



Introduction
Gastrointestinal protist (GIP) infections remain a public health concern in sub-Saharan Africa as major contributors to the burden of diarrhoeal disease (Chu et al., Reference Chu, Yang, Yang, Liang, Liu, Chen, Wang, Zhou and Wang2024). Because GIPs are faecal-orally transmitted through ingestion of contaminated water or fresh produce or via direct contact with infected individuals or animals, they are closely linked with poverty and inadequate household and sanitary infrastructures (Mertens et al., Reference Mertens, Arnold, Benjamin-chung, Boehm, Brown, Capone, Clasen, Fuhrmeister, Grembi, Holcomb, Knee, Kwong, Lin, Luby, Nala, Nelson, Njenga, Null, Pickering, Rahman, Reese, Steinbaum, Stewart, Thilakaratne, Cumming, Colford and Ercumen2023). Giardia duodenalis (syn. G. intestinalis and G. lamblia) and Cryptosporidium spp. are the most relevant GIP among African children, with estimated pooled prevalence rates of 18.3% (95% CI: 16.5–20.2) and 2.5% (95% CI: 1.8–3.2), respectively (Hajissa et al., Reference Hajissa, Islam, Sanyang and Mohamed2022; Kalavani et al., Reference Kalavani, Matin, Rahmanian, Meshkin, Bahadori Mazidi, Taghipour and Abdoli2024). Giardia duodenalis infections are rarely fatal in these regions. Data from meta-analyses (Muhsen and Levine, Reference Muhsen and Levine2012; Baker et al., Reference Baker, Hasso-agopsowicz, Pitzer, Platts-Mills, Peralta-santos, Troja, Archer, Guo, Sheahan, Lingappa, Jit and Lopman2021) and prospective and case-control studies (Kotloff et al., Reference Kotloff, Nataro, Blackwelder, Nasrin, Farag, Panchalingam, Wu, Sow, Sur, Breiman, Faruque, Zaidi, Saha, Alonso, Tamboura, Sanogo, Onwuchekwa, Manna, Ramamurthy, Kanungo, Ochieng, Omore, Oundo, Hossain, Das, Ahmed, Qureshi, Quadri, Adegbola, Antonio, Hossain, Akinsola, Mandomando, Nhampossa, Acácio, Biswas, O’reilly, Mintz, Berkeley, Muhsen, Sommerfelt, Robins-browne and Levine2013; Breurec et al., Reference Breurec, Vanel, Bata, Chartier, Farra, Favennec, Franck, Giles-vernick, Gody, Luong, Onambélé, Rafaï, Razakandrainibe, Tondeur, Tricou, Sansonetti, Vray and Ryan2016; Rogawski et al., Reference Rogawski, Bartelt, Platts-Mills, Seidman, Samie, Havt, Babji, Trigoso, Qureshi, Shakoor, Haque, Mduma, Bajracharya, Gaffar, Lima, Kang, Kosek and Ahmed2017) indicate that early-life G. duodenalis infections are not associated with diarrhoea but are linked to undernutrition and cognitive impairment. In contrast, Cryptosporidium spp. infections can cause a wide diversity of clinical manifestations ranging from asymptomatic to life-threatening depending on host nutritional and immunologic status (Khalil et al., Reference Khalil, Troeger, Rao, Blacker, Brown, Brewer, Colombara, De Hostos, Engmann, Guerrant, Haque, Houpt, Kang, Korpe, Kotloff, Lima, Wa, Platts-Mills, Shoultz, Forouzanfar, Hay, Reiner and Mokdad2018). Indeed, cryptosporidiosis is the second most common cause of moderate-to-severe diarrhoea, following the rotavirus pathogen, in children under 2 years of age in sub-Saharan Africa (Kotloff et al., Reference Kotloff, Nataro, Blackwelder, Nasrin, Farag, Panchalingam, Wu, Sow, Sur, Breiman, Faruque, Zaidi, Saha, Alonso, Tamboura, Sanogo, Onwuchekwa, Manna, Ramamurthy, Kanungo, Ochieng, Omore, Oundo, Hossain, Das, Ahmed, Qureshi, Quadri, Adegbola, Antonio, Hossain, Akinsola, Mandomando, Nhampossa, Acácio, Biswas, O’reilly, Mintz, Berkeley, Muhsen, Sommerfelt, Robins-browne and Levine2013), a situation further aggravated by the current absence of effective vaccines for humans (Rahman et al., Reference Rahman, Mi, Zhou, Gong, Ullah, Huang, Han and Chen2022). Balantioides coli (formerly known as Balantidium coli or Neobalantidium coli) is the only ciliated protozoan parasite able to infect humans. Human balantidiasis, while often asymptomatic, can lead to severe dysenteric illness in immunocompromised individuals (Barbosa et al., Reference Barbosa, Dib, Uchôa and Liu2018). It is regarded as an emerging public health concern in some African regions, especially in communities in close contact with pigs or using untreated water. However, its epidemiology is largely unknown because of a lack of routine diagnostic screening and underreporting (da Silva et al., Reference da Silva, Dib, Amendoeira, Class, Pinheiro, Fonseca and Barbosa2021).
Blastocystis sp. and Dientamoeba fragilis are among the most common microeukaryotes in the human gut globally, yet their clinical significance remains debatable (Shasha et al., Reference Shasha, Treygerman, Levy-dahari, Bilavsky, Hacham, Grupel, Paran, Praigrod and Zacay2024). Although there is epidemiological and clinical evidence associating Blastocystis sp. with intestinal (e.g. irritable bowel syndrome and inflammatory bowel disease) and extra-intestinal (e.g. urticaria) disorders (Tan, Reference Tan2008). Recent metagenomic population studies have linked Blastocystis sp. to relatively high microbiota diversity and stability, healthier diets and cardiometabolic benefits (Piperni et al., Reference Piperni, Nguyen, Manghi, Kim, Pasolli, Andreu-sánchez, Arrè, Bermingham, Blanco-Mínguez, Manara, Valles-Colomer, Bakker, Busonero, Davies and Fiorillo2024). Results from these surveys suggest that Blastocystis sp. might have a role in modulating both the structure and composition of the gut microbiota and promoting eubiosis (Stensvold, Reference Stensvold2025).
Giardia duodenalis, Cryptosporidium spp. and Blastocystis sp. exhibit significant genetic diversity within the small subunit ribosomal RNA (ssu rRNA) gene and other genetic markers. G. duodenalis encompasses 8 genetic assemblages (designated A–H) with marked differences in host range and specificity (Ryan et al., Reference Ryan, Feng, Fayer and Xiao2021). Recent large-scale, retrospective sequence analyses suggest that some of these assemblages may warrant recognition as independent species (Wielinga et al., Reference Wielinga, Williams, Monis and Thompson2023). At least 44 species and over 120 genotypes have been reported within the Cryptosporidium genus (Ryan et al., Reference Ryan, Feng, Fayer and Xiao2021). In Africa, 5 G. duodenalis assemblages and 13 Cryptosporidium species have been identified in humans, with assemblages A and B (within G. duodenalis) and C. hominis and C. parvum (within Cryptosporidium) causing most human cases of giardiasis and cryptosporidiosis (Squire and Ryan, Reference Squire and Ryan2017). Similarly, 44 distinct subtypes (ST) have been recognized within Blastocystis sp. (Maloney et al., Reference Maloney, Molokin, Seguí, Maravilla, Martínez-hernández, Villalobos, Tsaousis, Gentekaki, Muñoz-antolí, Klisiowicz, Oishi, Toledo, Esteban, Köster, de Lucio, Dashti, Bailo, Calero-bernal, González-barrio, Carmena and Santín2022; Santin et al., Reference Santin, Figueiredo, Molokin, George, Köster, Dashti, González-Barrio, Carmena and Maloney2024), of which ST1-ST3 are those predominantly found in African human populations (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024).
Madagascar is a very low-income country with a gross national annual income per capita of 595 USD in 2025 (International Monetary Fund, 2025) and a low public investment in WASH (Water, Sanitation and Hygiene) initiatives (UNICEF Executive Board, 2024; World Bank, 2025). These conditions, together with poor nutrition, create a favourable environment for the transmission of intestinal parasites (Global Nutrition Report, 2025). Previous epidemiological studies assessing the epidemiology of GIP in the island have reported G. duodenalis (7–79%) and Blastocystis sp. (12–77%) as the most prevalent protists of public health relevance in humans, followed by Cryptosporidium spp. (0.2–22%; Supplementary Table S1). However, genotyping studies assessing the molecular diversity of these pathogens are scarce and restricted to certain geographical areas. This information would be useful to ascertain sources of infection and transmission pathways.
This study aims at investigating the burden of GIP among infants, children, adolescents and young mothers at risk of social exclusion in Madagascar. Additionally, genotyping analyses were conducted on selected GIP pathogens to ascertain their molecular diversity.
Materials and methods
Study design and setting
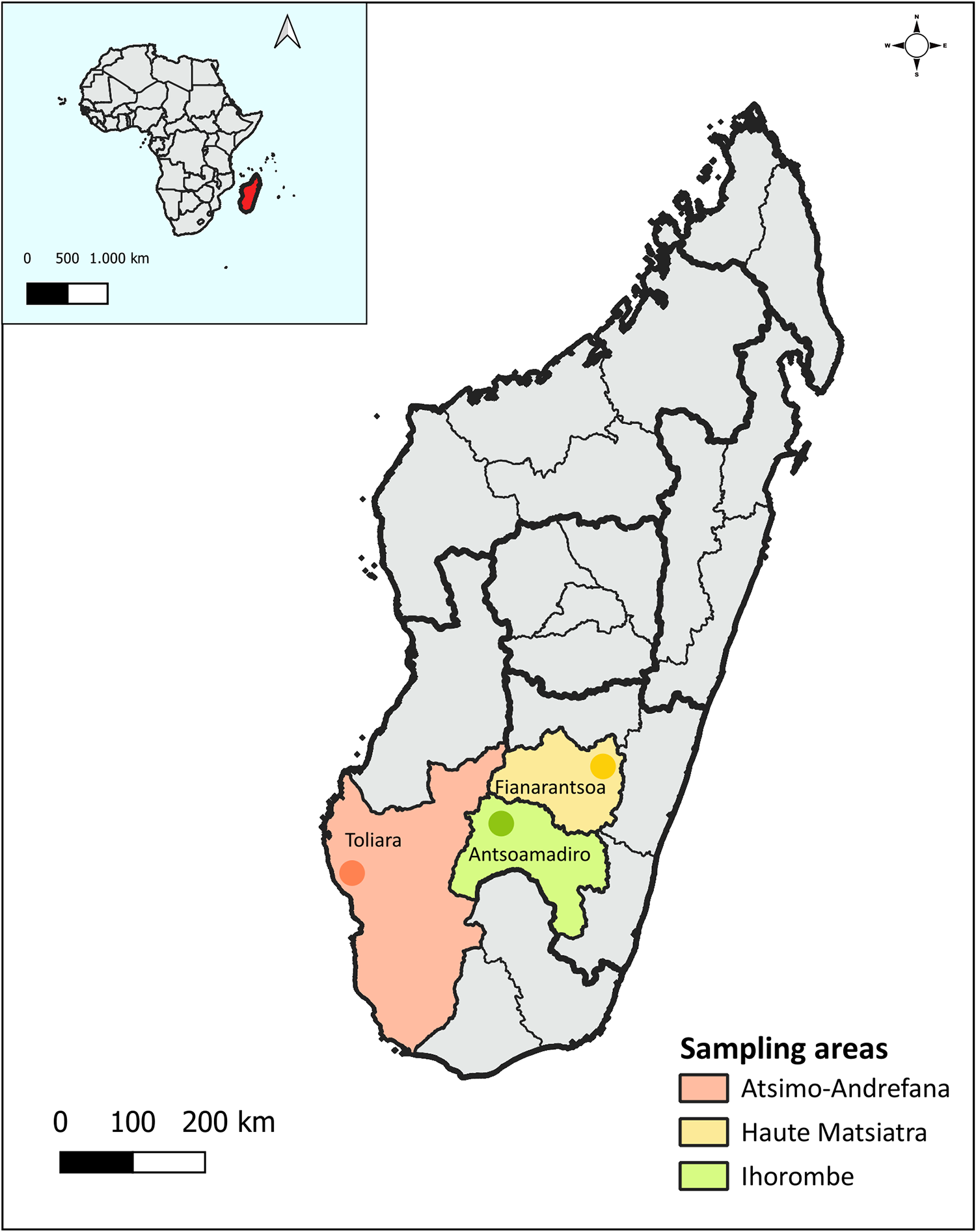
This is a prospective, cross-sectional observational study conducted between January to March and June to July 2023 among school students and young mothers with their babies from very low-resource areas in the cities of Antsoamadiro (Ihorombe Region), Fianarantsoa (Haute Matsiatra Region) and Toliara (Atsimo-Andrefana Region) in southern Madagascar (Figure 1). These localities were chosen for sampling purposes because data on the epidemiology of GIP in these areas are absent or in need of update.
