardy (Reference Gardner and Hardy2020) argued for an opposite direction of causality, emphasizing that LMC drives SR evolution, and then the observed inbreeding is a result of that local structure, and not the primary cause of it. Other authors, however, agree with the originally proposed direction of causality, i.e. inbreeding causes LMC (Yamauchi and Kobayashi, Reference Yamauchi and Kobayashi2011; Bonifacii and Field, Reference Bonifacii and Field2024).
In parasitological terminology, subpopulations are called ‘infrapopulations’ (equals the intensity of infestation, the number of conspecific parasites living on the same host (Bush et al., Reference Bush, Lafferty, Lotz and Shostak1997)) while a metapopulation refers to all individuals of a parasite species that live on different, spatially separated host populations, or live on different host species. Avian lice (Insecta: Psocodea: Phthiraptera) complete their entire life cycle on the host body surface. The life cycle is typically 2–4 months long (Johnson and Clayton, Reference Johnson, Clayton, Price, Hellenthal, Palma, Johnson and Clayton2003), and they primarily, but not exclusively, disperse to new host individuals during the breeding season, either between sexual partners (Hillgarth, Reference Hillgarth1996) or from parents to nestlings (Clayton and Tompkins, Reference Clayton and Tompkins1994). Thus, lice often complete several life cycles on the same host before transmission, strongly predisposing them to inbreeding. Males are the more competitive sex in avian lice (Piross et al., Reference Piross, Harnos and Rózsa2019); therefore, a female bias is expected to arise under LMC pressure.
Hamilton (Reference Hamilton1967) used haplodiploid wasps to exemplify LMC, where females can evidently control the sex of their offspring, although his theory is not restricted to haplodiploids. It is based on frequency-dependent selection and a maternal ability to control offspring SR, which may involve other mechanisms as well. Nevertheless, note that the sex determination system of parasitic lice is likely to be similar (although not entirely identical) to haplodiploidy. In their close relatives, the liposcelid booklice (Psocoptera) (Hodson et al., Reference Hodson, Hamilton, Dilworth, Nelson, Curtis and Perlman2017) and in the human body lice (Pediculus humanus ‘ecotype humanus’) (McMeniman and Barker, Reference McMeniman and Barker2006; de la Filia et al., Reference de la Filia, Andrewes, Clark and Ross2018), sex is determined by paternal genome elimination (PGE) suggesting that it is general in parasitic lice. This system is effectively similar to haplodiploidy in the sense that males silence their paternally inherited genes and, thereby, they are functionally haploid. In haplodiploids, and also in the functionally similar PGE systems, natural selection favours producing fewer sons and more daughters especially under LMC pressure (Harpur et al., Reference Harpur, Sobhani and Zayed2013; Harper et al., Reference Harper, Bagley, Thompson and Linnen2016).
Several authors have formerly observed female-biased SRs in parasitic lice and often attributed it to a reduced lifespan of males (Buxton, Reference Buxton1941; Baum, Reference Baum1968; Nelson, Reference Nelson and Linsley1972). Rothschild and Clay (Reference Rothschild and Clay1953) suggested that the phenomenon may be caused by sampling bias; males are the smaller-bodied sex, which are more easily overlooked during collection. Three decades ago, Rózsa et al. (Reference Rózsa, Rékási and Reiczigel1996) compared the SRs in 2 genera of lice from 2 species of Corvidae (Aves: Passeriformes). They found that infrapopulations tend to be more female-biased in smaller infrapopulations and more equal in larger ones. Using a historical data set published by Buxton (Reference Buxton1941), Rózsa (Reference Rózsa1997) demonstrated that the same positive relationship between SR and infestation intensity (infrapopulation size, the number of adult lice inhabiting the same host) was also found in human head lice (Pediculus humanus ‘ecotype capitis’). Both papers interpreted this relationship as follows: small size of infrapopulations led to high levels of inbreeding, which in turn resulted in LMC and a decrease in SRs. Conversely, the large size of infrapopulations led to outbreeding, and the proportion of males increased until it reached the Fisherian equilibrium (SR = 0.5).
