


Non-technical Summary
Some externally shelled cephalopods had uncoiled shapes, resembling straight or slightly curved cones. Among these animals, some species had mineral deposits in their chambered shells that likely occurred during life. These denser components invite comparisons with counterweights, presumably allowing these cephalopods to assume horizontal orientations while floating neutrally in the water. Using computer reconstructions, we explore the practical limits of these mineral deposit distributions and how they could have influenced posture, stability, and maneuverability. Furthermore, we demonstrate that these functional properties are heavily dependent upon the proportions of the soft body of these living animals, with shorter proportions allowing more room for deposits. While shorter body proportions can improve maneuverability, deposit-bearing cephalopods commonly display larger proportions that would be insufficient to allow deviation from a stable, downward-facing orientation.
Introduction
Externally shelled cephalopods were among the earliest active swimmers in Paleozoic seas, displaying a wide range of morphologies and hydromechanical properties throughout their extensive evolutionary history (Arkell et al. Reference Arkell, Furnish, Kummel, Miller, Moore, Schindewolf, Sylvester-Bradley and Wright1957; Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964; Saunders and Shapiro Reference Saunders and Shapiro1986; Holland Reference Holland1987b; Wright et al. Reference Wright, Calloman and Howarth1996; Klug and Korn Reference Klug and Korn2004; Hoffmann et al. Reference Hoffmann, Lemanis, Naglik, Klug, Klug, Korn, De Baets, Kruta and Mapes2015, Reference Hoffmann, Slattery, Kruta, Linzmeier, Lemanis, Mironenko, Goolaerts, De Baets, Peterman and Klug2021; Naglik et al. Reference Naglik, Tajika, Chamberlain, Klug, Klug, Korn, De Baets, Kruta and Mapes2015; Peterman and Ritterbush Reference Peterman and Ritterbush2022a,Reference Peterman and Ritterbushb). By the late Cambrian, externally shelled cephalopods (ectocochleates) evolved a chambered buoyancy apparatus, allowing them to explore new regions of their ecospace (Kröger et al. Reference Kröger, Vinther and Fuchs2011). This innovation presented new opportunities for swimming by increasing buoyancy and alleviating the burdens of their weight (Peterman et al. Reference Peterman, Barton and Yacobucci2019a). During the Ordovician, ectocochleates diversified considerably (Flower Reference Flower1957; Kröger and Zhang Reference Kröger, Servais and Zhang2009; Pohle et al. Reference Pohle, Kröger, Warnock, King, Evans, Aubrechtova, Cichowolski, Fang and Klug2022), expanding their biogeographic distributions, life habits, and habitats (Brett and Walker Reference Brett and Walker2002; Frey et al. Reference Frey, Beresi, Evans, King, Percival, Webby, Paris, Droser and Percival2004; Kröger and Zhang Reference Kröger and Zhang2009; Kröger et al. Reference Kröger, Servais and Zhang2009). Among these cephalopods, orthocones (i.e., those with straight shells) became incredibly successful throughout the Paleozoic. This morphotype characterizes more than 100 genera across several higher clades (Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964), each occupying a broad range of environmental settings (Holland Reference Holland1987b; Kröger et al. Reference Kröger, Servais and Zhang2009) and body sizes (Teichert and Kummel Reference Teichert and Kummel1960; Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964, Holland Reference Holland1987b; Klug et al. Reference Klug, De Baets, Kröger, Bell, Korn and Payne2015, Reference Klug, Fuchs, Pohle, Korn, De Baets, Hoffmann, Ware and Ward2025; Pohle and Klug Reference Pohle and Klug2018). Orthocone cephalopods also displayed high disparity in internal conch morphology. The siphuncle, which was responsible for regulating chamber liquid transport and buoyancy (Denton and Gilpin-Brown Reference Denton and Gilpin-Brown1966), varied considerably in size, shape, and composition between different higher clades (compare Orthocerida, Endocerida, and Actinocerida; Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964; Holland Reference Holland1987b; Kröger Reference Kröger2003). The divider walls within their shells (septa) also varied in shape and spacing (Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964). Furthermore, some groups had mineralized regions within their conchs that could have influenced syn vivo posture and stability (i.e., cameral deposits of orthocerids, and endosiphuncular deposits of endocerids, among others; Flower Reference Flower1955a, Reference Flower1957, Reference Flower1964; Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964; Fischer and Teichert Reference Fischer and Teichert1969; Holland Reference Holland1987b; Blind Reference Blind1991; Seuss et al. Reference Seuss, Mapes, Klug and Nützel2012; Niko et al. Reference Niko, Seuss and Mapes2018; Pohle et al. Reference Pohle, Kröger, Warnock, King, Evans, Aubrechtova, Cichowolski, Fang and Klug2022, Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). Each of these properties imply a diverse range of life habits were assumed by these animals with ostensibly similar conch morphologies. However, the full spectrum of orthocone functional capabilities and hydromechanical constraints is still poorly understood.
In this study, we exclusively focus on cameral deposit–bearing orthocones (Pseudorthocerida and other morphologically similar clades). However, our results are also relevant to the broader orthocone morphotype to some lesser degree. Note that several higher clades of orthocone cephalopods have been collectively referred to as nautiloids (Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964; Holland Reference Holland1987b). However, this subclass is now recognized as paraphyletic and is currently undergoing taxonomic revision (King and Evans Reference King and Evans2019; Pohle et al. Reference Pohle, Kröger, Warnock, King, Evans, Aubrechtova, Cichowolski, Fang and Klug2022).
Ectocochleate Cephalopod Hydromechanics
The shape of the conch (referring to the entire geometry of the shell) has profound consequences for the passive and active physical constraints on swimming (i.e., hydrostatics and hydrodynamics, respectively; Trueman Reference Trueman1940; Raup and Chamberlain Reference Raup and Chamberlain1967; Denton Reference Denton1974; Chamberlain Reference Chamberlain1976, Reference Chamberlain, House and Senior1981, Reference Chamberlain1993; Chamberlain and Westermann Reference Chamberlain and Westermann1976; Saunders and Shapiro Reference Saunders and Shapiro1986; Jacobs and Chamberlain Reference Jacobs, Chamberlain, Landman, Tanabe and Davis1996; Hoffmann et al. Reference Hoffmann, Lemanis, Naglik, Klug, Klug, Korn, De Baets, Kruta and Mapes2015; Naglik et al. Reference Naglik, Tajika, Chamberlain, Klug, Klug, Korn, De Baets, Kruta and Mapes2015; Peterman et al. Reference Peterman, Barton and Yacobucci2019a; Hebdon et al. Reference Hebdon, Ritterbush and Choi2021, Reference Hebdon, Polly, Peterman and Ritterbush2022a,Reference Hebdon, Ritterbush, Choi and Petermanb; Peterman and Ritterbush Reference Peterman and Ritterbush2022a,Reference Peterman and Ritterbushb). Hydrostatics concern buoyancy and the distribution of organismal mass, while hydrodynamics refer to how a particular shape moves through a fluid. An ectocochleate cephalopod has the capacity for neutral buoyancy when its chambered portion (the phragmocone) compensates for the weight of the animal’s shell and soft body (Denton and Gilpin-Brown Reference Denton and Gilpin-Brown1966; Greenwald and Ward Reference Greenwald, Ward, Saunders and Landman1987; Hoffmann et al. Reference Hoffmann, Lemanis, Naglik, Klug, Klug, Korn, De Baets, Kruta and Mapes2015; Peterman et al. Reference Peterman, Barton and Yacobucci2019a). Under this condition, the living cephalopod can effortlessly float in the water column, while using jet propulsion as a primary means of locomotion. The coiling of the conch (or lack thereof) influences the distribution of mass. The syn vivo orientation of any organism can be determined by understanding the relative locations of its hydrostatic centers. That is, the center of buoyancy (the centroid of the volume of water displaced) and the center of mass (the average center of mass location for each component of unique density). A stable orientation occurs when the centers of buoyancy and mass are vertically aligned, due to the mutually opposing forces of buoyancy and weight (Greenwald and Ward Reference Greenwald, Ward, Saunders and Landman1987; Hoffmann et al. Reference Hoffmann, Lemanis, Naglik, Klug, Klug, Korn, De Baets, Kruta and Mapes2015; Peterman et al. Reference Peterman, Barton and Yacobucci2019a; Peterman and Ritterbush Reference Peterman and Ritterbush2022b). Hydrostatic stability is proportionate to the separation between these hydrostatic centers. A hydrostatically stable cephalopod can resist external perturbations, and those produced by its own locomotion (i.e., when the jet thrust vector is misaligned with the hydrostatic centers; Chamberlain Reference Chamberlain, Saunders and Landman1987). However, this condition reduces maneuverability, presenting a classic physical trade-off (Peterman and Ritterbush Reference Peterman and Ritterbush2022b). Hydrodynamic drag is proportionate to the projected area perpendicular to the swimming direction and the degree of conch streamlining (Hebdon et al. Reference Hebdon, Ritterbush and Choi2021). Minimization of hydrodynamic drag can reduce the metabolic demands of locomotion and enable faster acceleration and swimming speeds (Jacobs and Chamberlain Reference Jacobs, Chamberlain, Landman, Tanabe and Davis1996; Hebdon et al. Reference Hebdon, Ritterbush and Choi2021, Reference Peterman and Ritterbush2022b; Peterman and Ritterbush Reference Peterman and Ritterbush2021; Ritterbush and Hebdon Reference Ritterbush and Hebdon2023).