Map of Madagascar showing the geographical location of the 3 cities (Antsoamadiro, Fianarantsoa and Toliara) where sampling was conducted. The map was created using QGIS v3.36.3. The base layers of the map were obtained from https://diva-gis.Org/data.Html (Madagascar) and https://open.Africa/dataset/africa-shapefiles (Africa) and are freely available for academic and non-commercial use and compatible with the CC BY 4.0 license.

Figure 1 Long description
The map of Madagascar highlights three sampling areas: Atsimo-Andrefana, Haute Matsiatra and Ihorombe. Atsimo-Andrefana is marked in orange, Haute Matsiatra in yellow and Ihorombe in green. The map includes a scale bar indicating distances of 0, 100 and 200 km. An inset map shows Madagascar's location relative to Africa, with a scale of 500 and 1,000 km. The sampling areas are labeled with the cities Toliara, Fianarantsoa and Antsoamadiro. The map outlines regional boundaries and provides a compass for orientation.
Antsoamadiro is a rural area that lacks basic services. Its local economy depends primarily on sapphire mining, and residents rely on wells for drinking water. Fianarantsoa is a more developed city with a diversified economy that includes livestock farming, agriculture and tourism. Its population enjoys better access to essential services. Toliara is a coastal city with peri-urban areas. The main sources of income there include salt production, as well as artisanal and subsistence fishing. All 3 areas are in the southern part of Madagascar, a region particularly vulnerable to prolonged droughts and flooding caused by cyclones – events that have intensified with climate change. As a result, its population is experiencing a severe food security crisis, with hunger levels nearing famine. This crisis is especially acute in parts of the Ihorombe region and even more critical in the far southern areas, such as the Atsimo-Andrefana region (World Bank, 2023).
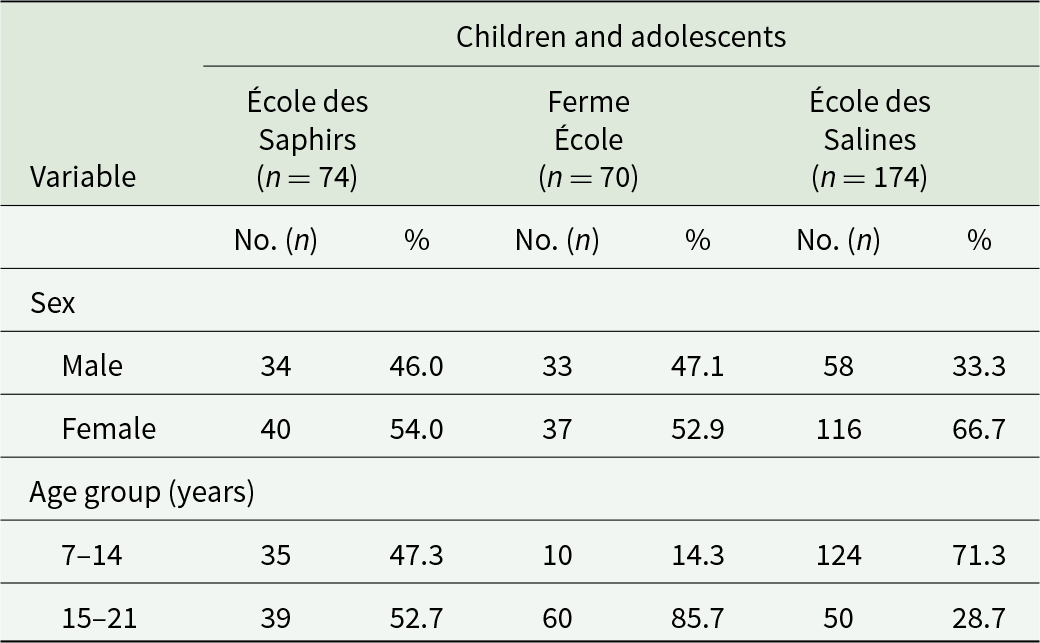
A non-probabilistic convenience sampling approach was used to select the study population, with a sample drawn proportionally to the total number of students enrolled in the 3 selected educational institutions. A total of 318 children and adolescents (7–21 years of age) were recruited: 74 from École des Saphirs in Antsoamadiro, 70 from Ferme École in Fianarantsoa and 174 from École des Salines in Toliara, the largest of the 3 institutions. Because Madagascar has high rates of illiteracy, access to education is promoted without age restrictions, which explains the broad age range of the participants (Table 1).
Distribution of participating children and adolescents, according to sex, age group and geographical origin in the present study

Table 1 Long description
The table reports counts and percentages of participating children and adolescents by sex and age group across three sites: École des Saphirs with 74 participants, Ferme École with 70, and École des Salines with 174. By sex, École des Saphirs was 46.0 percent male and 54.0 percent female, and Ferme École was 47.1 percent male and 52.9 percent female. École des Salines differed, with 33.3 percent male and 66.7 percent female. By age group, École des Saphirs was fairly balanced between ages 7–14 at 47.3 percent and ages 15–21 at 52.7 percent. Ferme École skewed older, with 14.3 percent ages 7–14 and 85.7 percent ages 15–21. École des Salines skewed younger, with 71.3 percent ages 7–14 and 28.7 percent ages 15–21. Comparisons should consider that the total number of participants differs across sites, especially the larger sample at École des Salines.
In addition, all young mothers aged 15–21 years (n = 57) and their children aged 0–5 years (n = 61) attending the Centre d’Accueil et de Soins Intégrés pour Femmes (CAIF) in Toliara were included in the study. Among the children, 43 were boys, and 18 were girls, all identified as being at risk of social exclusion.
All these 4 establishments are part of the local non-governmental organization (NGO) Bel Avenir. Children and adolescents from Antsoamadiro were similarly distributed according to the age groups considered (47.3% vs 52.7%). Most participants (85.7%) from Fianarantsoa belonged to the 15–21-year-old group. Most school students (71.3%) from Toliara belonged to the 7–14-year-old group.
Sample collection
With the support of the Health Department of the Bel Avenir NGO, visits were conducted to each institution to inform participants about the study’s objectives and procedures. In the case of minors, their parents or legal guardians were invited to attend an informational meeting held at each of the participating institutions. Each participant who volunteered to participate in the survey received a sampling kit that included a wide-mouth, high-capacity plastic container equipped with a lid and a uniquely labelled, a spatula and a magazine page. During information sessions, participants were instructed to defecate onto the magazine page to avoid soil contamination. Using the spatula, they were asked to collect approximately 5 g of faecal material from 3 different areas of the stool and place the sample into the provided container. Samples were either collected during school hours or submitted the following day at the respective centre. In cases of diarrhoea or very loose stools, participants collected the entire sample directly into the container. They were instructed to wash their hands thoroughly after completing the procedure for health and safety reasons. The samples were promptly transported to the laboratory or dispensary of each educational centre, then transferred into sterile collection tubes with 70% ethanol and stored at 4 °C for 2–4 weeks prior to shipping to Spain.
Epidemiological questionnaire
A structured epidemiological questionnaire, initially drafted in French, was used to collect information on sociodemographic (age, sex, number of household residents, education level and occupation), behavioural (personal and environmental hygiene practices, food preparation methods) and clinical (gastrointestinal symptoms) variables. The questionnaire was administered through face-to-face interviews at the time of sample collection in the participating institutions. For students under 15 years of age, the interview was conducted with the parents/legal guardians who attended the scheduled meeting at each educational centre. For parents/legal guardians who were non-French speakers or illiterate, interviews were conducted in Malagasy by a staff member of the NGO Bel Avenir, who provided oral translation of the epidemiological questionnaire.
Microscopy detection of GIPs
Collected stool samples were shipped to the Parasitology Laboratory at the Faculty of Pharmacy, University of Valencia (Spain), at room temperature for further processing. Samples were processed within 1 month after reception. Individual stool samples were concentrated and filtered using Midi Parasep Solvent Free devices (Apacor Ltd, Wokingham, United Kingdom) for 5 min at 2500 rpm. The sediment obtained was divided into 2 aliquots: 1 was intended for genomic DNA extraction and the other fixed in 10% formalin at a 1:3 ratio and intended for coproparasitological examination. Detection of parasitic forms (trophozoites and cysts) was performed by microscopic examination of both fresh smears from concentrated faecal material in saline and iodine solutions. Examination was conducted at 100× and 400× magnification (fresh smears) on a Ci-L plus microscope (Nikon, Tokyo, Japan). No Ziehl–Neelsen staining was performed for the specific detection of coccidian protists, including Cryptosporidium spp.
DNA extraction and purification
Total faecal DNA was extracted from 200 mg of each concentrated sample available using the commercial QIAamp Fast DNA Stool Mini Kit (Qiagen, Hilden, Germany), following the manufacturer’s instructions. The purified DNA was eluted in 100 μL of PCR-grade water and stored at 4 °C until further molecular analysis.