In response to Rózsa’s (Reference Rózsa1997) research note on human head lice, Perotti et al. (Reference Perotti, Catalá, Ormeño, Żelazowska, Biliński and Braig2004) published a paper that appeared to falsify its results. First, they gathered data on head lice infestations from schoolchildren, where they found no correlation between infestation intensity and SR. This is not at all surprising, since the intensity varied between 1 and 3000 in Buxton’s dataset, but only between 1 and 40 in Perotti et al. (Reference Perotti, Catalá, Ormeño, Żelazowska, Biliński and Braig2004). Covariation detected in a wide range of variability cannot necessarily be observed in a range of 2 orders of magnitude narrower. More importantly, Perotti et al. (Reference Perotti, Catalá, Ormeño, Żelazowska, Biliński and Braig2004) were the first to document the presence of Wolbachia sp. infections in head lice and they proposed that the female-bias of most samples of head lice might be caused either by these bacteria or by some other SR distorting bacterial infections. Wolbachia infections are transmitted maternally, and therefore, have an adaptive benefit in manipulating the hosts to produce exclusively female offspring (Werren, Reference Werren1997; Stouthamer et al., Reference Stouthamer, Breeuwer and Hurst1999). There are various physiological mechanisms through which Wolbachia can shift their hosts’ primary or secondary SRs. Some Wolbachia lineages apparently do not affect host SRs, while infections by other lineages render several arthropod populations or species entirely parthenogenetic (Hyder et al., Reference Hyder, Lodhi, Wang, Bukero, Gao and Mao2024).
Studying human head lice, Lang (Reference Lang1976) noted a positive covariation between infestation intensity and SR in some, but not all, populations. He ascribed shortened female longevity under crowded conditions to the aggressive copulatory attempts by males, which are harmful to newly emerged females not fully sclerotized before copulation and increase their mortality. Perotti et al. (Reference Perotti, Catalá, Ormeño, Żelazowska, Biliński and Braig2004) unified the aforementioned 2 ideas, proposing that bacterial distortion of primary or secondary SR was responsible for the female bias typical for small louse infrapopulations (which are common), and this bias was shifted towards higher tertiary (adult) SRs due to male copulatory aggression in large, crowded infrapopulations (which are rare). Hereafter, the term ‘Wolbachia + Crowding Hypothesis’ is used to refer to this hypothesis.
When Rózsa et al. (Reference Rózsa, Rékási and Reiczigel1996) and Rózsa (Reference Rózsa1997) proposed that LMC was responsible for the primary SR bias in lice, they referred to its supposed adaptive value. In contrast, when Perotti et al. (Reference Perotti, Catalá, Ormeño, Żelazowska, Biliński and Braig2004) proposed that infections with SR distorting bacteria and copulations harmful to females might have caused the SR bias, they referred to physiological and physical mechanisms of its manifestation in the zygote or embryonic stages (due to Wolbachia infections), and in the adult stages (due to sexual aggression). This latter hypothesis does not presume that biased SRs have adaptive value for lice. Thus, both the LMC and the Wolbachia + Crowding Hypotheses predict a strong female bias in small infrapopulations and an increasing proportion of males in larger ones. However, the predicted extent of SR bias differs between the 2 hypotheses.
Wolbachia infections (or other SR distorting bacteria) can cause arthropod populations and even species to become fully parthenogenetic (Hyder et al., Reference Hyder, Lodhi, Wang, Bukero, Gao and Mao2024), although many infected species continue to reproduce sexually. Fifteen species of avian lice have been shown to host Wolbachia (Kyei-Poku et al., Reference Kyei-Poku, Colwell, Coghlin, Benkel and Floate2005; Covacin and Barker, Reference Covacin and Barker2007; Mahmood et al., Reference Mahmood, Nováková, Martinů, Sychra and Hypša2023), none of which are parthenogenetic. Thus, the Wolbachia + Crowding Hypothesis would allow for the finding of at least some parthenogenetic (SR = 0) louse populations or species, although it is not necessarily expected.
Further, Lang (Reference Lang1976) and Perotti et al. (Reference Perotti, Catalá, Ormeño, Żelazowska, Biliński and Braig2004) presumed that mating occurs more frequently in larger infrapopulations, leading to increased female mortality due to the more frequent harmful interactions, which is an integral part of the Wolbachia + Crowding Hypothesis. If larger infrapopulations lead to higher female mortality and thus an increasing proportion of males, aggressive copulation is expected to increase. Logically, this would create a self-reinforcing cycle that can ultimately eliminate all females (SR = 1). Overall, this hypothesis predicts a positive covariation between intensity and SR, where SR may vary from 0 (a possible value) to 1 (a necessary value).