Orthoceratoid Functional Morphology
Orthocone cephalopods with empty phragmocones are considered extremely hydrostatically stable and are unable to considerably modify their downward-facing, vertical postures (Trueman Reference Trueman1940; Westermann Reference Westermann, Landman, Tanabe and Davis1996, Reference Westermann2013; Tsujita and Westermann Reference Tsujita and Westermann1998; Peterman et al. Reference Peterman, Barton and Yacobucci2019a,Reference Peterman, Ciampaglio, Shell and Yacobuccib; Peterman and Ritterbush Reference Peterman and Ritterbush2021). This condition is produced by the air-filled chambers and underlying soft body distributing mass adorally. In contrast, planispiral cephalopods wrap their soft bodies around the phragmocone (ranging from a fraction of a whorl to several whorls), reducing stability, increasing maneuverability, and possibly improving their horizontal motility (Raup and Chamberlain Reference Raup and Chamberlain1967; Saunders and Shapiro Reference Saunders and Shapiro1986; Klug and Korn Reference Klug and Korn2004; Peterman and Ritterbush Reference Peterman and Ritterbush2022b). Orthocones are also capable of relatively high swimming speeds and accelerations in the upward vertical direction due to their streamlined shape and low transverse area (Peterman and Ritterbush Reference Peterman and Ritterbush2021). While general swimming capabilities depend upon propulsive musculature (which varied between orthocone taxa; Kennedy et al. Reference Kennedy, Cobban and Klinger2002; Mutvei Reference Mutvei2002; Klug et al. Reference Klug, Riegraf and Lehmann2012; Doguzhaeva and Mapes Reference Doguzhaeva, Mapes, Klug, Korn, De Baets, Kruta and Mapes2015; Klug and Lehmann Reference Klug, Lehmann, Klug, Korn, De Baets, Kruta and Mapes2015; King and Evans Reference King and Evans2019), orthocones would have incurred little cost to move in the direction of the shell’s long axis. These properties support life habits involving low-energy vertical migration in the water column, with the potential for rapid upward escape maneuvers (Westermann Reference Westermann and Savazzi1998; Peterman and Ritterbush Reference Peterman and Ritterbush2021).
Cameral Deposits
Many cephalopods within the Orthoceratoidea have mysterious calcareous structures filling their chambers (Flower Reference Flower1955a, Reference Flower1957, Reference Flower1964; Fischer and Teichert Reference Fischer and Teichert1969; Blind Reference Blind1991; Seuss et al. Reference Seuss, Mapes, Klug and Nützel2012; Niko et al. Reference Niko, Seuss and Mapes2018; Pohle et al. Reference Pohle, Kröger, Warnock, King, Evans, Aubrechtova, Cichowolski, Fang and Klug2022, Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). These structures are termed “cameral deposits” and can consist of both primary aragonite and calcite that were precipitated syn vivo (Pohle et al. Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). While some earlier work has suggested a diagenetic origin for these deposits (Mutvei Reference Mutvei1956, Reference Mutvei2002, Reference Mutvei2018), a more recent morphogenetic model has been proposed that reconciles the patterns preserved in fossils with practical functional constraints of their once-living counterparts (Pohle et al. Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). This model involves transport of ion-rich liquid via thin organic sheets, similar to the pellicle of ectocochleate cephalopods (a thin conchiolin lining that acted as a wettable, capillary sheet; Denton and Gilpin-Brown Reference Denton and Gilpin-Brown1966; Greenwald and Ward Reference Greenwald, Ward, Saunders and Landman1987). The pellicle and cameral sheets connected the cameral liquid to the siphuncle, the organ spanning all chambers and allowing liquid transport within the phragmocone, via osmosis and active transport of ions (Denton and Gilpin-Brown Reference Denton and Gilpin-Brown1966; Ward and Martin Reference Ward and Martin1978; Greenwald and Ward Reference Greenwald, Ward, Saunders and Landman1987). Thus, cameral liquid can be transported even if liquid is not in direct contact with the siphuncle (i.e., decoupled; Denton and Gilpin-Brown Reference Denton and Gilpin-Brown1966; Ward Reference Ward1979; Greenwald and Ward Reference Greenwald, Ward, Saunders and Landman1987). This osmotic transport mechanism and network of capillary sheets also explain how cameral deposits can form without being entirely submerged in cameral liquid; a condition that would produce negative buoyancy unless the soft-body proportions were sufficiently low (Peterman et al. Reference Peterman, Barton and Yacobucci2019a). Cameral deposits also generally thicken toward the shell apex (Flower Reference Flower1955a, Reference Flower1957, Reference Flower1964; Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964; Fischer and Teichert Reference Fischer and Teichert1969; Blind Reference Blind1991), suggesting they continue to grow throughout the life span of an individual. This distribution of deposits also may have imposed constraints on stability by distributing more mass apically compared to the same mass of water (due to their comparatively higher density and different spatial distributions; Peterman et al. Reference Peterman, Barton and Yacobucci2019a).
While the growth model of Pohle et al. (Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025) improves our understanding of cameral deposit morphogenesis, the adaptive value of these structures remains enigmatic. The cameral sheet network may have improved liquid transport rates, and the apical infilling of chambers would have reduced the effective volume of the phragmocone, with both factors potentially increasing growth rates (Pohle et al. Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). Additionally, the dense cameral deposits would have influenced the distribution of organismal mass to some extent and therefore may have had hydrostatic consequences. Earlier work suggested that cameral deposits moved the center of mass apically and ventrally, allowing a horizontal posture in the water column (Flower Reference Flower1955a,Reference Flowerb, Reference Flower1957, Reference Flower1964; Fischer and Teichert Reference Fischer and Teichert1969; Westermann Reference Westermann1977; Crick Reference Crick1988; Chamberlain Reference Chamberlain1993). While such a condition can be inferred based on the apical and ventral distribution of these dense crystalline fabrics (Fig. 1A, Supplementary Fig. S1), the living animal must also maintain a delicate balancing act by maintaining near-neutral buoyancy. Additionally, the soft body (which is more voluminous than any other component) and the air-filled voids (which are around three orders of magnitude less dense than any other component) both strongly attract and repel the total center of mass, respectively (Peterman et al. Reference Peterman, Barton and Yacobucci2019a,Reference Peterman, Ciampaglio, Shell and Yacobuccib; Peterman and Ritterbush Reference Peterman and Ritterbush2022b). These conditions prescribe harsh physical constraints that are hard to overcome. More recent virtual models (Peterman et al. Reference Peterman, Barton and Yacobucci2019a) demonstrate that cameral deposits may have only slightly reduced hydrostatic stability, possibly producing a slightly more maneuverable condition compared with orthocones with empty shells (Peterman et al. Reference Peterman, Barton and Yacobucci2019a,Reference Peterman, Ciampaglio, Shell and Yacobuccib; Peterman and Ritterbush Reference Peterman and Ritterbush2022b). Furthermore, asymmetrical distributions of these deposits did not considerably influence the static orientation of these models (less than a few degrees; Peterman et al. Reference Peterman, Barton and Yacobucci2019a). However, these virtual models only represented theoretical morphologies with a prescribed body chamber ratio (BCR) of one-third total length and very generalized shapes as an attempt to isolate variables. Additionally, the orthocone morphotype provides more mechanical advantage for rotation compared with others (Peterman et al. Reference Peterman, Ciampaglio, Shell and Yacobucci2019b). That is, the large distance between the source of jet thrust (the hyponome) and the hydrostatic centers produces a larger lever arm (D hm; Fig. 1B), generating more torque for some value of jet thrust, possibly enabling postures other than vertical. For orthocones with empty shells, this increased mechanical advantage is overshadowed by their high hydrostatic stability (Peterman et al. Reference Peterman, Ciampaglio, Shell and Yacobucci2019b). However, cameral deposits have the potential to reduce hydrostatic stability to a greater degree than the length of this hyponome lever arm. Ultimately, the influence of conch morphology, cameral deposit distributions, and body chamber proportions on the hydromechanical properties of these living animals still remains poorly understood.
Schematic showing digital model construction from cameral deposit–bearing orthocones. A, Pseudorthoceratid cephalopod (Smithorthoceras unicamera; AMNH-FI-145916) showing asymmetrical distribution of cameral deposits (note venter is figured toward the right). B, Reconstructed orthocone with cameral deposits (purple) thickening apically. The soft-body (green) proportions can be altered in these models to explore the variability of key hydrostatic properties. The purple arrow denotes the transition point between cameral deposit–bearing phragmocone and empty phragmocone. The green arrow denotes the transition between the phragmocone and body chamber. Measurements related to maneuverability are displayed next to the model (D hm, the distance between the hyponome and center of mass; D bm, the distance between the centers of buoyancy and mass). Enlarging D hm and reducing D bm decrease stability while increasing maneuverability. C, Enlarged view of center of buoyancy (COB; tip of blue cone), and center of mass (COM; tip of red cone). D, Enlarged medial view of the shell (white) containing cameral deposits (purple) and siphuncle (green).

In the current study, we use exceptionally preserved fossils of pseudorthocerid cephalopods (Spyroceratinae) from the Buckhorn Asphalt (Middle Pennsylvanian, Boggy Formation, Oklahoma, USA; Taff Reference Taff1899; Seuss et al. Reference Seuss, Nützel, Mapes and Yancey2009) to generate virtual hydrostatic models. Furthermore, we modify these models to explore the limits of cameral deposit distributions within the phragmocone under several scenarios: asymmetric versus symmetric cameral deposit distributions, entirely apical versus entirely ventral deposit distributions, low versus high shell taper (i.e., expansion rate), orthocone (straight) versus cyrtocone (slightly curved) morphologies, and several different body chamber proportions for each of these conditions (producing 43 models in total). Finally, we compare these models with fossil orthoceratoids and other ectocochleates to understand whether cameral deposits influenced static orientations, stability, and maneuverability in these living animals. These experiments shed light on the adaptive value of cameral deposits, the broader spectrum of functional possibilities for early stem-group cephalopods, and the life habits of ubiquitous, ecologically significant animals that once dominated many Paleozoic marine communities.
Materials and Methods
Materials
The measured specimens are all from the Middle Pennsylvanian Buckhorn Asphalt (Boggy Formation, Oklahoma, USA). We focused on several genera within Spyroceratinae (Pseudorthoceratidae) to somewhat isolate variables of shell/septum thickness, septal spacing, and siphuncular anatomy, which vary to some degree within cameral deposit–bearing orthocones. These specimens include Smithorthoceras unicamera (Smith, Reference Smith1938), Arbuckleoceras tricamerae (Smith, Reference Smith1938), cf. Sulphurnites taffi Niko et al. Reference Niko, Seuss and Mapes2018 (see Supplementary Figs. S1–S5). These specimens were commonly associated with Dolorthoceras boggyense (Niko et al. Reference Niko, Seuss and Mapes2018); however, this species was excluded due to its differences in septal spacing and cameral deposit shape. A fragment of a cyrtocone (with slight to moderate curvature) was also uncovered while preparing a bulk sample of this material (AMNH-FI-145902; Supplementary Fig. S4). This specimen seems to have more curvature compared with other cyrtocones reported from the Boggy Formation (compare Unklesbayoceras; Niko et al. Reference Niko, Seuss and Mapes2018). There is no obvious indication that this specimen has been deformed to intensify its curvature. Curvature is also assumed to be exogastric based on the thicker cameral deposits on the convex side (Supplementary Fig. S4). Due to uncertainty in taxonomic affinity, we use the broad term “cyrtocone” when referring to this specimen. This specimen (AMNH-FI-145902) deserves more scrutiny in taxonomic work involving the Boggy Formation, or Pennsylvanian cephalopods more broadly.