Molecular detection and characterization of selected GIPs
The identification of G. duodenalis was performed using real-time PCR (qPCR) targeting a 62-base pair (bp) fragment of the ssu RNA gene of the parasite (Verweij et al., Reference Verweij, Schinkel, Laeijendecker, van Rooyen, van Lieshout and Polderman2003). G. duodenalis-positive isolates yielding cycle threshold (CT) values ≤34 by qPCR were subsequently reassessed by a nested PCR to amplify a fragment of the ssu rRNA gene (ssu-PCR) (Hopkins et al., Reference Hopkins, Meloni, Groth, Wetherall, Reynoldson and Thompson1997; Appelbee et al., Reference Appelbee, Frederick, Heitman and Olson2003) to determine the molecular diversity of the parasite at the assemblage level. Samples that tested positive by ssu-PCR were re-amplified at the genes codifying the glutamate dehydrogenase (gdh), β-giardin (bg) and triose phosphate isomerase (tpi) proteins to determine the molecular diversity of the parasite at the sub-assemblage level. A semi-nested PCR was used to amplify a fragment of the gdh gene (Read et al., Reference Read, Monis and Thompson2004), and nested PCRs were used to amplify fragments of the bg and tpi genes, respectively (Cacciò et al., Reference Cacciò, De Giacomo and Pozio2002; Sulaiman et al., Reference Sulaiman, Fayer, Bern, Gilman, Trout, Schantz, Das, Lal and Xiao2003; Lalle et al., Reference Lalle, Pozio, Capelli, Bruschi, Crotti and Cacciò2005).
The presence of Cryptosporidium spp. was assessed using a nested-PCR protocol targeting a 587-bp fragment of the ssu rRNA gene of the parasite (Tiangtip and Jongwutiwes, Reference Tiangtip and Jongwutiwes2002). A subtyping tool based on the amplification of an 850–900-bp partial sequence of the 60-kDa glycoprotein (gp60) gene was used to ascertain intra-species genetic diversity in samples that tested positive for C. hominis or C. parvum (Feltus et al., Reference Feltus, Giddings, Schneck, Monson, Warshauer and McEvoy2006) or C. felis (Rojas-Lopez et al., Reference Rojas-Lopez, Elwin, Chalmers, Enemark, Beser and Troell2020).
Screening for Blastocystis sp. was performed using direct PCR targeting a 600-bp fragment of the ssu rRNA gene of the protist (Scicluna et al., Reference Scicluna, Tawari and Clark2006).
For the identification of B. coli, a direct PCR protocol was used to amplify a 400-bp fragment of the of the internal transcribed spacer (ITS) regions as well as the ssu rRNA gene (ITS1–5.8s-rRNA–ITS2) (Ponce-Gordo et al., Reference Ponce-Gordo, Jimenez-ruiz and Martínez-díaz2008).
General PCR and gel electrophoresis procedures
The qPCR protocol described above was carried out on a Corbett Rotor Gene 6000 real-time PCR system (Qiagen), with reaction mixes containing 2 × TaqMan Gene Expression Master Mix (Applied Biosystems, CA, United States). All the semi-nested and nested PCR procedures mentioned above were performed on a 2720 Thermal Cycler (Applied Biosystems). Reaction mixes consistently included 2.5 units of MyTAQ DNA polymerase (Bioline GmbH, Luckenwalde, Germany) and 5–10 μL of MyTAQ Reaction Buffer containing 5 mM dNTPs and 15 mM MgCl2. Laboratory-confirmed positive and negative DNA samples of human origin were routinely used as controls and included in each round of PCR.
PCR amplicons were visualized on 1.5–2% agarose gels (Conda, Madrid, Spain) stained with Pronasafe (Conda) nucleic acid staining solution. A 100 bp DNA ladder (Boehringer Mannheim GmbH, Baden-Wurttemberg, Germany) was used for the sizing of obtained amplicons.
Sequence and phylogenetic analyses
PCR products of the expected size on agarose gel were submitted for Sanger sequencing in both directions using appropriate internal primer sets. DNA sequencing was conducted by capillary electrophoresis using the BigDye Terminator chemistry (Applied Biosystems) on an ABI PRISM 3130 automated DNA sequencer. Obtained chromatograms were visually inspected for quality control and for detecting the presence of ambiguous (double peak) positions. Blastocystis sp. ST confirmation and allele calling were carried out at the Blastocystis sp. 18S typing database (https://pubmlst.org/bigsdb?db=pubmlst_blastocystis_seqdef). Sequences obtained in this study were deposited in GenBank under accession numbers PV739810–PV739816 and PV764959–PV765060 (G. duodenalis), PV739032–PV739034 and PV763025 (Cryptosporidium spp.) and PV688270–PV688305 (Blastocystis sp.).
To analyse the phylogenetic relationship among various ST families of C. felis, a maximum-likelihood tree was constructed using MEGA version 11. based on substitution rates calculated with the general time reversible model and gamma distribution with invariant sites (G + I) (Tamura et al., Reference Tamura, Stecher and Kumar2021). Bootstrapping with 1000 replicates was used to determine support for the clades.
Statistical analyses
Binary logistic regression analyses were conducted to assess the relationship between GIP and the independent variables collected in the epidemiological questionnaire. A descriptive analysis of the sample was performed using basic statistics such as percentages. This was followed by a bivariate analysis using chi-square tests for categorical variables, identifying preliminary significant associations. The magnitude of the association was expressed as odds ratios (OR) with a 95% confidence interval (CI). P-value ≤ 0.05 was considered statistically significant. All analyses were performed using SPSS Statistics version 29.0.2 for Windows (IBM Corporation, Armonk, NY, USA).
Results
A total of 436 individual stool samples from children and adolescents (n = 318; range 7‒21 years, median: 14 years) and young women (n = 57; range 16‒21 years, median: 18 years) and their babies (n = 61; range 0‒5 years, median: 1 year) were available for analyses. Fully completed epidemiological questionnaires were provided by 186 school children and adolescents (response rate: 58.5%, 186/318) and 35 young women (response rate: 61.4%, 35/57).
Detection of GIPs
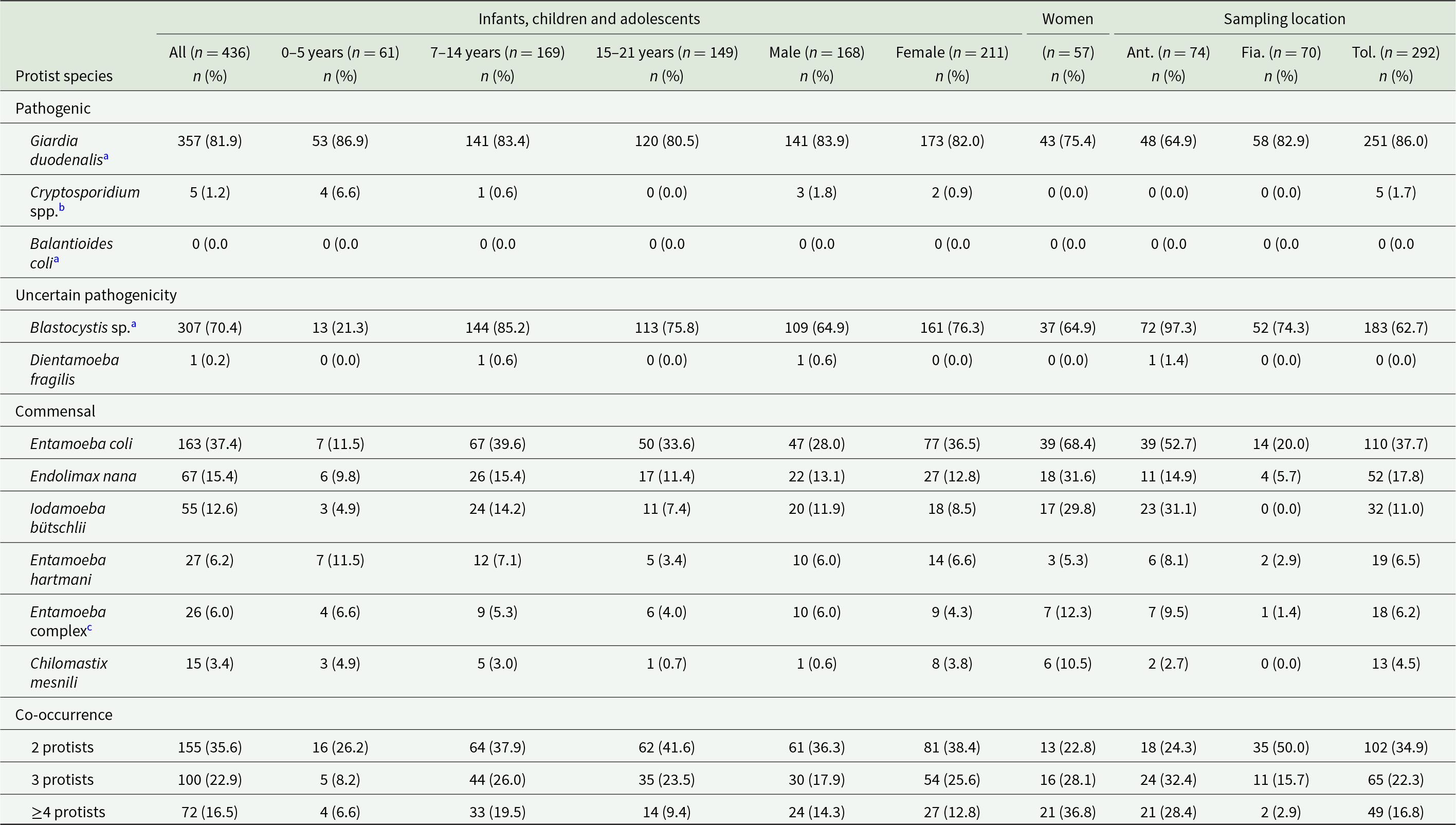
Table 2 shows composite results generated by conventional (microscopy examination) and/or molecular (PCR) methods. The overall prevalence of GIP in the study population was 96.6% (421/436; 95% CI: 94.4–98.1). At least 1 GIP species was observed in 96.0% (364/379; 95% CI: 93.6–97.8) and 100% (57/57; 95% CI: 93.7–100) of the participating children and adolescents and young women, respectively.
Frequencies of infection/colonization by intestinal protist species detected by microscopy in infants, children and adolescents (according to age group and gender) and young women in the 3 sampling locations investigated in the present study

Table 2 Long description
The table reports counts and percentages of intestinal protist infections or colonization detected by microscopy, broken down by age group, sex, women, and three sampling locations. Giardia duodenalis is the dominant finding, affecting 357 of 436 infants, children, and adolescents (81.9 percent), with similarly high levels across ages and sexes and the highest location value in Toliara (86.0 percent). Blastocystis is also common overall (70.4 percent) but shows strong variation: low in ages zero to five (21.3 percent), high in ages seven to fourteen (85.2 percent), and extremely high in Antsoamadiro (97.3 percent). Cryptosporidium is rare (1.2 percent overall) and appears only in the youngest age group and in Toliara. Several commensal protists are moderately frequent, including Entamoeba coli (37.4 percent overall) and Endolimax nana (15.4 percent), with notably higher values among women for some commensals, such as Entamoeba coli at 68.4 percent. Co-occurrence is common: 35.6 percent have two protists, 22.9 percent have three, and 16.5 percent have four or more, with the highest multiple-protist burden among women (four or more at 36.8 percent). Percentages reflect microscopy detection and should be interpreted with caution when comparing groups with different sample sizes.
a Protist species investigated by both microscopy and PCR methods.
b Protist species were investigated using only PCR methods.
c Entamoeba complex: E. histolytica/E. dispar/E. moshkovskii/E. bangladeshi.