In contrast, Hamilton’s LMC Hypothesis posits that parents produce few (but always more than zero) male offspring in strongly inbred infrapopulations. There are 2 reasons to presume that small infrapopulations tend to be more inbred than large ones. First, genetic drift exerts a stronger pressure to homogenize their genetic composition than that of the large ones. Second, small infrapopulations are more likely to originate from single infestations, while multiple infestations originating from different sources reduce inbreeding and also are more likely to yield larger infrapopulation size. Thus, LMC predicts a low, but non-zero proportion of males in small populations, and the Fisherian equilibrium (SR = 0.5) in large infrapopulations.
The aim of the present study is to examine the SRs of ricinid lice (Psocodea: Phthiraptera: Ricinidae) on hummingbirds (Apodiformes: Trochilidae), with particular attention to the magnitude of female bias and its relationship to infrapopulation size (infestation intensity). Our goal is to assess how the observed patterns correspond to predictions of the 2 hypotheses described earlier.
Materials and methods
The study species
Hummingbirds are small-bodied (mostly 2–24 g), nectarivorous and partly insectivorous birds distributed over the Western Hemisphere. They are known to be infested by 4 genera of lice (Oniki-Willis et al., Reference Oniki-Willis, Willis, Lopes and Rózsa2023), 2 of which are extremely rare and not included in the present study. On the other hand, Trochiloecetes spp. and Trochiliphagus spp. (Psocodea: Phthiraptera: Ricinidae) are typically more prevalent (prevalence is the proportion of infested hosts) and relatively large-bodied lice (relative to host size), with a body length of adults ranging from 1.55 to 3.35 mm (Harnos et al., Reference Harnos, Lang, Petrás, Bush, Szabó and Rózsa2017). As in most species of parasitic lice, males are smaller-bodied than females. Their infrapopulation size (intensity of infestation: the number of parasites per infested host) tends to be low; typically, only a few individuals can be found on an infested bird, and rarely a few dozen of them. Males are often absent in small infrapopulations, and it seems likely – though unproven – that females can reproduce parthenogenetically under such conditions (Nelson, Reference Nelson and Linsley1972). Due to their low intensities and relatively large body sizes, ricinid lice can be reliably sexed and counted by collectors. Collections are not necessarily exact; some individuals may be lost (Clayton and Drown, Reference Clayton and Drown2001). Compared to other avian lice, however, this error is relatively small in the present case because the hosts are relatively small and the ricinid lice are relatively large. Early authors noted that ricinids’ SRs exhibit an exceptionally strong female bias, with males often constituting only about 10% of the samples (Eichler, Reference Eichler1953; Carriker, Reference Carriker1964; Rheinwald, Reference Rheinwald1968; Nelson, Reference Nelson and Linsley1972), making them an excellent subject for parasite SR studies. Note that there is no published information regarding Wolbachia infections in ricinid lice.
Hummingbirds were identified to the species level; no subspecies were assigned. Their taxonomy and nomenclature follow Gill et al. (Reference Gill, Donsker and Rasmussen2025). Due to uncertainties of the species concept in parasitology (Mey, Reference Mey1998; Gustafsson and Najer, Reference Gustafsson and Najer2022), the taxonomy and nomenclature of many ricinid lice are not well resolved at the species level. Therefore, only genus-level louse names were applied, and their species identity was not considered. In this study, we follow Price et al. (Reference Price, Hellenthal, Palma, Price, Hellenthal, Palma, Johnson and Clayton2003) to recognize the genus Trochiliphagus, in contrast to Rheinwald (Reference Rheinwald2007), who synonymized this genus with the genus Ricinus.
Note that our data were obtained from 2 separate surveys that were neither coordinated nor specifically designed for the objectives of the present study.