Longitudinal sections of specimens were prepared to reveal the contents of their chambers and conch morphologies (i.e., cameral deposit distribution, shell and septal thickness, septal spacing, siphuncle size; each as a function of whorl height). Fourteen specimens were prepared (six of which were left in the matrix, yielding more accurate measurements of shell thickness; Supplementary Table S1). Specimens were placed in small silicone molds, oriented latitudinally, and impregnated with Buehler EpoThin2 Epoxy System. Once cured, the specimens were ground to approximately longitudinal midline using 8-inch perforated medium- and fine-grade diamond polishing wheels (220 grit and 600 grit, respectively). Specimens were then illuminated using diffused LED lighting and photographed using a Nikon D7500 DSLR camera, with a Nikon AF-S DX Micro-NIKKOR 40mm f/2.8G close-up lens. An additional specimen (AMNH-FI-145916) was ground to a thickness that would allow transmitted light to penetrate. This sectioned specimen was adhered to frosted geological thin section slides using the EpoThin 2 Epoxy System and placed in a 60 PSI pressure chamber to cure overnight (to prevent bubbles from forming). This specimen was photographed before and after staining with Feigl’s method (Ayan Reference Ayan1965) to distinguish between aragonite and calcite mineralogy. It was photographed with transmitted light using a Stack Shot device coupled with a Nikon D7500 DSLR camera fit with a 4× Nikon Plan microscope objective. A composite photograph was taken using Helicon stacking software.
Virtual Model Generation
An orthocone model was created by following the taper of a specimen with five consecutive chambers (AMNH-FI-138966; S. unicamera; Supplementary Fig. S2). A photo of this longitudinal section was imported into the computer modeling software Blender (Blender Development Team 2021) and positioned on the X-Z plane. The adoralmost portion of the model was scaled to 2 cm, a presumably adult aperture size estimated from AMNH-FI-160642. A circular whorl section was created with this diameter, and an array (Supplementary Table S2) was used to copy, translate, and scale this whorl section until it was 2.1 mm in diameter (i.e., 1120 times; Supplementary Table S2). The embryonic shell of Pseudorthoceras knoxense (figured in Kröger and Mapes Reference Kröger and Mapes2004: fig. 1.7), was used to reconstruct the remaining portion with two separate arrays (Supplementary Table S2). Reconstruction of the embryonic portion accounted for the higher degree of allometry at this stage and reduced the overall length compared with a perfectly isometric condition (which is also relevant for computing the body chamber length and is especially sensitive for a shell with such a low degree of taper).
The cyrtocone specimen recovered from this material was used as a basis for exploring the influence of conch curvature on hydrostatics. This specimen has presumably exogastric curvature (i.e., convex venter), based on the thicker cameral deposits on the convex side. This specimen presents an opportunity to compare the hydrostatic influence of conch curvature and higher whorl expansion for orthoceratoids containing cameral deposits. An isometric shell was constructed by using a longitudinal section of this specimen as a stencil and scaling the section to the same adult size as the low-taper orthocone (2 cm; array instructions in Supplementary Table S2). A straight version of this cephalopod was constructed to compare the influence of whorl expansion (i.e., shell taper) on orthocone hydrostatics. This version was made by setting the rotation factor in the array instructions to zero, while holding all other parameters constant (Supplementary Table S2).
Shell thickness was defined by copying the whorl section at the aperture and scaling it down to produce a thickness of 3.211% of whorl height. This number represents an average computed from measurements from each chamber of the 14 specimens (Supplementary Tables S1, S3). All replicated whorl sections were bridged together to form complete surfaces representing the exterior and interior of the shell. These two surfaces were joined and filled at the aperture to make a 3D volume representing the shell.
Septa were constructed as hemispheres that match the curvature of the cross-sectioned specimens. Septal thickness increased toward the center of the septum. This thickness taper was matched with specimen AMNH-FI-138966 (S. unicamera; Supplementary Fig. S2). Measurements of septal thickness at the midpoint between the siphuncle and shell wall (listed in Supplementary Tables S1, S3) closely match the corresponding thicknesses of the modeled septa. A septal neck was constructed from cylinders after adjusting the diameter of each circular face to match the cross section of AMNH-FI-138966. This septal neck was joined with the septum using a Boolean union in Blender (Blender Development Team 2021). A single septum was replicated with the same array instructions used to build each shell (Supplementary Table S2). However, 17 replications were skipped to produce the same septal spacing in AMNH-FI-138966. Later, septal foramina were created by subtracting a model of the siphuncle from all septa using the Boolean operations in Netfabb (v. 2023, Autodesk Inc.). These septa were unified to the shell tube using Netfabb to produce a complete model of the conch.
A siphuncle was created by making a circle with the normalized siphuncular diameter reported in Supplementary Table S3 (15.203% of whorl height). This circle was replicated with the same array instructions used to build each shell.
The soft body was constructed by isolating the interior surface of the body chamber and unifying it with the siphuncle. A model with a conservative head and 10 arms was then unified in Blender (Blender Development Team 2021) to produce the entire soft body of the reconstructed cephalopod. Different body chamber proportions were modeled to investigate the role of this parameter on the distribution of cameral deposits and resultant hydrostatic properties.
The interior surfaces of the phragmocone were isolated to make a model representing the cameral volumes. This model was later modified to specify the distribution of chamber contents (i.e., cameral liquid or cameral deposits).
Finally, a model representing the water displaced by the living animal was constructed by isolating all external surfaces of the model (i.e., the soft body protruding from the aperture and the shell exterior). This model is required for hydrostatic calculations (i.e., determining buoyancy, the center of buoyancy location, and hydrostatic stability).
Description of Virtual Hydrostatic Models
The previously outlined procedure was followed for all three shell types: (1) a low-taper orthocone resembling an S. unicamera (Smith Reference Smith1938) with an apical angle of 1.146° through most of its ontogeny; (2) a high-taper orthocone (straight version of case 3; apical angle of 2.576° through all of its ontogeny; Supplementary Table S1); and (3) a high-taper, exogastric cyrtocone resembling AMNH-FI-145902 (Supplementary Fig. S4). For each of these three conch morphologies, different BCRs were modeled (i.e., body chamber length measured from shell aperture to the septal suture line, normalized by total conch length; BCR): (1) 40%, (2) 30%, (3) 25%, (4) the percentage required to make the centers of mass and buoyancy nearly coincide, and (5) 20%. Note that lengths on the cyrtocone were measured as curvilinear lengths (i.e., similar to measuring a length of string after straightening).
The low-taper orthocones, with each BCR mentioned earlier, were filled with cameral deposits that met the conditions for neutral buoyancy (described in the next section). Asymmetric (ventrally thicker) and symmetric (dorsoventrally and laterally) cameral deposits were created following the taper angles displayed in longitudinal sections of AMNH-FI-145916 (Fig. 1, Supplementary Fig. S1) and AMNH-FI-138966 (Supplementary Fig. S2), respectively. These cameral deposits were modeled by subtracting 3D cones with the appropriate taper angle from the model representing the cameral volumes. For all cameral deposits, Boolean operations were performed in Netfabb (v. 2023, Autodesk Inc.) to produce 3D models with the appropriate volumes (detailed later).
Cameral deposits in the high-taper orthocones were scaled according to the differences in shell taper angles. That is, if their cameral deposits tapered as gradually as the low-taper orthocone models, they would meet the adoralmost septum. Therefore, they must taper more quickly to produce a more realistic condition, consistent with observed specimens and the most current growth model (Pohle et al. Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). Only theoretical (symmetric, apical, ventral, and oblique) deposits were modeled for these theoretical morphologies, because asymmetry would have little influence in mass distribution. That is, the higher taper of the cameral deposits produces a quick gradient between empty chambers and completely filled chambers. The cyrtocone models have the same properties as the high-taper orthocone (aside from its nonzero rotation value; Supplementary Table S2). Therefore, its cameral deposits were modeled with the same conditions as the high-taper orthocone.
For BCRs of 30% and 25% (a realistic range based on minimum estimates from cameral deposit–bearing pseudorthocerids; Supplementary Table S4), cameral deposit distributions were pushed to their extremes in the apical, ventral, and oblique directions. Apical deposits were created by removing the adoral region of the cameral volumes until the target volume was reached. Ventral deposits were created by the same means, but first, all camerae were halved (i.e., dorsal side removed). Oblique deposits are distributed apically and ventrally. These models were made by subtracting a rectangular prism of arbitrary dimensions (large enough to encompass the shell). An edge of this prism was placed at the second most recent suture of the conch, then its angle was adjusted until subtracting this prism produced the desired volume of cameral deposits. This condition was chosen based on practical constraints on cameral deposit formation (i.e., formation in the most recent chamber would probably be too early for this process to occur). Similar extreme distributions were modeled for the cyrtocone model, distributing deposits apically, on the concave half of the chambers (dorsum), and the convex half of the chambers (venter).
Calculating Hydrostatic Properties
Cameral deposits were modeled to fill the chamber volumes for several different scenarios: (1) dorsoventrally symmetric, (2) ventrally asymmetric, (3) apically distributed, (4) ventrally distributed, and (5) oblique (i.e., distributed toward the apex and venter). First, the volume of cameral deposits (
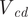 $ {V}_{cd} $
) required for neutral buoyancy was solved by using the volumes and assigned densities of the previously modeled components (shell, soft body, cameral volumes, and volume of water displaced):
$ {V}_{cd} $
) required for neutral buoyancy was solved by using the volumes and assigned densities of the previously modeled components (shell, soft body, cameral volumes, and volume of water displaced):
 $$ {V}_{cd}=\frac{V_{wd}{\rho}_{wd}-{V}_{sb}{\rho}_{sb}-{V}_{sh}{\rho}_{sh}}{\rho_{cd}} $$
$$ {V}_{cd}=\frac{V_{wd}{\rho}_{wd}-{V}_{sb}{\rho}_{sb}-{V}_{sh}{\rho}_{sh}}{\rho_{cd}} $$
where V and
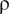 $ \unicode{x03C1} $
represent volume and density, respectively. The subscripts refer to each model component (cd = cameral deposits, wd = water displaced, sb = soft body, and sh = shell).