Abbreviations: Ant., Antsoamadiro; Fia., Fianarantsoa; and Tol., Toliara.
Among pathogenic species, G. duodenalis (81.9%, 357/436; 95% CI: 77.9–85.4) was the most prevalent species found, followed by Cryptosporidium spp. (1.2, 5/436; 95% CI: 0.1–2.1). None of the stool samples analysed tested positive for B. coli. Among potentially pathogenic species, Blastocystis sp. (70.4%, 307/436; 95% CI: 65.9–74.7) was far more prevalently found than D. fragilis (0.2%, 1/436; 95% CI: 0.01–1.3). Among commensal species, Entamoeba coli (37.4%, 163/436; 95% CI: 32.8–42.1) was more prevalently detected, followed by Endolimax nana (15.4%, 67/436; 95% CI: 12.1–19.1), Iodamoeba bütschlii (12.6%, 55/436; 95% CI: 9.7–16.1), Entamoeba hartmani (6.2%, 27/436; 95% CI: 4.1–8.9), members of the Entamoeba complex (6.0%, 26/436; 95% CI: 3.9–8.6) and Chilomastix mesnili (3.4%, 15/436; 95% CI: 1.9–5.6; Table 2).
Among pathogens, G. duodenalis infections were similarly distributed among participating infants, children and adolescents regardless age group (range: 80.5–86.9%) and sex (range: 82.0–83.9%), but this rate was slightly lower in young women (75.4%). The highest G. duodenalis infection rate was identified in Toliara (86.0%) and the lowest in Antsoamadiro (64.9%). Four out of 5 (80.0%) of the Cryptosporidium-positive cases (all from Toliara) were identified in children younger than 5 years of age (Table 2).
Among potential pathogens, Blastocystis sp. colonization was lower in children under 5 years of age (21.3%), but this rate was relatively stable in older individuals (range: 75.8–85.2) and young women (64.9%). The highest Blastocystis sp. colonization rate was identified in Antsoamadiro (97.3%) and the lowest in Toliara (62.7%). Dientamoeba fragilis was detected by microscopy, in a fresh smear, in a single sample (a 12-year-old boy) from Antsoamadiro (Table 2).
Infection/colonization by a single GIP species was identified in 21.6% (94/436) of samples. Two species were identified in 35.6% (155/436), most frequent combination being G. duodenalis + Blastocystis sp. (73.6%, 114/155). Three species were found in 22.9% (100/436), while 4 or more species were present in 16.5% (72/436) of samples. Only 15 of the participants (all from the infant/school students’ group) tested negative for GIP in the coproparasitological examination (Table 2).
Molecular characterization of G. duodenalis-positive DNA samples
Giardia duodenalis-positive samples by qPCR (n = 356) generated CT values ranging from 16.7 to 44.9 (median: 33.5; standard deviation: 4.9). Of these, 38.8% (138/356) had CT values ≤34 and were selected for genotyping purposes to maximize available resources. Of them, 84.1% (116/138) were successfully amplified at the ssu rRNA marker, 39.9% (55/138) at the gdh marker and 23.9% (33/138 each) at the bg and tpi marker. Only 15.9% (22/138) of the samples failed to be amplified in at least 1 marker. Genotyping results at 1, 2, 3 and 4 loci were generated for 45.7% (53/116), 25.0% (29/116), 8.6% (10/116) and 20.7% (24/116), respectively.
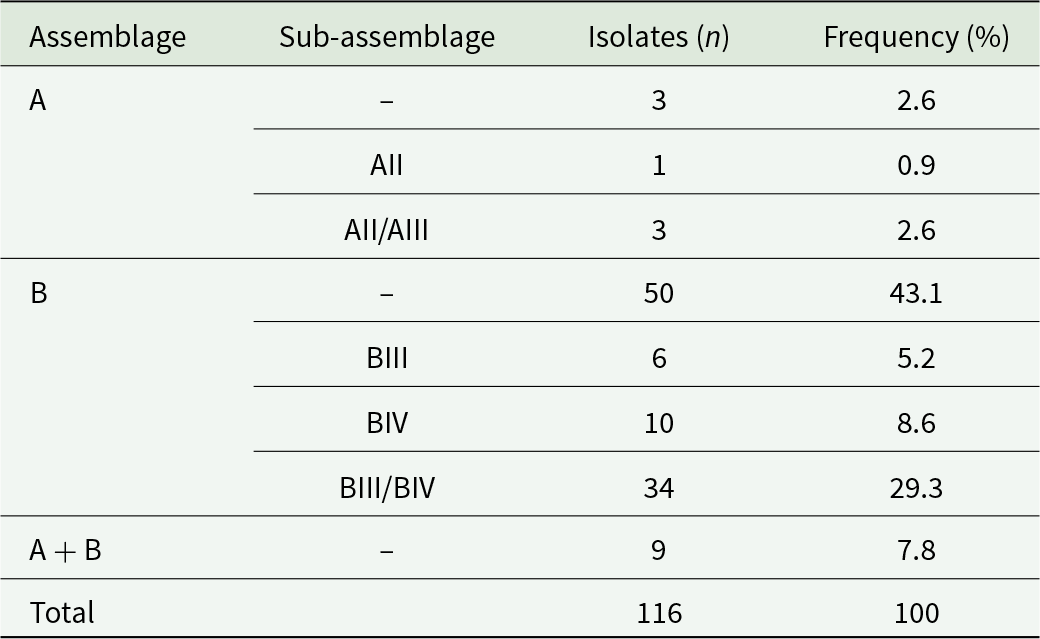
Overall, assemblage B (86.2%, 100/116) was more prevalent than assemblage A (6.0%, 7/116). Mixed A + B infections were detected in 7.8% (9/116) of cases. Detailed information of the intra-assemblage genetic diversity observed in these samples is presented in Table 3 and Supplementary Table S2.
Molecular diversity and frequency of Giardia duodenalis assemblages and sub-assemblages in the studied population

Table 3 Long description
The table reports counts and percentages of Giardia duodenalis isolates by assemblage and, where available, sub-assemblage, totaling 116 isolates. Assemblage B was most common with 50 isolates (43.1%). Within assemblage B, sub-assemblage BIII/BIV was the largest subgroup at 34 isolates (29.3%), followed by BIV at 10 (8.6%) and BIII at 6 (5.2%). Assemblage A was uncommon with 3 isolates (2.6%), and additional A sub-assemblage entries included AII at 1 (0.9%) and AII/AIII at 3 (2.6%). Mixed infections labeled A plus B accounted for 9 isolates (7.8%). Percentages are presented as shares of the full set of 116 isolates, and some rows represent subtyping detail rather than separate totals for an assemblage.
Molecular characterization of Blastocystis sp.-positive DNA samples
Overall, 77.1% (336/436) of the faecal DNA samples analysed produced amplicons of the expected size on agarose gels. Of these, 63.7% (214/336) were confirmed by Sanger sequencing. In the absence of Sanger sequencing confirmation, the 122 samples with PCR products compatible with Blastocystis sp. were conservatively considered negative to this protist by PCR.
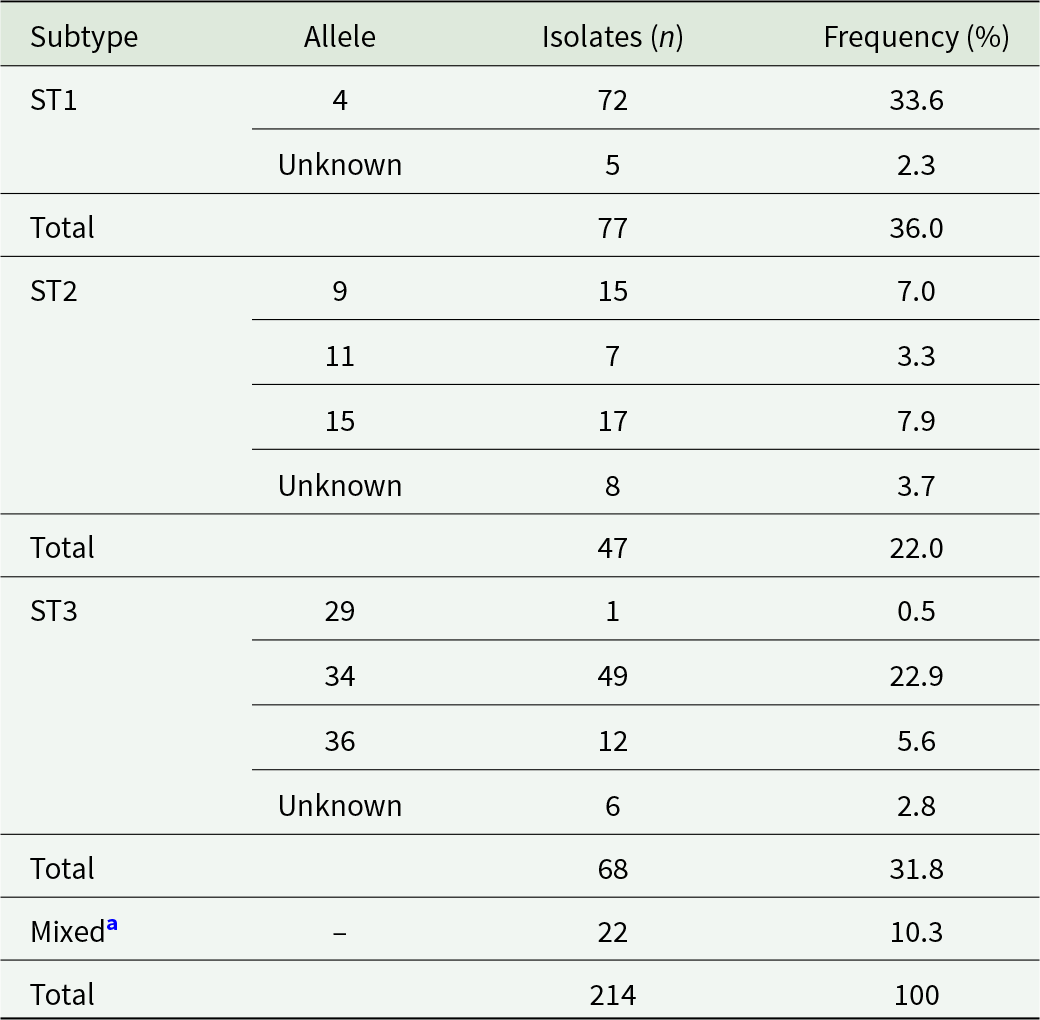
Three distinct STs of Blastocystis sp. were observed at the ssu rRNA locus, with ST1 being the most prevalent (36.0%, 77/214), followed by ST3 (31.8%, 68/214), and ST2 (22.0%, 47/214; Table 4). The remaining Blastocystis sp.-positive samples (10.3%, 22/214) corresponded to mixed infections as determined by overlapping sequence traces at chromatogram inspection. Detailed information of the intra-allelic genetic diversity observed in these samples is presented in Table 4 and Supplementary Table S3.