South American samples
More than 17 000 hummingbirds representing several species were mist-netted, ringed and deloused from 1977 to 2002 in Brazil by YO-W and EOW. The collection sites were located in the states of Espirito Santo, São Paulo, Minas Gerais, Rio de Janeiro, Mato Grosso, Santa Catarina, Amazonas and Pará, in decreasing order of sample size. A preliminary and almost complete list of localities (with forest types and coordinates) was formerly published (Oniki-Willis and Willis, Reference Oniki-Willis and Willis2018). Further localities were in Santa Teresa, Museu de Biologia Mello Leitão and the Biological Reserve Augusto Ruschi; both in Espirito Santo State. While gently blowing on the birds’ plumage to separate the feathers, lice were examined on the birds with the naked eye and then collected with pointed tweezers. Carefully examining non-infested birds usually took about 2 min, while searching and collecting lice from infested hosts took about 6–12 min. The samples were stored in 70% ethanol and identified in the laboratory under a stereomicroscope. Only host–parasite species pairs with sufficiently large number of host individuals (>10, an arbitrary limit) infested by at least 1 adult louse were included in the analysis; smaller samples were excluded. Choosing this particular arbitrary threshold did not affect the present results qualitatively. Due to ongoing taxonomic work based on these specimens, vouchers have not yet been deposited, except for slides containing samples of Trochiloecetes naevius, which were donated to the entomology collection of the Smithsonian Institution (Washington, DC, USA), the Department of Plant Industry (Gainesville, Florida, USA), and to Bernard Nelson in California (USA).
North American samples
Seventy-seven dead specimens of Ruby-throated Hummingbirds (Archilochus colubris) were examined from 1991 to 2023 in Canada by TG. All except 1 originated from Manitoba; 1 specimen was from Kenora, Ontario (about 200 km east of Winnipeg, Manitoba). Birds were salvaged from local wildlife rehabilitation hospitals (Wildlife Haven and Prairie Wildlife Rehabilitation Centre) and washed consecutively, twice in soapy water and once in clear water. Wash-water was passed through a 90-µm sieve, and the residues were sorted under a stereomicroscope. A total of 28 adult Trochiloecetes sp. were collected and preserved in 70% or 95% ethanol. Vouchers are deposited in the J.B. Wallis/R.E. Roughley Museum of Entomology, Department of Entomology, University of Manitoba. Trochiliphagus sp. did not occur in these samples.
Prevalence and intensity
Only adults (imagoes) were included to calculate prevalence (the proportion of infested birds) and mean intensity (the mean number of adult lice per infested bird). Therefore, the prevalence and mean intensity values provided in Table 1 are not directly comparable to those obtained from other studies. Unfortunately, differentiating late third-stage nymphs versus adults is not always easy in some ricinid lice. For the South American samples, YO-W and EOW introduced the category of ‘subadult’ lice, which they applied to lice smaller and lighter than adults but already possessing visible genitalia in the abdomen. It was not clear whether these subadults were late third-stage nymphs or teneral adults (or both). To ensure comparability across samples and to reduce uncertainty, these subadult specimens were excluded from all analyses. Specimens collected in North America had been preserved in ethanol prior to microscopic examination, and adults were distinguished from nymphs and sexed accurately.
Basic sample characteristics and Type 1 linear regression models of the relationship between log(intensity) and sex ratio (proportion of males) of intensity pools in 10 host–parasite species pairs

Table 1 Long description
The table summarizes, for 10 hummingbird host–louse species pairs, sample sizes, infestation prevalence, intensity ranges and means, observed male proportion in pooled infestations, and a linear model linking log-transformed intensity to sex ratio. Slopes are positive for all pairs, indicating higher intensity is associated with a higher proportion of males. Five pairs show statistically significant positive relationships: Chlorostilbon lucidus with Trochiloecetes, Hylocharis chrysura with Trochiliphagus, Hylocharis chrysura with Trochiloecetes, Leucochloris albicollis with Trochiloecetes, and Florisuga fusca with Trochiloecetes; Archilochus colubris with Trochiloecetes is also significant but based on small samples. The strongest evidence appears for Leucochloris albicollis with Trochiloecetes, with many hosts and pools and a very small p-value. Sample sizes vary widely, from 11 to 505 hosts and from 4 to 30 pools, and prevalence ranges from about 0.06 to 0.44. Mean intensity spans roughly 1.7 to 7.9 lice per infested host, with the widest intensity range reaching 53 in Leucochloris albicollis with Trochiloecetes. Sex ratios range from about 0.19 to 0.45 across pairs, so even where trends are significant, male proportion remains below one half. Interpretation should consider that several pairs have few pools, which can limit precision and make non-significant results harder to detect.
N host is the number of infested host individuals, N pool is the number of infestation pools (after hosts with equal infestation intensity were pooled together), prevalence is the proportion of infested birds, intensity is the number of adult lice on an infested bird, and P is the significance of deviation from the horizontal (bold numbers indicate significance <0.05).