$ \unicode{x03C1} $
represent volume and density, respectively. The subscripts refer to each model component (cd = cameral deposits, wd = water displaced, sb = soft body, and sh = shell).
The percentage of the phragmocone filled with cameral deposits to satisfy a neutrally buoyant condition (
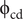 $ {\unicode{x03D5}}_{\mathrm{cd}} $
) was computed with the following equation:
$ {\unicode{x03D5}}_{\mathrm{cd}} $
) was computed with the following equation:
 $$ {\phi}_{cd}=\frac{\left(\frac{V_{wd}{\rho}_{wd}-{V}_{sb}{\rho}_{sb}-{V}_{sh}{\rho}_{sh}}{V_{ct}}\right)-\left({\rho}_{cg}\right)}{\left({\rho}_{cd}-{\rho}_{cg}\right)}\ast 100 $$
$$ {\phi}_{cd}=\frac{\left(\frac{V_{wd}{\rho}_{wd}-{V}_{sb}{\rho}_{sb}-{V}_{sh}{\rho}_{sh}}{V_{ct}}\right)-\left({\rho}_{cg}\right)}{\left({\rho}_{cd}-{\rho}_{cg}\right)}\ast 100 $$
where
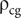 $ {\unicode{x03C1}}_{\mathrm{cg}} $
is the density of the cameral gas (0.001 g/cm3), and
$ {\unicode{x03C1}}_{\mathrm{cg}} $
is the density of the cameral gas (0.001 g/cm3), and
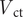 $ {V}_{\mathrm{ct}} $
is the total volume of all empty camerae.
$ {V}_{\mathrm{ct}} $
is the total volume of all empty camerae.
For comparisons, some models were constructed with no cameral deposits; instead, some amount of cameral liquid was required to produce a neutrally buoyant condition. The percentage of cameral liquid in the phragmocone required for this condition (
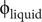 $ {\unicode{x03D5}}_{\mathrm{liquid}} $
) was computed with the following equation:
$ {\unicode{x03D5}}_{\mathrm{liquid}} $
) was computed with the following equation:
 $$ {\unicode{x03D5}}_{\mathrm{liquid}}=\frac{\left(\frac{V_{wd}{\rho}_{wd}-{V}_{sb}{\rho}_{sb}-{V}_{sh}{\rho}_{sh}}{V_{ct}}\right)-\left({\rho}_{cg}\right)}{\left({\rho}_{cl}-{\rho}_{cg}\right)}\ast 100 $$
$$ {\unicode{x03D5}}_{\mathrm{liquid}}=\frac{\left(\frac{V_{wd}{\rho}_{wd}-{V}_{sb}{\rho}_{sb}-{V}_{sh}{\rho}_{sh}}{V_{ct}}\right)-\left({\rho}_{cg}\right)}{\left({\rho}_{cl}-{\rho}_{cg}\right)}\ast 100 $$
where
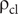 $ {\unicode{x03C1}}_{\mathrm{cl}} $
is the density of cameral liquid (set equal to that of seawater; Greenwald and Ward Reference Greenwald, Ward, Saunders and Landman1987).
$ {\unicode{x03C1}}_{\mathrm{cl}} $
is the density of cameral liquid (set equal to that of seawater; Greenwald and Ward Reference Greenwald, Ward, Saunders and Landman1987).
The static orientation of each model (i.e., that assumed by the living animal) and its hydrostatic stability values were determined by computing the 3D locations of the centers of buoyancy (B) and mass (M). The center of buoyancy is equal to the center of volume of the model representing the water displaced by the living animal (i.e., its centroid). The total center of mass location depends on the local centers of each material of unique density.
 $$ M=\frac{\sum \left(L{m}_i\right)}{\sum {m}_i} $$
$$ M=\frac{\sum \left(L{m}_i\right)}{\sum {m}_i} $$
where M is the total center of mass in a principal direction, L is the local center of mass measured from an arbitrary datum (the center of the aperture), and mi is the mass of each modeled object of unique density. The 3D coordinates of each center were computed in Meshlab (Cignoni et al. Reference Cignoni, Callieri, Corsini, Dellepiane, Ganovelli, Ranzuglia, Scarano, De Chiara and Erra2008). For models containing cameral liquid, the centers of mass were set to the center of volume of all chambers. This is a reasonable assumption based on the capillary retention of liquid around the septal margins in the living animals (Ward and Chamberlain Reference Ward and Chamberlain1983; Peterman et al. Reference Peterman, Ritterbush, Ciampaglio, Johnson, Inoue, Mikami and Linn2021). The modeled densities for each material are as follows: water displaced and cameral liquid (1.025 g/cm3), soft body (1.049 g/cm3), shell and cameral deposits (2.54 g/cm3), and cameral gas (0.001 g/cm3).
The hydrostatic stability index (St) was computed from the locations of B and M:
 $$ {S}_t=\frac{\;\sqrt{{\left({B}_x-{M}_x\right)}^2+{\left({B}_y-{M}_y\right)}^2+{\left({B}_z-{M}_z\right)}^2}}{\sqrt[3]{V_{wd}}} $$
$$ {S}_t=\frac{\;\sqrt{{\left({B}_x-{M}_x\right)}^2+{\left({B}_y-{M}_y\right)}^2+{\left({B}_z-{M}_z\right)}^2}}{\sqrt[3]{V_{wd}}} $$
The subscripts x, y, and z correspond to the 3D coordinates of each hydrostatic center. Note that all models are bilaterally symmetric, and experience negligible variations in the y direction.
When B and M are aligned vertically, the living animal is in equilibrium. This static orientation was measured as the angle between the aperture and the vertical axis (
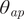 $ {\theta}_{ap} $
). Thus, angles of −90° and +90° denote downwardly and upwardly facing animals, respectively. Angles of zero denote a horizontally facing aperture.
$ {\theta}_{ap} $
). Thus, angles of −90° and +90° denote downwardly and upwardly facing animals, respectively. Angles of zero denote a horizontally facing aperture.
The distance between the hydrostatic centers (
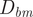 $ {D}_{bm} $
) represents a lever arm corresponding to the restoring torque (i.e., gravity acting on the organismal mass and buoyancy acting in the opposite direction). The distance between the hyponome (source of jet thrust) and the center of mass (
$ {D}_{bm} $
) represents a lever arm corresponding to the restoring torque (i.e., gravity acting on the organismal mass and buoyancy acting in the opposite direction). The distance between the hyponome (source of jet thrust) and the center of mass (
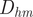 $ {D}_{hm} $
) represents another lever arm that grants mechanical advantage while rotating away from the static equilibrium orientation. Therefore, the ratio between these two lever arms (Fig. 1) is a scalable, dimensionless quantity with higher values representing more mechanical advantage for changing posture in the water column (i.e., more maneuverability):
$ {D}_{hm} $
) represents another lever arm that grants mechanical advantage while rotating away from the static equilibrium orientation. Therefore, the ratio between these two lever arms (Fig. 1) is a scalable, dimensionless quantity with higher values representing more mechanical advantage for changing posture in the water column (i.e., more maneuverability):
 $$ \mathrm{Lever}\;\mathrm{arm}\hskip1em \mathrm{ratio}=\frac{D_{hm}}{D_{bm}} $$
$$ \mathrm{Lever}\;\mathrm{arm}\hskip1em \mathrm{ratio}=\frac{D_{hm}}{D_{bm}} $$
Note that ratios of one or greater do not necessarily mean that these living cephalopods could practically overcome their restoring moments (which increase to a maximum at an angle of 90° from their static orientation). That is, the thrust produced by a cephalopod is often a fraction of its own weight (0.2 for extant Nautilus; Chamberlain Reference Chamberlain1990). Therefore,
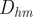 $ {D}_{hm} $
might have to be many times larger than
$ {D}_{hm} $
might have to be many times larger than
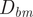 $ {D}_{bm} $
to considerably change orientation. Aside from unknown propulsive potential and its variability within ectocochleates, this lever arm ratio can still be used to gauge relative differences in mechanical advantage between different morphologies.
$ {D}_{bm} $
to considerably change orientation. Aside from unknown propulsive potential and its variability within ectocochleates, this lever arm ratio can still be used to gauge relative differences in mechanical advantage between different morphologies.
Comparison of Proportions with Extant Nautilus.
A computer model of an extant Nautilus pompilius Linnaeus, Reference Linnaeus1758 from an earlier study (Peterman et al. Reference Peterman, Barton and Yacobucci2019a) was used as a point of comparison for linear BCRs in orthocone cephalopods. This model was unraveled to form an orthocone cephalopod using Blender. Maintaining the relative location of the terminal septum produced an orthocone around 57 cm long, with a linear BCR of 37.2%.
Results
All measurements required to compute hydrostatic properties are listed in Supplementary Tables S5 and S6 (i.e., centroids, volumes, and masses for each model component). The digitally reconstructed orthocones and cyrtocones enable estimations of total conch size. A low-taper (1.146°) orthocone resembling Smithorthoceras would have a total conch length of 45.5 cm, assuming a 2-cm-wide aperture, and a juvenile portion (last 1.8%; Supplementary Table S2) that tapers more rapidly than the rest of the shell. Assuming neutral buoyancy, cephalopods with these proportions would have weighed around 56 g (Supplementary Table S6). Isometric high-taper (2.576°) orthocones and cyrtocones would have conch lengths (linear and curvilinear, respectively) of 21.5 cm and would have weighed ~26.4 g (Supplementary Table S6), with the same adult aperture size of 2 cm.