Molecular diversity and frequency of Blastocystis subtypes and alleles in the studied population

Table 4 Long description
The table summarizes Blastocystis subtypes and their alleles, reporting isolate counts and percent frequency in a total of 214 isolates. ST1 is the most frequent subtype with 77 isolates, accounting for 36.0 percent; most ST1 isolates are allele 4 with 72 isolates, while 5 are listed as unknown allele. ST3 is next with 68 isolates at 31.8 percent, dominated by allele 34 with 49 isolates; allele 36 has 12 isolates, allele 29 has 1 isolate, and 6 are unknown. ST2 contributes 47 isolates at 22.0 percent, led by allele 15 with 17 isolates, followed by allele 9 with 15, allele 11 with 7, and 8 unknown. Mixed infections account for 22 isolates at 10.3 percent and are not assigned a specific allele. Some entries are marked unknown allele, reflecting sequences that could not be confidently assigned due to overlapping trace patterns.
a Samples with overlapping sequence traces at chromatogram inspection.
Molecular characterization of Cryptosporidium spp.-positive DNA samples
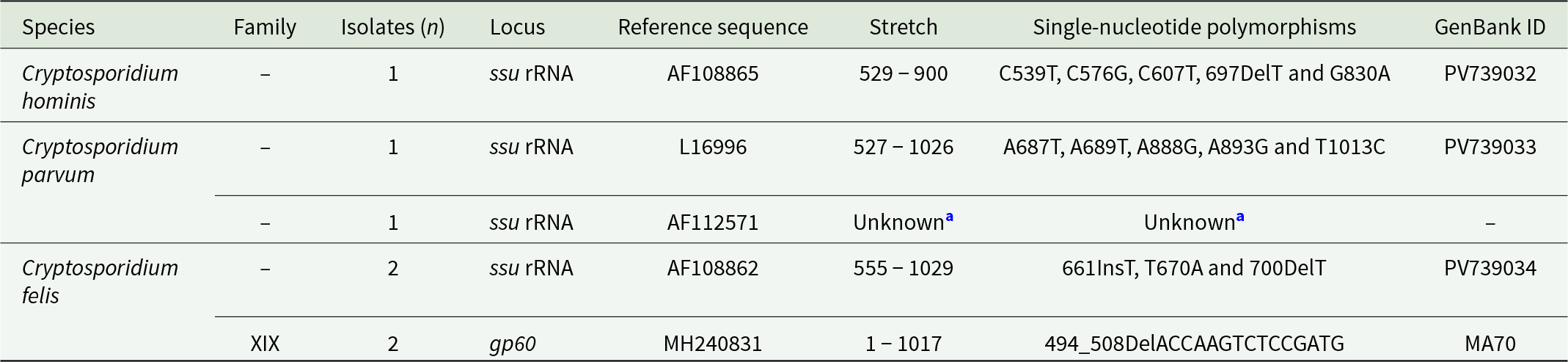
Sequence analysis of the 5 samples amplified at the ssu rRNA locus allowed the identification of 3 Cryptosporidium species, including C. hominis (20.0%, 1/5), C. parvum (40.0%, 2/5) and C. felis (40.0%, 2/5). Detailed information of the intra-species genetic diversity observed in these samples is presented in Table 5. The 2 sequences identified as C. felis corresponded to 2 children under 5 years of age sampled at the CAIF (Toliara region), with no family relationship between them.
Frequency and molecular diversity of Cryptosporidium spp. Identified at the ssu RNA and gp60 loci in the population under study, Madagascar 2008–2024. GenBank accession numbers are provided

Table 5 Long description
The table lists Cryptosporidium species detected in the study population and summarizes isolate counts, the gene locus sequenced, the reference sequence used, the sequenced stretch, observed single nucleotide changes or small insertions and deletions, and GenBank identifiers. Cryptosporidium hominis was represented by one isolate at the ssu rRNA locus, with multiple listed nucleotide changes and GenBank ID PV739032. Cryptosporidium parvum was represented by one isolate at ssu rRNA with several nucleotide changes and GenBank ID PV739033, plus an additional ssu rRNA entry tied to reference AF112571 where the stretch and polymorphisms could not be determined and no GenBank ID is provided. Cryptosporidium felis was the most frequent, with two isolates at ssu rRNA showing an insertion, a substitution, and a deletion, deposited as PV739034. The same species also had two isolates typed at the gp60 locus in family XIX, showing a deletion event and a GenBank ID recorded as MA70. Comparisons across rows indicate that only C. felis has data at both loci, while C. hominis and C. parvum are only reported at ssu rRNA. One entry includes unknown sequence details, so diversity at that locus may be underestimated.
a Sequence of insufficient quality to determine stretch and single-nucleotide polymorphisms.
gp60: 60 kDa glycoprotein; ssu rRNA: small subunit ribosomal RNA.
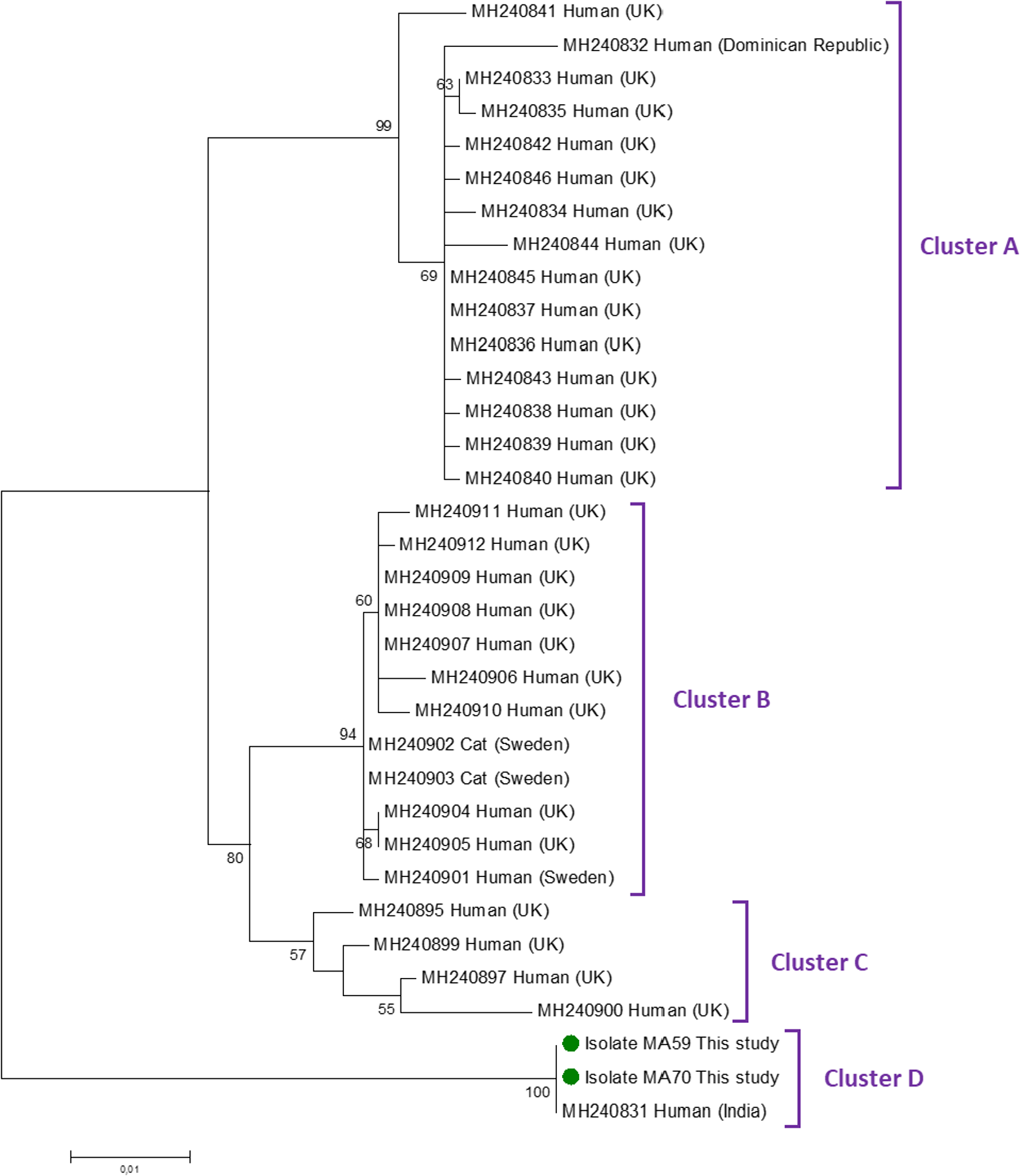
Only the 2 C. felis samples were successfully amplified using the gp60 marker. Sequence analysis confirmed that both belonged to family XIX. Figure 2 shows the maximum-likelihood tree generated with representative C. felis sequences of the 4 clusters (A–D) proposed by Rojas-Lopez et al. (Reference Rojas-Lopez, Elwin, Chalmers, Enemark, Beser and Troell2020). Our 2 gp60 family XIX isolates fell within Cluster D. Its closest match (98.5% identity) was a C. felis isolate identified in a subject from India.
Phylogenetic relationship among Cryptosporidium felis subtype family XIX revealed by a maximum likelihood analysis of the partial gp60 gene. Substitution rates were calculated by using the general time-reversible model. Numbers on branches are bootstrapping values over 50% using 1000 replicates. The filled green circles indicate the nucleotide sequences generated in the present study.

Figure 2 Long description
The diagram is a maximum-likelihood tree illustrating the phylogenetic relationships among Cryptosporidium felis subtype family XIX. The tree is oriented vertically, with branches extending from top to bottom. The tree is divided into four clusters labeled A, B, C and D. Cluster A includes sequences from humans in the UK and the Dominican Republic, with bootstrapping values of 90 and 69. Cluster B contains sequences from humans in the UK and Sweden, as well as cats in Sweden, with bootstrapping values of 60 and 59. Cluster C includes sequences from humans in the UK, with a bootstrapping value of 57. Cluster D features two isolates from the present study, labeled as Isolate MAS9 and Isolate MA70, marked with filled green circles and a sequence from a human in India, with a bootstrapping value of 100. The branches are labeled with specific sequence identifiers, such as MH240831 and MH240832, indicating the source and location of each sequence. The scale bar at the bottom left represents genetic distance.