Merging similar-sized infrapopulations into intensity pools
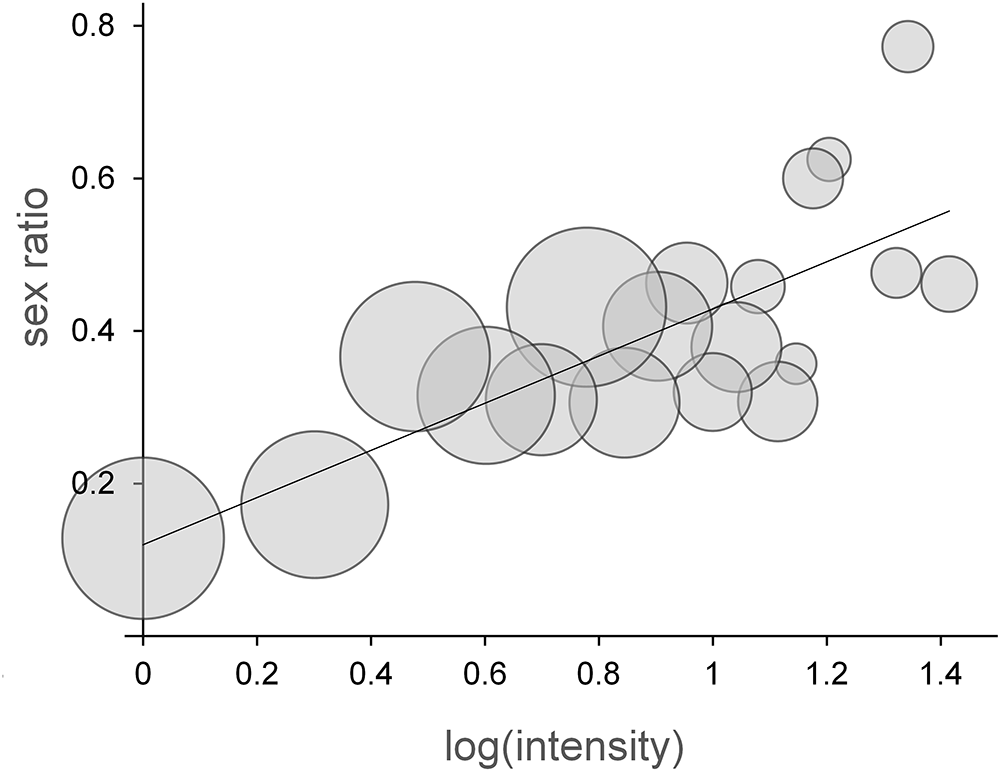
In each host–parasite species pair, infrapopulations of the same size (i.e. lice from infestations of the same intensity) were merged into a single intensity pool. Taking the largest sample as an example (Trochiloecetes sp. collected from the Black Jacobin, Florisuga fusca), there were 87 birds each infested by 2 adult lice. These infrapopulations were united into an intensity pool (which had a total of 174 adult lice) to calculate the SR belonging to this particular intensity value (intensity = 2; SR = 0.172, see also Figure 1). In the case of low intensities (1, 2, etc.), this meant that several merged infrapopulations formed 1 pool, because there were many small infrapopulations having the same intensity value. In the case of high intensities (which were very rare), usually only 1 infrapopulation formed a pool, because no other infrapopulation had the same intensity. Thus, roughly the same number of lice belonged to the low-intensity vs the large-intensity pools. Consequently, there was no need to perform statistical control for the size (or ‘weight’) of pools (see Figure 1).
The relationship between log(intensity) and sex ratio (proportion of males) of intensity pools in the largest sample of the present study, Trochiloecetes sp. infestations of the Black Jacobin (Florisuga fusca). The circles are proportional to the number of lice constituting each intensity pool. The line represents the Type 1 linear regression model of the relationship. See statistical details in Table 1.

Figure 1 Long description
The bubble chart illustrates the relationship between log(intensity) and sex ratio. The horizontal axis is labeled log(intensity), ranging from 0 to 1.4. The vertical axis is labeled sex ratio, ranging from 0 to 0.8. Bubble size represents the number of lice in each intensity pool. A fitted line with a positive slope indicates that the sex ratio generally increases as log(intensity) increases. Notable data points include a low sex ratio near (0, 0.15) and a high sex ratio near (1.4, 0.75). There are no color-coded categories or groupings. The bubbles are scattered, with larger bubbles clustered towards higher log(intensity) values, suggesting higher numbers of lice in those pools. The line represents a Type 1 linear regression model of the relationship.