Hydrostatics
Under a neutrally buoyant condition, as the BCR decreases, more mass can be dedicated to the chamber contents (cameral liquid or calcareous cameral deposits, etc.; Table 1). That is, these chamber contents have more influence on the mass distribution (and therefore orientation and stability) for cephalopods with shorter BCRs. The following sections describe hydrostatic properties for 43 theoretical morphologies, designed to test the practical limits of posture and stability for orthoceratoid cephalopods (Figs. 2–8, Table 1).
Hydrostatics and computed proportions for each experimental condition (see Figs. 2–8). BCR, body chamber ratio (body chamber length normalized by total conch length); Φcd, percentage of the phragmocone occupied by cameral deposits; D bm, distance between the centers of buoyancy and mass; St, hydrostatic stability index; θap, apertural angle in degrees (measured from the vertical; i.e., −90° is downwardly facing); D hm, distance between the hyponome (source of jet thrust) and the center of mass; D hm/D bm, lever arm ratio (a dimensionless index representing maneuverability).

Hydrostatic models of low-taper orthocones with asymmetric, ventrally heavy cameral deposits. All models are displayed with their proper static orientations A, Body chamber ratio (BCR) of 30%; B, BCR of 25%; C, BCR of 24.3%; and D, BCR of 20%. E, Close-up view of medial section showing the taper and distribution of cameral deposits. Model components are color coded depending on their bulk density values (green, soft body; purple, cameral deposits; gray, shell). The tips of the blue and red cones denote the centers of buoyancy and mass, respectively. To improve visibility, arrows are located adjacent to the model. The blue and red arrows point toward the centers of buoyancy and mass. The green and purple arrows denote the end of the soft body and cameral deposits, respectively. All models are scaled with adult aperture diameters of 2 cm.

Low-Taper Orthocones with Asymmetrical Cameral Deposits.
Ventrally heavy cameral deposits, resembling those preserved in medial sections (Fig. 1A, Supplementary Fig. S1) do not considerably move the center of mass ventrally (Fig. 2). With a BCR of 30%, the orthocone model experiences a stable (St = 0.278), downward-facing orientation (Table1). The largest deviation from this default, vertical posture in the current models (7.7°; Fig. 2C, Table 1) occurs when the BCR is 24.3% (the proportions that would produce nearly zero hydrostatic stability with symmetrical deposits, detailed in the next section). A shorter BCR of 20% would produce an upward-facing vertical posture with relatively high hydrostatic stability (St = ~0.2; Table 1).
Low-Taper Orthocones with Symmetrical Cameral Deposits.
The maximum BCR possible under a neutrally buoyant condition (with the measured taper angles, septal spacing, and shell and septal thicknesses; Supplementary Table S1) is about 40% (Fig. 3A, Table 1, Supplementary Table S6). This condition produces an empty conch without cameral deposits or cameral liquid. This BCR also produces a maximum in hydrostatic stability (St = 0.668; Table 1, Fig. 3A) with a downward-facing, vertical posture. Five percent decreases in BCR (to 30% and 25%), considerably reduce hydrostatic stability (~0.277 and 0.040, respectively; Table 1, Fig. 3B,C) due to the apical distribution of mass by larger volumes of cameral deposits (Supplementary Table S6). Hydrostatic stability decreases to a minimum around a BCR of 24.3%. This proportion causes the centers of mass and buoyancy to nearly coincide. A 20% BCR produces an upward-facing vertical orientation (Fig. 3E).
Hydrostatic models of low-taper orthocones with symmetric cameral deposits. All models are displayed with their proper static orientations A, Body chamber ratio (BCR) of 40% (note there is no room for cameral deposits, which would produce negative buoyancy at this ratio); B, BCR of 30%; C, BCR of 25%; D, BCR of 24.3%; and E, BCR of 20%. F, Close-up view of medial section showing empty phragmocone; and G, close-up view of medial section showing symmetric cameral deposits in phragmocone. Model components are color coded depending on their bulk density values (green, soft body; purple, cameral deposits; gray, shell). The tips of the blue and red cones denote the centers of buoyancy and mass, respectively. To improve visibility, arrows are located adjacent to the model. The blue and red arrows point toward the centers of buoyancy and mass. The green and purple arrows denote the end of the soft body and cameral deposits, respectively. All models are scaled with adult aperture diameters of 2 cm.

Low-Taper Orthocones with Theoretical Distributions of Chamber Contents.
Theoretical morphologies allow the limits of cameral deposit distributions to be explored. BCRs of 30% and 25% were explored with these theoretical distributions (Fig. 4). These ratios were chosen based on minimum estimates from closely related orthoceratoids (Supplementary Table S4), and because conchs with larger ratios (approaching 40%, see previous section) cannot accommodate any extra mass in their chambers. A BCR of 30% would require 27.6% of the phragmocone volume to be filled with liquid for a neutrally buoyant condition (or 11.2% cameral deposits; Table 1). At this BCR, an even distribution of cameral liquid produces about twice the hydrostatic stability compared with conchs with lifelike cameral deposits (Table 1, Fig. 4A vs. Fig. 2A). A BCR of 25% would require 38.6% of the phragmocone to be filled with liquid (or 15.7% cameral deposits; Table 1). An even distribution of liquid produces around an order of magnitude higher stability compared with the same conch with lifelike cameral deposits (Table 1, Fig. 4E vs. Fig. 2B). Apically distributing cameral deposits optimally moves the center of mass in the apical direction (Fig. 4B,F). At a BCR of 30%, this condition only slightly reduces hydrostatic stability (Table 1, Fig. 4B vs. Fig. 2A). However, at a BCR of 25%, apically distributing cameral deposits reduces hydrostatic stability by an order of magnitude and reverses the static orientation to upward-facing (Table 1, Fig. 4F vs. Fig. 2B). Confining the cameral deposits to the ventral side of the siphuncle only slightly moves the center of mass in the ventral direction (Fig. 4C,G, Supplementary Table S5). The oblique deposit models more optimally move the center of mass in this direction, because they allow the cameral deposits to occupy a larger portion of the ventral wall of the phragmocone (Fig. 4D,H, Supplementary Table S5). These oblique distributions produce the largest deviations from vertical orientations among the corresponding 30% and 25% BCR models (~4° and ~0.7°, respectively; Table 1, Fig. 4D,H).
Hydrostatic models of low-taper orthocones with theoretical distributions of chamber contents. All models are displayed with their proper static orientations A, Body chamber ratio (BCR) of 30% with an even distribution of liquid in the phragmocone (27.6% of all chamber volumes); B, BCR of 30% with an apical distribution of cameral deposits; C, BCR of 30% with a ventral distribution of cameral deposits; D, BCR of 30% with an oblique distribution of cameral deposits (ventral and apical direction); E, BCR of 25% with an even distribution of liquid in the phragmocone (38.6% of all chamber volumes); F, BCR of 25% with an apical distribution of cameral deposits; G, BCR of 25% with a ventral distribution of cameral deposits; and H, BCR of 25% with an oblique distribution of cameral deposits. Model components are color coded depending on their bulk density values (green, soft body; purple, cameral deposits; blue, cameral liquid; gray, shell). The tips of the blue and red cones denote the centers of buoyancy and mass, respectively. To improve visibility, arrows are located adjacent to the model. The blue and red arrows point toward the centers of buoyancy and mass. The green and purple arrows denote the end of the soft body and cameral deposits, respectively. All models are scaled with adult aperture diameters of 2 cm.

High-Taper Orthocones with Symmetrical Cameral Deposits.
Increasing the taper angle of the conch from 1.146° to 2.576° approximately halves the hydrostatic stability of the empty-phragmocone models (Table 1, Fig. 5A vs. Fig. 3A). Note that maintaining a 40% BCR conch (a linear ratio), produces different soft-body volume to0 total volume ratios, which is more relevant for the distribution of mass. Increasing the taper angle of the conch and maintaining a linear BCR increase this volume ratio from 67.7% to 72.1% (Supplementary Table S6), distributing less mass in the adoral direction. Note that a considerable portion of the soft body lies adapically of the hydrostatic centers and is responsible for this reduction in hydrostatic stability. Similar hydrostatic stabilities occur at a BCR of 30% between the low- and high-taper models (Fig. 3B vs. Fig. 5B, Supplementary Table S5). Hydrostatic stability decreases to a minimum as the BCR approaches 22.365% (Table 1, Fig. 5D). Note that a very minor reduction in this ratio could reverse static orientation. Reducing the BCR to 20% produces an upward orientation that is not as stable as its low-taper counterpart (Table 1, Fig. 5E vs. Fig. 3E).
Hydrostatic models of high-taper orthocones with symmetric cameral deposits. All models are displayed with their proper static orientations A, Body chamber ratio (BCR) of 40% (note there is no room for cameral deposits, which would produce negative buoyancy at this ratio); B, BCR of 30%; C, BCR of 25%; D, BCR of 22.365%; and E, BCR of 20%. F, Close-up view of medial section showing empty phragmocone; and G, close-up view of medial section showing symmetric cameral deposits in phragmocone. Model components are color coded depending on their bulk density values (green, soft body; purple, cameral deposits; gray, shell). The tips of the blue and red cones denote the centers of buoyancy and mass, respectively. To improve visibility, arrows are located adjacent to the model. The blue and red arrows point toward the centers of buoyancy and mass. The green and purple arrows denote the end of the soft body and cameral deposits, respectively. All models are scaled with adult aperture diameters of 2 cm.

High-Taper Orthocones with Theoretical Distributions of Chamber Contents.
Theoretical distributions of chamber contents in the high-taper models often produce similar conditions to the corresponding low-taper models (i.e., even liquid; apical, ventral, and oblique cameral deposits; Table 1, Fig. 6 vs. Fig. 4). The models with even distributions of liquid in their phragmocones have a lower percentage of their chambers filled with liquid compared with their low-taper counterparts at BCRs of 30% and 25% (18.8% and 32.7%, respectively; Supplementary Table S6). This reduction is due to the larger ratio of the soft-body volume to total volume when holding the linear BCR constant. Despite this reduction in chamber liquid, hydrostatic stability decreases due to the influence of the soft body on the mass distribution (however, hydrostatic stability is still rather large in these models). Like the low-taper models (Fig. 4), apically distributing cameral deposits produces the greatest reduction in hydrostatic stability for a particular BCR (Table 1, Fig. 6). Likewise, the oblique distributions produce the greatest tilt from a vertical postures, but only <3° (Table 1, Figs. 6D,H).