Risk factor analyses
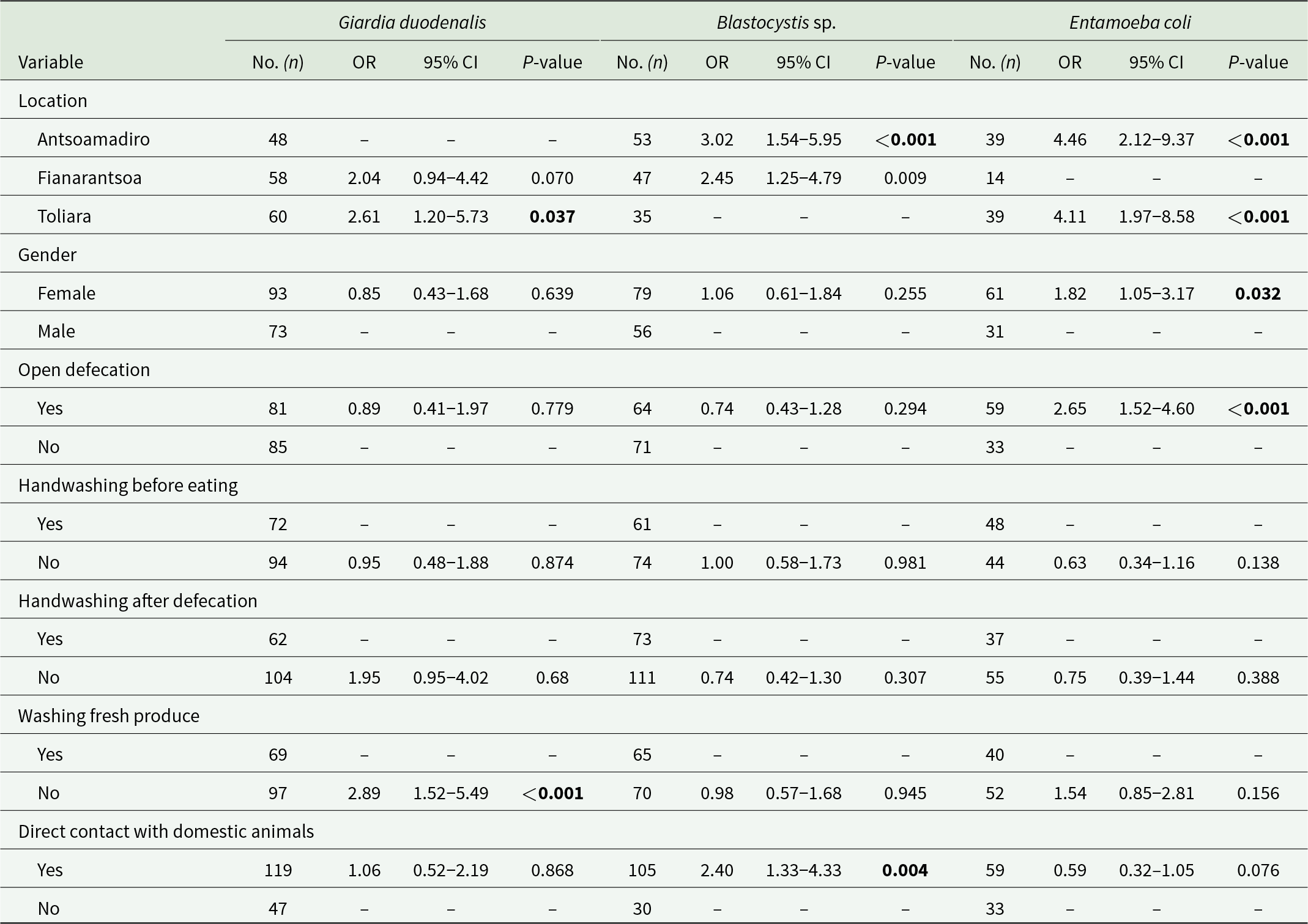
Binary logistic regression analyses were performed using survey data to assess the risk of infection/colonization by any GIPs based on sociodemographic and behavioural variables. An evaluation of the overall prevalence of GIP among the 221 surveyed participants revealed significant associations with the assessed risk factors only for those species that were well represented in the surveyed population. These included a well-established pathogen (G. duodenalis), a protist of uncertain pathogenicity (Blastocystis sp.) and a commensal (E. coli) (Table 6). Individuals residing in Toliara had a 2.61-fold higher likelihood of infection with G. duodenalis (95% CI: 1.20–5.73; P = 0.037). Inhabitants of Antsoamadiro exhibited a 3.02-fold increased risk of Blastocystis sp. carriage (95% CI: 1.54–5.95; P < 0.001). Furthermore, individuals living in Toliara (95% CI: 1.97–8.58; P < 0.001) and Antsoamadiro (95% CI: 2.12–9.37; P < 0.001) were more likely to be colonized by E. coli compared to those residing in Fianarantsoa. Participants who did not wash fruits and vegetables prior to consumption had a 2.89-fold higher risk of infection with G. duodenalis (95% CI: 1.52–5.94; P < 0.001) compared to those who did. Individuals with close contact with domestic animals were more likely to carry Blastocystis sp. (95% CI: 1.33–4.33; P = 0.004), whereas being female (95% CI: 1.05–3.17; P = 0.032) and open defecation (95% CI: 1.52–4.60; P < 0.001) were factors associated with a higher odds of colonization by E. coli.
Binary logistic regression analyses for variables associated with the most common intestinal protists identified in the present study. Odds ratios (OR) and 95% confidence intervals (95% CI) are indicated. Statistically significant values are in bold

Table 6 Long description
The table reports binary logistic regression results for three intestinal protists, listing sample size, odds ratios with 95 percent confidence intervals, and p-values for each exposure category. For Blastocystis, location is a strong factor: Antsoamadiro has about three times higher odds and Fianarantsoa about two and a half times higher odds than the reference location, both statistically significant. For Entamoeba coli, Antsoamadiro and Toliara each show about four times higher odds than the reference location, both statistically significant, while Fianarantsoa has no estimate reported. For Giardia duodenalis, Toliara shows higher odds than the reference location and not washing fresh produce is associated with nearly three times higher odds; both are statistically significant, while Fianarantsoa is not statistically significant. Female gender is associated with higher odds of Entamoeba coli, and open defecation is also associated with higher odds of Entamoeba coli; both are statistically significant. Direct contact with domestic animals is associated with higher odds of Blastocystis and is statistically significant, while its association with Entamoeba coli is not statistically significant. Several hygiene variables, including handwashing before eating and after defecation, show no statistically significant associations for most outcomes, and some cells are blank or marked with dashes indicating reference categories or unavailable estimates.
Symptoms and signs associated to pathogenic GIP
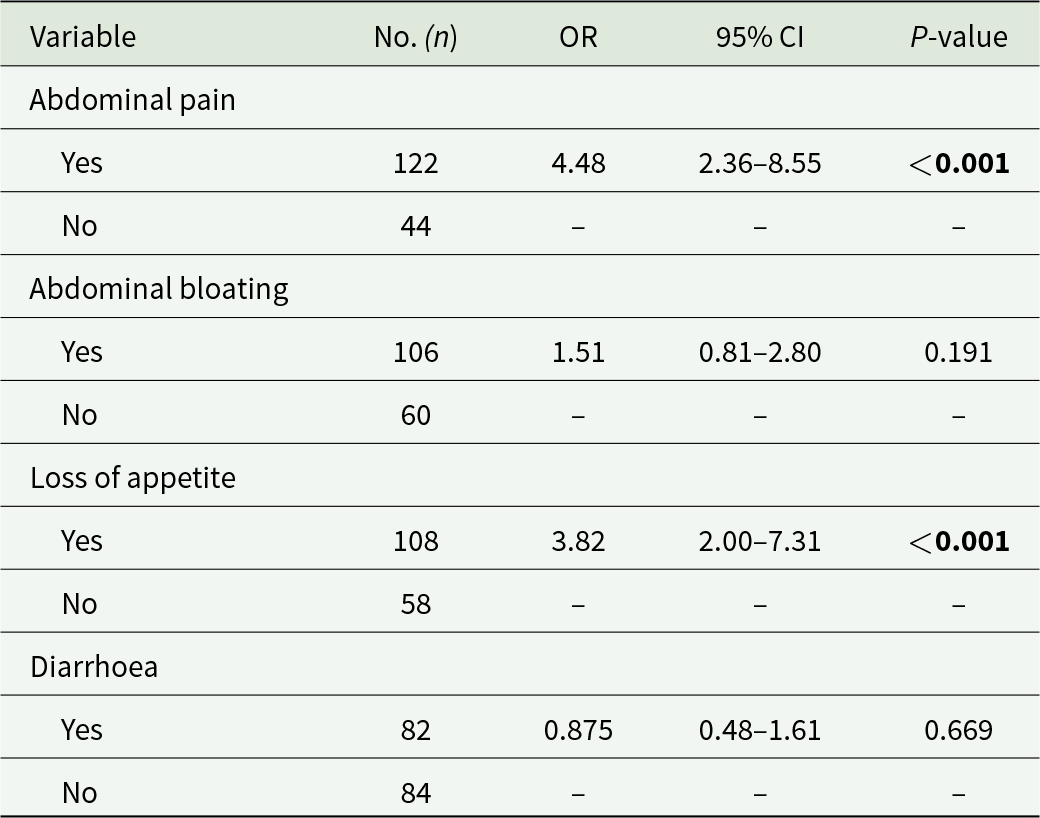
The most common symptoms and clinical signs (abdominal pain, abdominal distension, loss of appetite and diarrhoea) were analysed in relation to the occurrence of pathogenic GIP (including G. duodenalis and Cryptosporidium spp.) infections in the surveyed population. Individuals infected with G. duodenalis were significantly more likely to experience abdominal pain (OR: 4.48; 95% CI: 2.36–8.55; P < 0.001) and loss of appetite (OR: 3.82; 95% CI: 2.00–7.31; P < 0.001; Table 7).
Binary logistic regression analyses for the clinical manifestations associated with Giardia duodenalis infections (n = 166) in the present study. Odds ratios (OR) and 95% confidence intervals (95% CI) are indicated. Statistically significant values are bolded

Table 7 Long description
The table reports binary logistic regression results linking four clinical symptoms to Giardia duodenalis infection in 166 participants, giving counts, odds ratios, confidence intervals, and p-values. Abdominal pain was reported by 122 people and showed higher odds of infection (odds ratio 4.48; confidence interval 2.36 to 8.55; p-value less than 0.001) compared with those without pain (44). Loss of appetite was reported by 108 people and was also associated with higher odds (odds ratio 3.82; confidence interval 2.00 to 7.31; p-value less than 0.001) compared with those without appetite loss (58). Abdominal bloating (106 yes, 60 no) had a smaller, non-significant association (odds ratio 1.51; confidence interval 0.81 to 2.80; p-value 0.191). Diarrhoea (82 yes, 84 no) was not associated with higher odds (odds ratio 0.875; confidence interval 0.48 to 1.61; p-value 0.669). Rows labeled “No” serve as the reference group and therefore do not list odds ratios, confidence intervals, or p-values.
Discussion
GIP infections are closely associated with poverty, inadequate sanitation and limited access to healthcare (International Monetary Fund, 2025). This study focuses on populations at higher risk of infection and has several key strengths including (1) the analysis of a large panel of stool samples (n = 436), (2) the combined use of conventional (microscopy) and molecular (PCR) methods for the detection of GIP, (3) the genotyping of a significant proportion of samples that tested positive to GIP species known to be pathogenic or potentially pathogenic and (4) the analysis of risk factors potentially associated with higher odds of GIP infections.