Statistics
Intensity values of the pools were log10-transformed. SR was defined as the proportion of males within an intensity pool, expressed on the 0–1 scale. The relationships between log(intensity) (as a predictor variable) and SR (as a predicted variable) were analysed using Type 1 linear regressions over the pools; all tests were 2-tailed.
Results
Number of species and individuals
The South American sample included 9 host–parasite species pairs with sufficiently large numbers of infested hosts (N host > 10), with a total of 1051 infested birds and 3995 adult lice. Chrysuronia versicolor, Chlorostilbon lucidus, Thalurania glaucopis and Florisuga fusca with Trochiloecetes spp. infestations, Eupetomena macroura with Trochiliphagus spp. infestations, Hylocharis chrysura and Leucochloris albicollis with both Trochiloecetes and Trochiliphagus spp. infestations were included. Refer to Table 1 for basic sample characteristics including the number of infested hosts, prevalence, mean intensity and SR. The North American sample included 77 specimens of Ruby-throated Hummingbirds (Archilochus colubris). Eleven birds hosted a total of 28 adult Trochiloecetes sp.
Relationship between infestation intensity (infrapopulation size) and SR
Infrapopulation size (infestation intensity) varied from 1 (by definition) to 53 adult lice per bird (the latter value refers to 1 Trochiloecetes-infested Leucochloris albicollis). The frequency distributions of host individuals among intensity categories exhibited the characteristic aggregated shape (Crofton, Reference Crofton1971). Species-level SRs varied from 0.185 to 0.452 (Table 1), with a considerable variability within species. This variation was related to infrapopulation size. Overall, the proportion of males was often about 5–15% at low levels of intensity (say, at 1–2 adult lice per bird) and about 50% at the highest levels of intensity (dozens of adult lice per bird).
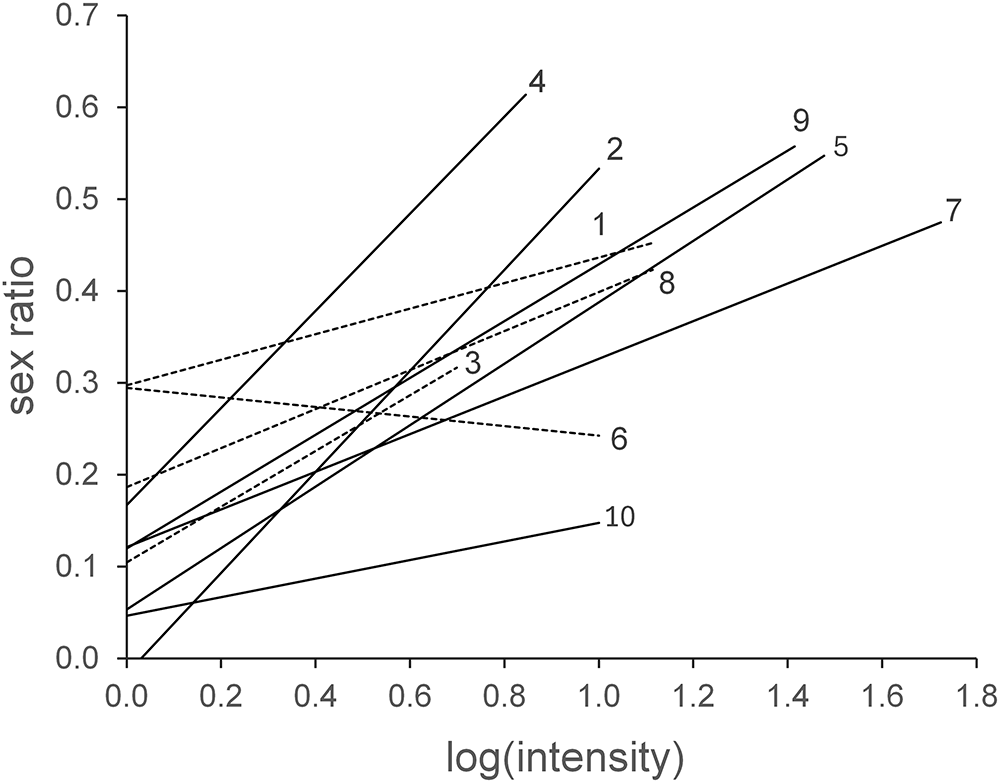
Highly significant positive interactions (P < 0.001) were observed between parasite infrapopulation size and SR in the Trochiloecetes infestations of Leucochloris albicollis and Florisuga fusca. Note that these 2 samples had the highest numbers of infested birds and intensity pools. Additionally, significant positive interactions (P < 0.05) were identified in the Trochiliphagus infestations of Hylocharis chrysura, as well as in the Trochiloecetes infestations of Hylocharis chrysura, Chlorostilbon lucidus and Archilochus colubris (the latter was the North American sample). Visual representation and statistical details are provided in Figure 2 and Table 1.