Hydrostatic models of high-taper orthocones with theoretical distributions of chamber contents. All models are displayed with their proper static orientations A, Body chamber ratio (BCR) of 30% with an even distribution of liquid in the phragmocone (18.8% of all chamber volumes); B, BCR of 30% with an apical distribution of cameral deposits; C, BCR of 30% with a ventral distribution of cameral deposits; D, BCR of 30% with an oblique distribution of cameral deposits (ventral and apical direction); E, BCR of 25% with an even distribution of liquid in the phragmocone (32.7% of all chamber volumes); F, BCR of 25% with an apical distribution of cameral deposits; G, BCR of 25% with a ventral distribution of cameral deposits; and H, BCR of 25% with an oblique distribution of cameral deposits. Model components are color coded depending on their bulk density values (green, soft body; purple, cameral deposits; blue, cameral liquid; gray, shell). The tips of the blue and red cones denote the centers of buoyancy and mass, respectively. To improve visibility, arrows are located adjacent to the model. The blue and red arrows point toward the centers of buoyancy and mass. The green and purple arrows denote the end of the soft body and cameral deposits, respectively. All models are scaled with adult aperture diameters of 2 cm.

High-Taper Cyrtocones with Symmetrical Cameral Deposits.
High-taper, exogastric cyrtocones (curved and ventrally convex) with isometric conchs reconstructed from a fragmentary individual (AMNH-FI-145902; Supplementary Fig. S4) can produce a wider range of nonvertical orientations at lower BCRs (Fig. 7). A neutrally buoyant, 40% BCR conch without chamber contents has a static apertural orientation of ~−60° (i.e., tilted dorsally by 30° compared with a similarly proportioned orthocone; Table 1). Shortening the BCR to 30% allows ~7.7% of its phragmocone to be filled with cameral deposits and produces a condition that is closer to vertical (aperture orientation ~−77.7°; Table 1). At shorter BCRs, the aperture orientation is tilted ventrally from vertical, so that the venter is increasingly pointed upward (i.e., upside down; Table 1, Fig. 7).
Hydrostatic models of high-taper cyrtocones with symmetric cameral deposits. All models are displayed with their proper static orientations A, Body chamber ratio (BCR) of 40% (note there is no room for cameral deposits, which would produce negative buoyancy at this ratio); B, BCR of 30%; C, BCR of 25%; D, BCR of 22.365%; and E, BCR of 20%. F, Close-up view of medial section showing empty phragmocone; and G, close-up view of medial section showing symmetric cameral deposits in phragmocone. Model components are color coded depending on their bulk density values (green, soft body; purple, cameral deposits; gray, shell). The tips of the blue and red cones denote the centers of buoyancy and mass, respectively. To improve visibility, arrows are located adjacent to the model. The blue and red arrows point toward the centers of buoyancy and mass. The green and purple arrows denote the end of the soft body and cameral deposits, respectively. All models are scaled with adult aperture diameters of 2 cm.

High-Taper Cyrtocones with Theoretical Distributions of Chamber Contents.
Chamber contents were modified to explore a wide range of hydrostatic differences for the cyrtocone morphotype (i.e., even distributions of liquid; cameral deposits placed apically, or on the concave or convex side of the siphuncle; Fig. 8). Only BCRs of 30% and 25% were explored based on minimum estimates from incomplete specimens commonly exceeding these values (Supplementary Table S4). For models with a 30% BCR, the rather low percentage of cameral liquid (18.9%; Table 1) and cameral deposits in the phragmocone (~7.7%; Table 1) does not considerably influence orientation or stability, with all models assuming somewhat stable, downward-facing apertures with slight dorsal tilts (Fig. 8A–D). For models with BCRs of 25%, the percentage of the allowable cameral liquid and cameral deposits increases to 32.8% and ~13.3%, respectively (Table 1). Hydrostatic stability is slightly reduced in these models (Table 1). However, most models have nearly downward-facing apertures. The model with apically distributed deposits is an exception, with an aperture tilted ventrally by ~19.6° from vertical (Table 1). Note that results for these cyrtocones apply to cephalopods with both exogastric and endogastric (ventrally concave) curvature. However, for the latter, the apertural orientation is reversed about the vertical axis (i.e., venter-up orientations become dorsum-up).
Hydrostatic models of high-taper cyrtocones with theoretical distributions of chamber contents. All models are displayed with their proper static orientations A, Body chamber ratio (BCR) of 30% with an even distribution of liquid in the phragmocone (18.9% of all chamber volumes); B, BCR of 30% with an apical distribution of cameral deposits; C, BCR of 30% with a cameral deposits restricted to the convex side of the shell; D, BCR of 30% with cameral deposits restricted to the concave side of the shell; E, BCR of 25% with an even distribution of liquid in the phragmocone (32.8% of all chamber volumes); F, BCR of 25% with an apical distribution of cameral deposits; G, BCR of 25% with cameral deposits restricted to the convex side of the shell; and H, BCR of 25% with cameral deposits restricted to the concave side of the shell. Model components are color coded depending on their bulk density values (green, soft body; purple, cameral deposits; blue, cameral liquid; gray, shell). The tips of the blue and red cones denote the centers of buoyancy and mass, respectively. To improve visibility, arrows are located adjacent to the model. The blue and red arrows point toward the centers of buoyancy and mass. The green and purple arrows denote the end of the soft body and cameral deposits, respectively. All models are scaled with adult aperture diameters of 2 cm.

Lever Arm Ratio and Maneuverability
The ratio of two lever arms, (1) the distance from the source of jet thrust to the center of mass (D hm; Fig. 1B) and (2) the distance between the hydrostatic centers (D bm; Fig. 1B,C), is proportional to the ratio of the maneuvering torque to the restoring torque, given the same thrust potential between all compared cephalopods. Larger values represent higher maneuverability (defined here as the ease of changing orientation away from the static, equilibrium condition). These ratios are reported for all 43 hydrostatic models in Figure 9. The lever arm ratios for different morphologies belonging to earlier studies also appear in Figure 9 and Table 1. Cameral deposits generally reduce hydrostatic stability (and therefore D bm) by distributing the center of mass adapically. This change in mass distribution can also slightly increase D hm. Therefore, under the right circumstances, maneuverability can be increased through the addition of cameral deposits. However, this property heavily depends on BCR, with small differences (~5%) changing the lever arm ratio by an order of magnitude or more. When BCR approaches some value that satisfies a zero-stability condition, the lever arm ratio approaches infinity (note that these models in the current study only approach this condition and do not completely satisfy it; BCR of 24.3% for the low-taper models, and 22.365% for the high-taper models; Table 1, Fig. 9). Cephalopods with BCRs approaching 30% do not gain considerable benefits in maneuverability, with lever arm ratios falling below extant chambered nautilus. As BCR approaches ~25%, these cephalopods experience higher maneuverability, depending on cameral deposit distribution and conch morphology. The low-taper orthocones with 25% BCR have higher maneuverability than the chambered nautilus and, in some cases, higher than a highly maneuverable planispiral ammonoid (i.e., sphaerocone; Table 1, Fig. 9). The high-taper orthocone and cyrtocone models have comparatively lower lever arm ratios at 25% BCR. This difference is likely attributable to the difficulty in isolating variables. That is, while maintaining a linear BCR, this ratio by volume increases with conch expansion, thus decreasing the available cameral deposit mass under a neutrally buoyant condition.
Lever arm ratio (D hm/D bm) versus body chamber ratio (BCR) for each experimental condition in Figs. 2–8 (also see Table 1). D hm is measured from the source of jet thrust (the hyponome) to the center of mass location. D bm is the distance between the centers of buoyancy and mass. Therefore, increasing D hm (the maneuvering lever arm) and reducing D bm (the restoring lever arm) can improve maneuverability. BCR is measured as the body chamber length (from aperture to terminal suture), normalized by total length. A, Low-taper orthocones; B, high-taper orthocones; and C, high-taper cyrtocones. The gray shaded regions denote BCRs deemed less likely for cameral deposit–bearing cephalopods. That is, BCRs ≥ 40% would not have room for cameral deposits under a neutrally buoyant condition. BCRs < ~25% may produce unfavorable hydrostatics. The dashed horizontal lines denote the lever arm ratios for extant Nautilus (based on measurements from models in Peterman et al. [Reference Peterman, Barton and Yacobucci2019a]), and a theoretical sphaerocone ammonoid (based on measurements from models in Peterman and Ritterbush [Reference Peterman and Ritterbush2022b]). Cephalopods with lever arm ratios lower than that of Nautilus would not likely be able to modify orientation. Cephalopods with lever arm ratios approaching and exceeding the sphaerocone line could more easily change their orientations.