This study shows that nearly all participants (96.6%) were infected with at least 1 species of GIP. These findings reflect an epidemiological context in which infections, reinfections and colonizations are frequent due to continuous exposure to sources of faecal contamination. Pathogenic G. duodenalis was the most prevalent GIP found either in infants, children and adolescents (81–87%) and young mothers (75%). These figures are among the highest reported in Madagascar. In previous studies conducted in paediatric populations in the country, G. duodenalis infection rates ranged from 1 to 13% by microscopic examination (Randremanana et al., Reference Randremanana, Randrianirina, Gousseff, Dubois, Razafindratsimandresy, Hariniana, Garin, Randriamanantena, Rakotonirina, Ramparany, Ramarokoto, Rakotomanana, Ratsirorahina, Rajantonirina, Talarmin and Richard2012; Remonja et al., Reference Remonja, Rakotoarison, Rakotonirainy, Mangahasimbola, Randrianarisoa, Jambou, Vigan-Womas, Piola and Randremanana2017; Razafiarimanga et al., Reference Razafiarimanga, Yao, Rajerison, Randriamampianina, Rahelinirina, Rakotoarison, Alexandra, Elisoa, Pascal and Ronan2022) and from 26 to 80% according to PCR (Frickmann et al., Reference Frickmann, Schwarz, Rakotozandrindrainy, May and Hagen2015; Habib et al., Reference Habib, Andrianonimiadana, Rakotondrainipiana, Andriantsalama, Randriamparany, Randremanana, Rakotoarison, Vigan-womas, Rafalimanantsoa, Vonaesch, Sansonetti, Collard and Afribiota2021; Richert et al., Reference Richert, Kołodziej, Zarudzka, Kasprowicz, Świetlik and Korzeniewski2024). Surveys conducted at the clinical, community or volunteer levels have also revealed the presence of G. duodenalis cysts in 8–10% of the samples analysed either by microscopy or direct fluorescence assay (Greigert et al., Reference Greigert, Abou-Bacar, Brunet, Nourrisson, Pfaff, Benarbia, Randrianarivelojosia, Razafindrakoto, Solotiana, Morel, Candolfi and Poirier2018; Spencer and Irwin, Reference Spencer and Irwin2020; Mondot et al., Reference Mondot, Poirier, Abou-Bacar, Greigert, Brunet, Nourrisson, Randrianarivelojosia, Razafindrakoto, Morel, Rakotomalala, Leclere, Le Roux, Monot, Lepage and Candolfi2021). Comparatively lower G. duodenalis infection rates ranging from 8 to 52% have been documented in Madagascar’s closest mainland neighbour countries including Malawi (Bitilinyu-Bangoh et al., Reference Bitilinyu-Bangoh, Riesebosch, Rebel, Chiwaya, Verschoor, Voskuijl and Schallig2024), Mozambique (Bauhofer et al., Reference Bauhofer, Cossa-moiane, Marques, Guimarães, Munlela, Anapakala, Chiláule, Cassocera, Langa, Chissaque, Sambo, Manhique-coutinho, Bero, Kellogg, Gonçalves and de Deus2021; Muadica et al., Reference Muadica, Balasegaram, Beebeejaun, Köster, Bailo, Hernández-de-Mingo, Dashti, Dacal, Saugar, Fuentes and Carmena2021a), Tanzania (Franz et al., Reference Franz, Fuss, Mazigo, Ruganuza and Müller2023), Zambia (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024) or Zimbabwe (Mason and Patterson, Reference Mason and Patterson1987).
As far as we know, this is the first report assessing the genetic diversity of G. duodenalis human infections in Madagascar. Assemblage B was far more prevalent than assemblage A. These results were consistent with those reported in similar molecular-based studies conducted in other sub-Saharan countries, including Angola (64% vs 36%) (Dacal et al., Reference Dacal, Saugar, de Lucio, Hernández-de-Mingo, Robinson, Köster, Aznar-ruiz-de-alegría, Espasa, Ninda, Gandasegui, Sulleiro, Moreno, Salvador, Molina, Rodríguez and Carmena2018), Mozambique (88–90% vs 7–10%) (Messa et al., Reference Messa, Köster, Garrine, Gilchrist, Bartelt, Nhampossa, Massora, Kotloff, Levine, Alonso, Carmena and Mandomando2021; Muadica et al., Reference Muadica, Köster, Dashti, Bailo, Hernández-de-Mingo, Balasegaram and Carmena2021b), Tanzania (85% vs 6%) (Forsell et al., Reference Forsell, Granlund, Samuelsson, Koskiniemi, Edebro and Evengård2016) and Zambia (71% vs 13%) (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024). Differences in assemblage distribution might be the consequence of distinct source of infection or transmission pathways. At the local/regional level, this could be the case of our study, where a significant proportion of the assemblage A cases clustered together in Toliara. Depending on the marker (gdh or tpi) considered, ambiguous BIII/BIV sequences accounted for 54–60% of the assemblage B isolates, indicative of insufficient discriminative power. This is due to the presence of high allelic sequence heterozygosity in the genomes of these parasites (Woschke et al., Reference Woschke, Faber, Stark, Holtfreter, Mockenhaupt, Richter, Regnath, Sobottka, Reiter-Owona, Diefenbach, Gosten-Heirich, Friesen, Ignatius, Aebischer and klotz2021). Indeed, some authors have indicated that there is no phylogenetic support for the BIII and BIV groups, so the use of these ST names for assemblage B should be disregarded (Brynildsrud et al., Reference Brynildsrud, Tysnes, Robertson and Debenham2018).
The Cryptosporidium spp. infection rates detected here were consistent with previous reports in asymptomatic children from Madagascar, where prevalence rates ranging from 0.2% to 3% have been observed using microscopy and/or qPCR (Frickmann et al., Reference Frickmann, Schwarz, Rakotozandrindrainy, May and Hagen2015; Remonja et al., Reference Remonja, Rakotoarison, Rakotonirainy, Mangahasimbola, Randrianarisoa, Jambou, Vigan-Womas, Piola and Randremanana2017; Richert et al., Reference Richert, Kołodziej, Zarudzka, Kasprowicz, Świetlik and Korzeniewski2024). Higher prevalence rates, between 3% and 10%, have been reported in clinical and community-based studies using microscopy, qPCR and immunofluorescence assays (Greigert et al., Reference Greigert, Abou-Bacar, Brunet, Nourrisson, Pfaff, Benarbia, Randrianarivelojosia, Razafindrakoto, Solotiana, Morel, Candolfi and Poirier2018; Spencer and Irwin, Reference Spencer and Irwin2020). The prevalence observed in this study is lower than that reported in several sub-Saharan African countries, including Mozambique (2%) (Muadica et al., Reference Muadica, Balasegaram, Beebeejaun, Köster, Bailo, Hernández-de-Mingo, Dashti, Dacal, Saugar, Fuentes and Carmena2021a), Kenya (10%) (Essendi et al., Reference Essendi, Muleke, Miheso and Otachi2022) and Zambia (4%) (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024).
The presence of C. hominis on the island has previously been documented in asymptomatic children, with a reported prevalence of 4% (Krumkamp et al., Reference Krumkamp, Conraths, Cacciò, Schares, Hogan, Winter, Jaeger, Melhem, Rakotozandrindrainy, May, Rakotozandrindrainy and Eibach2022), and in symptomatic children, where rates have ranged from 5% (Areeshi et al., Reference Areeshi, Dove, Papaventsis, Gatei, Combe, Grosjean, Leatherbarrow and Hart2008) to 22% (Manouana et al., Reference Manouana, Lorenz, Mbong, Nguema, Maiga-ascofaré, Akenten, Amuasi, Rakotozandrindrainy, Rakotozandrindrainy, Mbwana, Lusingu, Byrne, Melhem, Zinsou, Adegbite, Hogan, Winter, May, Kremsner, Borrmann, Eibach and Adegnika2020). A mixed-population study involving both symptomatic and asymptomatic children found a C. hominis prevalence of 13% (Krumkamp et al., Reference Krumkamp, Aldrich, Maiga-ascofare, Mbwana, Rakotozandrindrainy, Borrmann, Caccio, Rakotozandrindrainy, Adegnika, Lusingu, Amuasi, May, Eibach and CRYPTO2021). Cryptosporidium parvum has also been previously reported in asymptomatic Malagasy children, with a prevalence of 19% (Habib et al., Reference Habib, Andrianonimiadana, Rakotondrainipiana, Andriantsalama, Randriamparany, Randremanana, Rakotoarison, Vigan-womas, Rafalimanantsoa, Vonaesch, Sansonetti, Collard and Afribiota2021). In other sub-Saharan countries, such as Kenya (Essendi et al., Reference Essendi, Muleke, Miheso and Otachi2022), Malawi (Morse et al., Reference Morse, Nichols, Grimason, Campbell, Tembo and Smith2007), Mozambique (Muadica et al., Reference Muadica, Balasegaram, Beebeejaun, Köster, Bailo, Hernández-de-Mingo, Dashti, Dacal, Saugar, Fuentes and Carmena2021a) and Zambia (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024), both C. hominis and C. parvum are the predominant species in human infections, with prevalence ranges of 15–48% and 18–82%, respectively. Unfortunately, none of the C. parvum isolates identified in the present study could be genotyped at the gp60 marker. This information would have been crucial to determine the potential contribution of zoonotic sources, as human cryptosporidiosis cases caused by C. parvum (most of them belonging to the ST family IIc) are considered to be primarily of anthroponotic origin in Africa (Nader et al., Reference Nader, Mathers, Ward, Pachebat, Swain, Robinson, Chalmers, Hunter, van Oosterhout and Tyler2019; Robertson et al., Reference Robertson, Johansen, Kifleyohannes, Efunshile and Terefe2020).
Although C. felis is specifically adapted to infect felines, both domestic and wild, an increasing number of cases have been reported in humans, including both immunocompetent and immunocompromised individuals. In sub-Saharan Africa, C. felis has been detected in Mozambique (0.4%) (Muadica et al., Reference Muadica, Balasegaram, Beebeejaun, Köster, Bailo, Hernández-de-Mingo, Dashti, Dacal, Saugar, Fuentes and Carmena2021a) and Zambia (1%) (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024). It has also been identified in non-human primates, calves, horses and foxes (Jiang et al., Reference Jiang, Roellig, Lebbad, Beser, Troell, Guo, Li, Xiao and Feng2020; Rojas-Lopez et al., Reference Rojas-Lopez, Elwin, Chalmers, Enemark, Beser and Troell2020). The detection of C. felis in Madagascar suggests that at least some cryptosporidiosis cases in the study population may result from zoonotic transmission, either through direct contact with infected animals or indirectly via ingestion of contaminated food or water.
Interestingly, B. coli was not detected in the surveyed population. This finding was somewhat unexpected, as this ciliate is typically highly prevalent in domestic pigs and is known to cause human infections, particularly among individuals living in close proximity to pigs. Such infections are especially common in low-resource settings with limited or no access to adequate sanitation infrastructure (da Silva et al., Reference da Silva, Dib, Amendoeira, Class, Pinheiro, Fonseca and Barbosa2021), as is the case in the areas investigated in this study. Indeed, human infections with B. coli have been reported at prevalence rates ranging from 0.1% to 9% in various African countries, including Ethiopia (Feleke et al., Reference Feleke, Beyene, Feleke, Jember and Abera2019), Libya (Al Kilani et al., Reference Al Kilani, Dahesh and El Taweel2008) and Nigeria (Udeh et al., Reference Udeh, Goselle, D-popova, Abelau, Popov, Jean and David2008). Currently, we do not have a clear explanation for the absence of B. coli in our data. However, assessing its true prevalence in local pig populations (the primary reservoir of this pathogen) would provide valuable information.