Type 1 linear regression models of the relationship between log(intensity) and sex ratio (proportion of males) of intensity pools in 10 host–parasite species pairs. Continuous lines indicate significant, dashed lines non-significant relationships (P = 0.05, 2-tailed). The numbers refer to the species pairs named in Table 1. Note that regression line No. 9 represents Florisuga fusca, the sample already visualized with intensity pools on Figure 1.

Figure 2 Long description
The x-axis is labeled log(intensity) and ranges from 0.0 to 1.8. The y-axis is labeled sex ratio and ranges from 0.0 to 0.7. Sex ratio represents the proportion of males. Each of the 10 lines is numbered 1 through 10, corresponding to distinct host-parasite species pairs. Continuous lines indicate significant relationships and dashed lines indicate non-significant relationships at the 0.05 level, 2-tailed. The majority of lines show a positive association between log(intensity) and sex ratio. Line 4 begins near (0.0, 0.3) and rises steeply, reaching approximately (0.9, 0.6), making it the steepest among all lines. Line 2 starts near (0.0, 0.35) and rises to approximately (1.0, 0.55). Line 1 begins near (0.0, 0.3) and reaches approximately (1.0, 0.5). Line 9 starts near (0.8, 0.4) and extends to approximately (1.6, 0.6). Line 5 begins near (0.8, 0.35) and rises to approximately (1.8, 0.65). Line 7 starts near (1.0, 0.3) and reaches approximately (1.8, 0.65). Line 6 begins near (0.4, 0.25) and rises gradually to approximately (1.2, 0.45). Line 3 starts near (0.0, 0.25) and rises slowly to approximately (1.2, 0.4). Line 8 begins near (0.4, 0.15) and rises to approximately (1.4, 0.3). Line 10 is the flattest, starting near (0.2, 0.1) and reaching only approximately (1.4, 0.2). Most lines are continuous, indicating significant relationships. A smaller number are dashed, indicating non-significant relationships. Lines 4, 2, 9 and 5 show the steepest positive slopes. Lines 8 and 10 show the weakest positive slopes and remain in the lower range of sex ratio values throughout.
Discussion
This is the most substantial evidence to date, based on the largest number of species and individuals, to document the positive relationship between infestation intensity and SR in parasitic lice. The direction of these correlations appears to correspond to the predictions of both the Wolbachia + Crowding Hypothesis and the LMC Hypothesis. However, considering the different ranges of SR variability predicted by these 2 hypotheses, the observed patterns align more closely with the LMC Hypothesis, which predicts low but non-zero SRs for small infrapopulations and an equal SR for large ones.
Admittedly, these samples were obtained from monitoring studies that were not explicitly designed for the present study. Different collectors, using different collection methods at different times and on different continents, gathered samples of varying sizes. The fact that South and North American hummingbird lice still exhibited a similar relationship between infestation intensity and SR signifies that these results are not dependent on particular species or details of collection methodology.
It is reasonable to assume that sexual competition between closely related males occurs within infrapopulations, especially within small ones. Furthermore, it can be assumed that the transmitting individuals are usually females, partly because their numbers exceed those of males. However, there is no reason to presume competition for opportunities to transfer to new hosts. In the ricinid-hummingbird system, this level of competition is negligible for 2 reasons. First, prevalence is low; more than 80% of birds are not infested (see Table 1). Second, even the infested ones tend to harbour only 1 or a very few lice, while heavy infestations (where competition might be expected (Bush and Malenke, Reference Bush and Malenke2008)) are rare. Thus, females do not limit each other’s chances for transmission and subsequent invasion success. Using Maynard Smith’s (Reference Maynard Smith1964) haystack metaphor, female mice will not compete for new haystacks to invade when >80% of the neighbouring haystacks are empty, and most of the remaining ones are almost empty (having only 1–2 rival mice living in them). Accordingly, males are the more competitive sex in lice, and it is adaptive for the parents to reduce their proportion in the offspring under LMC pressure, i.e. in small subpopulations.