Discussion
Constraints on BCR
The influence of cameral deposits on the orientation, stability, and maneuverability of orthocone and cyrtocone cephalopods strongly depends upon BCR. Cameral deposit–bearing cephalopods are often partially preserved, obscuring our understanding of their conch proportions. Furthermore, the intraspecific, interspecific, and ontogenetic variability in these proportions is poorly understood. Hydrostatic analyses offer some constraints to these body proportions and therefore help constrain the swimming capabilities and life habits of these cephalopods. A ratio of ~40% body chamber length to total length (BCR) would produce a neutrally buoyant condition only with a completely empty phragmocone. Therefore, the BCR of cameral deposit–bearing cephalopods must be shorter than ~40% for cephalopods that must meet this condition. This proportion also agrees well with those of extant Nautilus. That is, if a Nautilus shell were unraveled into an orthocone, its BCR would be around 37% (see “Materials and Methods”). Negative buoyancy seems unlikely, but is within the realm of possibility for demersal cephalopods in shallow waters. However, those with more distal distributions above anoxic waters (Kröger et al. Reference Kröger, Servais and Zhang2009) would not have been able to support life in the water column if swimming was too costly. Many orthoceratoid fossils are incomplete. However, incomplete body chamber fossils can provide minimum estimates of body chamber length without preserved apertures, as long as the terminal septum is preserved. Assuming isometric proportions, minimum BCRs for cameral deposit–bearing orthocones commonly fall between one-quarter and one-third total length, with less-complete specimens saturating the dataset with even lower values (see Supplementary Table S4). Furthermore, Flower (Reference Flower1955a) reports roughly 6–14 adoralmost camerae lack cameral deposits in mature individuals. The low-taper models with lifelike distributions of cameral deposits (Fig. 2), have similar numbers of empty camerae. For BCR values of 30%, 25%, and 24.3%, the number of empty adoralmost camerae are 14, 11, and ~8.5, respectively (~24.6%, ~18.6%, and ~14.4% of all camerae, respectively). Note that the septal spacing of Smithorthoceras, the representative species for the low-taper models herein, is somewhat high (see Supplementary Table S1), suggesting that fewer empty camerae may be more analogous to the reports of Flower (Reference Flower1955a). Some cephalopods (e.g., Orthoceros regularis Schlotheim, Reference Schlotheim1820) are reported to have up to 32 empty camerae before cameral deposits are present, with only ~15 apical camerae containing deposits (Flower Reference Flower1964). This condition suggests that there is some degree of variability in cameral deposit distributions and proportions among orthocones. A phragmocone that has fewer chambers with deposits may suggest a longer BCR and/or deposits that more quickly taper throughout ontogeny (i.e., more closely resembling the current models with apically distributed cameral deposits; Fig. 3B,F). While these proportions do not necessarily narrow down the body chamber proportions to a specific value, they suggest that BCR values between 25% and 30% are plausible. Low BCR values of 20% are unlikely, based on incomplete body chambers measured from other studies commonly exceeding this value (Teichert and Kummel Reference Teichert and Kummel1960; Klug et al. Reference Klug, De Baets, Kröger, Bell, Korn and Payne2015; Pohle et al. Reference Pohle, Kröger, Warnock, King, Evans, Aubrechtova, Cichowolski, Fang and Klug2022; Supplementary Table S4). Furthermore, 20% BCRs produce upside-down orientations in orthocones and venter-up orientations in exogastric cyrtocones. These orientations seem rather awkward for the living animals. An upward-facing orthocone may find difficulty maintaining its attitude in the water column, because the hyponome would have to jet in the downward direction, which would be somewhat obstructed by the conch. Alternatively, this constraint could be alleviated if the animal was slightly positively buoyant or perfectly neutral. However, this condition contrasts with extant nautilid cephalopods, which find very slightly negative buoyancy easier to manage (Ward and Martin Reference Ward and Martin1978). If the cyrtocone was curved in the opposite direction (i.e., endogastric; concave venter), it would achieve a dorsum-up, horizontally facing soft body at this ratio. This orientation would be more analogous to modern cephalopods like squid and cuttlefish. While these conditions seem difficult to manage in most cases, it should be noted that some extant animals regularly assume upside-down orientations while swimming (e.g., upside-down catfish [Blake and Chan Reference Blake and Chan2007]; lionfish [Morris and Akins Reference Morris and Akins2009]; horseshoe crabs [Fisher Reference Fisher1975]; etc.). At any rate, based on estimates from measured fossils and the hydromechanical assumptions already noted, BCR values of between ~30% and ~25% are considered as the most plausible range in the current study.
Influence of Cameral Deposits on Static Orientation
Cameral deposits are often ventrally thicker, suggesting that they may have moved the center of mass ventrally, promoting more horizontal static orientations in these living cephalopods (Flower Reference Flower1955a,Reference Flowerb, Reference Flower1957, Reference Flower1964; Fischer and Teichert Reference Fischer and Teichert1969; Westermann Reference Westermann1977; Crick Reference Crick1988; Chamberlain Reference Chamberlain1993). Our results do not support this condition in most cases. Even when distributing all available cameral deposits on the ventral side of the siphuncle (Figs. 4C,G, 6C,G), or optimizing cameral deposit distributions to apically and ventrally move the center of mass (Figs. 4D,H, 6D,H), the cephalopods are only rotated by a few degrees (Table 1). Cameral deposits have a somewhat limited influence on the mass distribution due to their relatively small percentage of the total organismal mass (<16% in all cases, excluding the 20% BCR models; Table 1 and Supplementary Table S6), even if they have a higher density and smaller volume compared with the same mass of cameral liquid (factor of ~2.6 times; Hoffmann et al. Reference Hoffmann, Lemanis, Naglik, Klug, Klug, Korn, De Baets, Kruta and Mapes2015). A static horizontal orientation could occur under a few conditions: (1) when the centers of buoyancy and mass coincide (approaching BCRs of 24.3% and 22.365% in the low- and high-taper orthocones, respectively), and (2) when neutral buoyancy is violated, and larger masses of cameral deposits are placed in the phragmocone (potentially rendering these cephalopods immobile). These constraints were discussed in recent studies involving orthocone hydrostatics (Peterman et al. Reference Peterman, Barton and Yacobucci2019a,Reference Peterman, Ciampaglio, Shell and Yacobuccib); however, the current models add more context to their limits by exploring a broader range of BCRs and cameral deposit distributions. Nevertheless, these more recent approaches focused on hydromechanics do not explain why cameral deposits are often ventrally thicker. This distribution may relate to the morphogenesis of these deposits and the morphology of the cameral sheets within the phragmocone (for review, see Pohle et al. Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). However, this characteristic remains enigmatic and leaves open questions related to the adaptive value of such asymmetry.
Could Cameral Deposits Improve Maneuverability?
The term “maneuverability” can have many meanings (e.g., change in orientation per unit time, ability to change course while moving, turning radius, etc.). In the current study, we define maneuverability as the ability to modify orientation. Thus, maneuverability is the counterpart to stability, with the latter property imposing harsh constraints on how the living animal can deviate from its equilibrium condition, where the centers of buoyancy and mass are vertically aligned. Stability–maneuverability trade-offs are classic constraints encountered by swimming organisms (Weihs Reference Weihs1993, Reference Weihs2002; Fish Reference Fish2002; Peterman and Ritterbush Reference Peterman and Ritterbush2022a,Reference Peterman, Shell, Ciampaglio and Yacobuccib). Our analyses add important context to how some of the earliest swimming animals navigated these physical trade-offs. One interpretation of cameral deposits is that they offered a selective advantage by improving certain swimming abilities. That is, higher maneuverability could have improved the ability of these animals to capture prey items and escape predators by having fewer constraints on the range of orientations they could assume.
Orthocone and cyrtocone cephalopods without cameral deposits have much higher hydrostatic stability compared with their planispirally coiled counterparts (Table 1). However, these uncoiled morphotypes also generally have longer lever arms for maneuvering (i.e., the distance between the source of jet thrust, and the center of mass; Fig. 1B). The addition of cameral deposits has the potential to apically move the center of mass, which reduces the distance between the hydrostatic centers (the restoring lever arm), and increases the maneuvering lever arm (to a lesser extent). This configuration suggests that under the right circumstances, these uncoiled morphotypes can have greater mechanical advantage for modifying orientation, even if their static orientations are still nearly vertical. The lever arm ratio (D hm/D bm; Fig. 9) is used as a proxy for maneuverability herein. The modeled cephalopods demonstrate differences spanning several orders of magnitude, depending on BCR and cameral deposit distribution. The chambered nautilus can be used as a point of reference for this ratio, because its swimming behavior and related constraints are observable. Nautilus is considered a highly stable animal (Chamberlain Reference Chamberlain, Saunders and Landman1987; Peterman et al. Reference Peterman, Barton and Yacobucci2019a), capable of assuming a narrow range of orientations. Its rocking behavior during backward swimming demonstrates this limitation well (±10° or so; Chamberlain Reference Chamberlain, House and Senior1981). Therefore, a lever arm ratio below that of Nautilus (~17.5; Table 1, Fig. 9) could have produced even higher constraints on maneuverability. A sphaerocone ammonoid from an earlier study (Peterman and Ritterbush Reference Peterman and Ritterbush2022b) is much less stable and has fewer constraints on maneuverability (lever arm ratio of 45.9). Cephalopods with ratios approaching or exceeding this value would be able to easily modify their orientations.
Orthocones and cyrtocones with BCRs of 30% have lever arm ratios below those of Nautilus, regardless of how cameral deposits are distributed (Fig. 9). Cephalopods with these proportions could not considerably deviate from a stable, downward-facing posture. However, a small reduction in BCR to 25% considerably increases maneuverability. The most lifelike model in this study (a low-taper orthocone, resembling Smithorthoceras, with asymmetrical cameral deposits; Fig. 2) exceeds the maneuverability of a sphaerocone ammonoid at this BCR. The high-taper orthocones with 25% BCR and cameral deposits experience increases in maneuverability, with a lever arm ratio in between a Nautilus and sphaerocone. However, note that by maintaining this linear ratio, the soft-body ratio by volume increases with conch expansion, so comparisons with the low-taper models are not entirely analogous.
As BCR decreases in orthocone cephalopods (Fig. 9A,B), the hydrostatic centers eventually coincide (or nearly so), approaching a condition of no preferred orientation and infinite maneuverability. The computed BCRs of 24.3% and 22.365% for the low- and high-taper orthocones in the current study nearly meet this condition. Note that for models with symmetric cameral deposits, a small fraction of a percent decrease would create a spike in Figure 9, causing the lever arm ratio to approach infinity, and further decreases would invert the static orientation. Ventrally heavy deposits would prevent infinite maneuverability (and zero stability) from being reached, because the center of mass would be ventrally offset, but only at an extremely narrow range of BCRs. The cyrtocone conch, which is identical to the high-taper orthocone in all but curvature, does not experience comparable gains in maneuverability with the addition of cameral deposits. Even extremely skewed cameral deposit distributions (i.e., apical, or either side of the siphuncle) have little effect. These results suggest that cephalopods with slight shell curvature could have avoided conditions with extremely low stability across a range of BCR values. However, this relationship also raises the question of why cameral deposits appear at all in cyrtocone cephalopods.