Blastocystis sp. was the second most prevalent GIP identified in the studied population. Colonization rates among infants/school students varied markedly, ranging from 21% in those under 5 years of age to 85% in older individuals. Among young mothers, the colonization rate was 65%. Previous microscopy-based studies in Malagasy children reported colonization rates of 12% (Habib et al., Reference Habib, Andrianonimiadana, Rakotondrainipiana, Andriantsalama, Randriamparany, Randremanana, Rakotoarison, Vigan-womas, Rafalimanantsoa, Vonaesch, Sansonetti, Collard and Afribiota2021) and 41% (Richert et al., Reference Richert, Kołodziej, Zarudzka, Kasprowicz, Świetlik and Korzeniewski2024). Studies conducted in clinical settings or among volunteer cohorts have reported higher prevalence rates, ranging from 70% (Greigert et al., Reference Greigert, Abou-Bacar, Brunet, Nourrisson, Pfaff, Benarbia, Randrianarivelojosia, Razafindrakoto, Solotiana, Morel, Candolfi and Poirier2018) to 78% (Mondot et al., Reference Mondot, Poirier, Abou-Bacar, Greigert, Brunet, Nourrisson, Randrianarivelojosia, Razafindrakoto, Morel, Rakotomalala, Leclere, Le Roux, Monot, Lepage and Candolfi2021), using microscopy and qPCR. A DNA metabarcoding study reported a prevalence of 74% (Barrett et al., Reference Barrett, Titcomb, Janko, Pender, Kauffman, Solis, Randriamoria, Young, Mucha, Moody, Kramer, Soarimalala and Nunn2024). In contrast, studies from other sub-Saharan African countries such as Mozambique (Muadica et al., Reference Muadica, Balasegaram, Beebeejaun, Köster, Bailo, Hernández-de-Mingo, Dashti, Dacal, Saugar, Fuentes and Carmena2021a), Tanzania(Forsell et al., Reference Forsell, Granlund, Samuelsson, Koskiniemi, Edebro and Evengård2016) and Zambia (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024) have reported lower prevalence rates, ranging from 14% to 61%.
The diversity and frequency of Blastocystis STs identified in our study population were consistent with previous studies conducted in Madagascar, which reported a prevalence of ST1 ranging from 35 to 47%, ST2 from 18% to 21%, ST3 from 25% to 39% and mixed infections below 8% (Greigert et al., Reference Greigert, Abou-Bacar, Brunet, Nourrisson, Pfaff, Benarbia, Randrianarivelojosia, Razafindrakoto, Solotiana, Morel, Candolfi and Poirier2018; Barrett et al., Reference Barrett, Titcomb, Janko, Pender, Kauffman, Solis, Randriamoria, Young, Mucha, Moody, Kramer, Soarimalala and Nunn2024). Similar ST distributions have been reported in Mozambique (Muadica et al., Reference Muadica, Köster, Dashti, Bailo, Hernández-de-Mingo, Balasegaram and Carmena2021b), Tanzania (Forsell et al., Reference Forsell, Granlund, Samuelsson, Koskiniemi, Edebro and Evengård2016) and Zambia (Mutengo et al., Reference Mutengo, Kaduková, Mulunda, Chabala, Dashti, Hayashida, Chinyanta, Chisanga, Castro, Sánchez, Mwansa, Köster, González-Barrio, Maloney, Santín, Sotillo and Carmena2024), with ST1 prevalence ranging from 23% to 35%, ST2 from 23% to 44%, and ST3 from 21% to 46%. In addition to the commonly reported subtypes (ST1–ST3), ST4 has also been detected, though at low prevalence rates (<9%), in Mozambique (Muadica et al., Reference Muadica, Köster, Dashti, Bailo, Hernández-de-Mingo, Balasegaram and Carmena2021b) and Senegal (El Safadi et al., Reference El Safadi, Gaayeb, Meloni, Cian, Poirier, Wawrzyniak, Dabboussi, Delhaes, Hamze, Hamze, Riveau and Viscogliosi2014). Notably, ST4 is far more prevalent in European populations and is rarely reported, or even absent, in sub-Saharan Africa (Forsell et al., Reference Forsell, Granlund, Samuelsson, Koskiniemi, Edebro and Evengård2016), which may explain its absence in our study. The exclusive detection of subtypes ST1–ST3, along with the absence of animal-adapted STs (such as ST5, ST6 and ST7), suggests that a substantial, albeit still undetermined, proportion of the Blastocystis sp. detected in our study may have an anthroponotic origin.
In this survey, light microscopy and PCR were used for the detection of G. duodenalis and Blastocystis sp. In both cases, PCR demonstrated significantly higher sensitivity, highlighting the superior diagnostic performance compared to conventional microscopy. They are especially effective for identifying subclinical or low-intensity infections, as is often the case with G. duodenalis in endemic areas (Tijani et al., Reference Tijani, Köster, Guadano-procesi, George, Abodunrin, Adeola, Dashti, Bailo, González-Barrio and Carmena2023).
Regarding risk factors, G. duodenalis infection was more frequent in Toliara, whereas Blastocystis sp. predominated in Antsoamadiro. Both locations share similar socioeconomic conditions, with high poverty levels and limited access to WASH services. Participants who reported not washing fresh fruits and vegetables before consumption had a higher risk for G. duodenalis infection, consistent with findings from studies conducted in Cuba (Jerez Puebla et al., Reference Jerez Puebla, Núñez, Martínez Silva, Rojas Rivero, Martínez González, Méndez Sutil, Ayllón Valdés, Atencio Millán and Müller2015) and Ethiopia (Hajare et al., Reference Hajare, Betcha, Sharma, Bhosale, Upadhye, Kuddus and Eriso2022). Blastocystis sp. close contact with animals showed a significant association, similar to previous observations in Colombian children (Hernández et al., Reference Hernández, Maloney, Molokin, George, Morales, Chaparro-olaya and Santin2023); however, these findings should be interpreted with caution, as other surveys have not found evidence of zoonotic transmission (Paulos et al., Reference Paulos, Köster, de Lucio, Hernández-de-Mingo, Cardona, Fernández-Crespo, Stensvold and Carmena2018).
In this study, a significant association was found between G. duodenalis infection and clinical symptoms such as abdominal pain and loss of appetite. However, no correlation between specific assemblages and symptoms was observed. The relationship between clinical manifestations of giardiasis and G. duodenalis genetic variants remains controversial. Some studies, including large case-control surveys in endemic areas, found no clear associations (Rafiei et al., Reference Rafiei, Baghlaninezhad, Köster, Bailo, Hernández-de-Mingo, Carmena and Rivero-Juárez2020; Messa et al., Reference Messa, Köster, Garrine, Gilchrist, Bartelt, Nhampossa, Massora, Kotloff, Levine, Alonso, Carmena and Mandomando2021), while others have reported genotype-specific symptom profiles. For instance, AI, AII and BIII have been linked with abdominal pain, and AI with weight loss in patients with giardiasis in Iran (Viesy et al., Reference Viesy, Abdi, Haghani, Valizadeh and Mirzaei2020). Similarly, a study in Syria reported a significant association between AII and weight loss in symptomatic patients (Skhal et al., Reference Skhal, Aboualchamat, Al Mariri and Al Nahhas2017). In southern Iran, AII was more frequently linked to abdominal pain, nausea and vomiting (Sarkari et al., Reference Sarkari, Ashrafmansori, Hatam, Motazedian, Asgari and Mohammadpour2012).
This study has several limitations that may have affected the accuracy of the results and the strength of the conclusions. First, faecal samples were initially preserved in 70% ethanol for transport prior to subsequent formalin fixation, which may have reduced the sensitivity of conventional microscopy for the detection of fragile protozoa such as D. fragilis, potentially leading to an underestimation of its true prevalence. Second, the epidemiological data may not be fully representative of Madagascar, as the study relied on a convenience sample of children, adolescents and young women from 4 local institutions affiliated with the NGO Bel Avenir. Third, completion of the questionnaire was limited by logistical and linguistic constraints, as most parents/guardians spoke only Malagasy and many were illiterate, requiring translation that was not always feasible due to temporal unavailability of the Malagasy/French translator. Absences and ethical considerations towards participants who provided samples but declined to be interviewed further reduced representativeness. Finally, the relatively small number of participants in some localities may have reduced statistical power.
Conclusion
This study highlights the considerable burden of GIP infections in infants, children, adolescents and young women in Madagascar. Giardia duodenalis infections were significantly associated with clinical manifestations such as abdominal pain and loss of appetite, but not with diarrhoea. Further research is warranted to determine whether G. duodenalis-positive young children are at increased risk of impaired growth and cognitive development. The study also provides novel insights into the genetic diversity of G. duodenalis and Cryptosporidium spp. Future research should adopt a ‘One Health’ approach, including sampling animal and environmental (e.g. water sources), to more accurately elucidate the transmission routes of enteric protists involved in diarrhoeal diseases.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182026102200.
Acknowledgements
We thank the NGO Bel Avenir, particularly Ms Emèlie Tahinjanahary and Ms Volanjary Madio Hèlene, for their support during the fieldwork.
Author contributions
Conceptualization: M.T.; Data curation: D.C. and M.T.; Funding acquisition: D.C. and M.T.; Investigation: G.T.-V., V.G., P.C.K., A.D. and S.S.; Methodology: A.D., S.S., M.T., M.V.F. and M.G.; Project administration: M.T.; Resources: S.S., D.C. and M.T.; Supervision: S.S., D.C. and M.T.; Validation: S.S., D.C. and M.T.; Writing – original draft: G.T.-V., D.C. and M.T.; Writing – review and editing: G.T.-V., P.C.K., A.D., S.S., D.C. and M.T.
Financial support
This research was funded by the Call for Development Cooperation Projects of the Vice rector for Internationalization and Cooperation of the University of Valencia (Project: COOPUV16/2022). Additional funding was obtained from the Health Institute Carlos III (ISCIII) and the Spanish Ministry of Economy and Competitiveness (project: PI19CIII/00029). G.T.-V. had a predoctoral research contract (ACIF/2021/411) funded by the Generalitat Valenciana (Department of Innovation, Universities, Science and Digital Society) and cofounded by the European Social Fund.
Competing interests
The authors declare that they have no competing interests.
Ethical standards
This study was approved by the Ethics Committee for Research in Human Subjects of the University of Valencia (Ref. H1655289183675, 21 June 2022) and by the Biomedical Research Ethics Committee of the Ministry of Public Health of Madagascar (Ref. CERBM No. 09-MSANS/SG/AMM/CERBM, 14 March 2022) and was conducted in accordance with the principles set out in the Declaration of Helsinki of the World Medical Association and the applicable ethical regulations governing medical research involving human participants. Signed informed consent was obtained from all subjects involved in the study or their parents/legal guardians.



 Open access
Open access