Nevertheless, LMC and the Wolbachia + Crowding Hypothesis are mutually non-exclusive. The latter refers to the mechanisms by which different SRs may emerge, whereas the LMC refers to the adaptive value of SRs under different conditions. One cannot rule out the possibility that louse SRs – which are adaptive from a Hamiltonian perspective – result from cooperation between lice and the SR distorting bacteria that infect them. It is possible that both hypotheses are valid to some degree, influencing SRs in lice simultaneously. If this is the case, SR distortion in ricinid lice can be viewed as a mutualistic aspect of their relationship with the bacteria that infect them.
Note that the presence of Wolbachia sp. (or other SR distorting bacteria) infections in ricinid lice is still hypothetical; future molecular studies may verify their occurrence in these ectoparasites. Future molecular studies should also test whether levels of inbreeding truly differ between small vs large infrapopulations. Another possible way to differentiate between the 2 hypotheses would be to identify the timing of SR distortion within the louse developmental cycle. LMC implies that louse parents adjust the primary SR of their offspring so that the relationship between infrapopulation size and SR appears in the egg and nymphal (larval) stages. On the other hand, the Wolbachia + Crowding Hypothesis implies that the female bias in the primary or secondary SRs shifts towards equality (in large infrapopulations) only in the tertiary SR of the adult phase. Therefore, future studies utilizing molecular techniques (Bressa et al., Reference Bressa, Papeschi and Toloza2015; Hodson et al., Reference Hodson, Hamilton, Dilworth, Nelson, Curtis and Perlman2017) to determine the genetic sex of eggs or nymphal stages are expected to provide more conclusive insights into the mechanisms driving the extraordinary SR patterns observed in parasitic lice.
Data availability statement
The datasets used and analysed above are available from the corresponding author upon reasonable request.
Acknowledgements
Y. O-W. and E. O. Willis are grateful for a fellowship from the Brazilian CNPq (Conselho Nacional de Pesquisa e Desenvolvimento Tecnológico), the National Geographic Society, Volkswagen Foundation, FAPESP (Fundação de Amparo à Pesquisa de São Paulo), and Aracruz Florestal S. A. for conducting field work in various states in eastern Brazil throughout the years. International Council for Bird Preservation, Pan American Section (ICBP), U.S. Fish and Wildlife Service and Aracruz Florestal S.A. helped acquiring the special bands for hummingbirds, used in 1993 and subsequent years. CEMAVE (Centro Nacional de Pesquisa e Conservaçao de Aves Silvestres) provided permission for banding birds. We appreciate help from the directors of various biological reserves, national parks, farm owners who provided permission and logistical support, and workers at the Museu de Biologia ‘Mello Leitão’ at Santa Teresa, ES, who saved banded hummingbirds found dead. We also thank Mr. José Batista and family, who provided lodging and monitored visiting hummingbirds in their yard in Avaré. TG thanks the Department of Entomology and the Faculty of Agricultural and Food Sciences at the University of Manitoba for their continued support, and Canadian Wildlife Service, Environment Canada for permits to possess and examine hummingbirds for ectoparasites. TG also thanks the staff at Wildlife Haven and Prairie Wildlife Rehabilitation Centre for the care with which they handled and processed birds for this study.
LR used Grammarly (https://app.grammarly.com/) as an AI tool to enhance the grammar and fluency of the text.
We appreciate our reviewers for their time and effort in enhancing this article.
Author contributions
LR and OS conceptualized the study. YO-W, EOW and TG conducted fieldwork and data gathering. The first manuscript draft was written by LR. All authors, except EOW, read, edited, reviewed and approved of the final manuscript.
Financial support
This work was supported by the Hungarian National Research, Development, and Innovation Office under contracts RRF-2.3.1-21-2022-00006 and K143622, by the Slovak Research and Development Agency under contract APVV-22-0440 (LR), and by Discovery Grants from the Natural Sciences and Engineering Council of Canada (TG).
Competing interests
The authors declare no conflicts of interest.
Ethical standards
Not applicable.


 Open access
Open access