Constraints on Life Habits and Paleoecology
Stability and maneuverability are highly sensitive to BCR, raising questions about the variability in life habits and swimming constraints among all orthocone cephalopods. If BCR fell below 30% in some species, these orthocones could have had higher maneuverability than planispiral cephalopods, potentially enabling a variety of orientations and jetting directions. Furthermore, body chamber proportions may have fluctuated during ontogeny as new septa were constructed and emptied during growth, suggesting hydrostatic changes within an individual could have been more extreme at BCRs around 25%. Some ectocochleate species display variability in body chamber length (Doguzhaeva and Mapes Reference Doguzhaeva, Mapes, Klug, Korn, De Baets, Kruta and Mapes2015; Kiselev Reference Kiselev2018), raising questions about the variation of BCR and the regulatory capabilities of cameral deposit–bearing cephalopods.
Orthocone cephalopods may have had the highest swimming speed and efficiency of any ectocochleate (when accounting for size and potential differences in propulsive musculature), but only in the apical direction (Peterman and Ritterbush Reference Peterman and Ritterbush2021). An orthocone with a short body chamber and high maneuverability would be able to modify its orientation before or during jetting. In this scenario, the cephalopod could travel in the apical direction in a non-vertical orientation without a strong restoring moment counteracting this posture (Peterman et al. Reference Peterman, Ciampaglio, Shell and Yacobucci2019b). This mode of locomotion may have had similarities to those of modern coleoid cephalopods; however, these orthocones would have lacked important control surfaces like mantle fins, which considerably aid in turning maneuvers and dynamic stabilization (Stewart et al. Reference Stewart, Bartol and Krueger2010; Bartol et al. Reference Bartol, Ganley, Tumminelli, Krueger and Thompson2022).
While orthocones with short body chambers would have had higher maneuverability, they would have faced the additional constraint of jetting in alignment with their hydrostatic centers to move in that direction (i.e., according to the thrust angle; Okamoto Reference Okamoto, Landman, Tanabe and Davis1996; Klug and Korn Reference Klug and Korn2004; Peterman et al. Reference Peterman, Shell, Ciampaglio and Yacobucci2020b). Misalignment of this thrust vector would cause more rotation and less translation for these hydrostatically unstable morphologies. Alternatively, orthocones with BCRs approaching and exceeding 30% would have been able to move while jetting in a broader swath of angles because less energy would be lost due to rocking. This stable scenario may have been more advantageous for demersal orthocones, because they could translate laterally above the seafloor with their senses fixed on the benthos. In contrast, unstable orthocones with BCRs approaching 25% could not assume a stable, downward-facing posture while moving laterally. Instead, these morphologies may have been better suited to catching swimming prey items by rotating their conchs in any direction. Likewise, this condition could have improved predator evasion by allowing the long axis of the conch to quickly travel in a broader range of directions. While diurnal migration would not be excluded for orthocones with short or long BCRs, a highly stable condition would be more ideal for more rapid vertical movements (Peterman and Ritterbush Reference Peterman and Ritterbush2021). On the other hand, high maneuverability might be advantageous for capturing prey in open-water conditions during comparatively slower vertical excursions. These trade-offs between hydrostatic stability, maneuverability, and thrust angle could be better explored in a physical setting with neutrally buoyant, untethered robots (like those of Peterman and Ritterbush [Reference Peterman and Ritterbush2022a]).
Preserved color patterns could add important context to orthocone swimming capabilities and possible soft-body proportions. The ability of some orthocones to assume non-vertical orientations is supported by some color patterns being restricted to the dorsal surface, suggesting countershading (Packard Reference Packard, Wiedmann and Kullman1988; Westermann Reference Westermann and Savazzi1998; Kröger et al. Reference Kröger, Servais and Zhang2009; Manda and Turek Reference Manda and Turek2015). This condition would only be possible if hydrostatic stability (and therefore BCR) was sufficiently low enough. While monochromatic and circumferential color patterns would be expected on conchs with a wide range of body chamber proportions, dorsally restricted patterns would be expected only on those with short BCRs (approaching 25%). Such comparisons would require exceptional specimens that not only preserve color patterns, but also enough of the conch to estimate accurate body proportions.
Finally, modifications of the conch could have modulated these physical constraints. That is, conch ornamentation and aperture shape vary within orthoceratoids, implying further nuances in functional capabilities for morphologies with similar hydrostatics. Adding annulations to an orthocone conch has been demonstrated to alter fluid wake while delaying the transition point between more turbulent wake structures as Reynolds number increases (Thakor et al. Reference Thakor, Seh, Gladson, Fernandez, Ivany, Green and Sun2023). Furthermore, ornamentation on ammonoid conchs has been demonstrated to considerably improve rocking attenuation by providing a dynamic stability mechanism (Peterman et al. Reference Peterman, Hebdon, Lusch, Byron, Panah and Ritterbush2025). The modeled specimens from the Buckhorn Asphalt have very fine transverse lirae; however, the hydromechanical influence of these structures is likely inconsequential (based on the fine ornamentation models of Peterman et al. [Reference Peterman, Hebdon, Lusch, Byron, Panah and Ritterbush2025]). Variability in aperture shape (e.g., rostra, lateral sinuses, and constrictions; Arkell et al. Reference Arkell, Furnish, Kummel, Miller, Moore, Schindewolf, Sylvester-Bradley and Wright1957; Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964) also implies potential differences in soft-body morphology and, therefore, life habits, sensing, and possibly propulsive potential. Some orthocone cephalopods had partially calcified structures interpreted as apertural lids or jaws (i.e., the aptychopsis; Holland et al. Reference Holland, Stridsberg and Bergström1978; Turek Reference Turek1978; Stridsberg Reference Stridsberg1984; Holland Reference Holland1987a,Reference Hollandb; Mironenko Reference Mironenko2021). Based on earlier hydrostatic analyses (Peterman et al. Reference Peterman, Hebdon, Ciampaglio, Yacobucci, Landman and Linn2020a), such features do not likely account for enough mass to considerably influence the center of mass location. However, these structures bear importance in understanding possible interactions between the prey and predators of these animals. Finally, the amount of soft body extending from the aperture has the potential to influence the distribution of mass to some degree. However, based on first-order principles, this influence would be rather low due to the similarities in densities between the soft body and seawater (~1.049 vs. ~1.025 g/cm3). That is, more soft body extending from the aperture would increase soft-body mass, but it would also increase the mass of water displaced, compensating for potential decreases in buoyancy. Similarly, both the centers of mass and buoyancy would move adorally in this scenario, preventing a disproportionate increase in stability. The differences in modeled soft-body proportions in earlier computer models demonstrate this relationship, yielding minor changes in hydrostatics with no soft-body extension (Peterman et al. Reference Peterman, Barton and Yacobucci2019a), conservative extension (Peterman and Ritterbush Reference Peterman and Ritterbush2021), and liberal extension (Peterman et al. Reference Peterman, Ciampaglio, Shell and Yacobucci2019b).
While our results are more directly relevant for cephalopods that are morphologically similar to Pseudorthoceratida, we would expect similar trends for other groups. That is, as BCR decreases, these cephalopods would experience a reduction in stability and a nonlinear increase in maneuverability within some narrow window. Cephalopods with larger siphuncles (i.e., Endocerida and Actinocerida) and more densely packed septa would devote more mass to these features, requiring a relative reduction in soft-body mass to maintain neutral buoyancy. We would expect that the trend between maneuverability and BCR would be slightly shifted for such cases. Endocerids, with mineralized endosiphuncular deposits, would have also had the additional constraint of distributing denser material within the siphuncle, a narrower region relative to the entire phragmocone. This constraint suggests it may have been more challenging for such deposits to act as counterweights. However, these relationships should be investigated with computer models and cross-validated with proportions preserved in fossil specimens. Additionally, these differences in internal morphologies may play important roles in the function of the buoyancy apparatus and metabolism of these living animals (Kröger Reference Kröger2003). These characteristics suggest there may have been differences in life habits and behavior between morphologically disparate groups that are not captured by hydromechanical analyses.
Orthocone cephalopods also display high variability in adult body size (Teichert and Kummel Reference Teichert and Kummel1960; Teichert et al. Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erben, Moore and Zeller1964; Holland Reference Holland1987b; Klug et al. Reference Klug, De Baets, Kröger, Bell, Korn and Payne2015, Reference Klug, Fuchs, Pohle, Korn, De Baets, Hoffmann, Ware and Ward2025; Pohle and Klug Reference Pohle and Klug2018). Hydrostatic properties like buoyancy, orientation, and hydrostatic stability index are perfectly scalable if body proportions stay the same throughout ontogeny. In contrast, hydrodynamic properties strongly depend on scale (i.e., Reynolds number; the ratio of inertial to viscous forces). Smaller body sizes and/or swimming speeds could operate in conditions where fluid viscosity begins to play important roles. Therefore, allometric growth, the hydrodynamic consequences of size, and the coupling of hydrostatics and hydrodynamics are important areas for future study.
Caveats and Alternate Hypotheses
While we highlight potential differences in swimming capabilities within orthocone and cyrtocone morphotypes, the adaptive value of cameral deposits in these animals remains enigmatic. There are several caveats that obscure relationships between these deposits and their function as internal counterweights (Flower Reference Flower1955a,Reference Flowerb, Reference Flower1957, Reference Flower1964; Fischer and Teichert Reference Fischer and Teichert1969; Westermann Reference Westermann1977; Crick Reference Crick1988; Chamberlain Reference Chamberlain1993). First and foremost, some cameral deposit–bearing orthocones have preserved body chambers estimated around 30% and higher (Supplementary Table S4), thus gaining no benefits in maneuverability (Fig. 9). Additionally, the modeled cyrtocone does not experience considerable increases in maneuverability across a broader range of BCRs, yet cameral deposits are still present. Asymmetrical, ventral-heavy deposits would maintain an unstable dorsum-up posture only for a very narrow range of BCRs, yet this asymmetry is present across morphologically disparate taxa, often with BCRs exceeding this functional range (Supplementary Table S4). Cameral deposits have a few other functional interpretations that cannot be discounted. The relative thickness of these deposits increases in the apical direction. While this distribution can be explained by apical chambers being older and having more time for deposits to form, it also would have reinforced the thinnest, and therefore weakest regions of the shell. An increase in mechanical strength could have been beneficial for resisting stress due to predation or hydrostatic pressure (the latter increasing with depth). Increases in mechanical strength are supported by a report of a sublethally injured psuedorthocerid (Seuss et al. Reference Seuss, Mapes, Klug and Nützel2012) and the lower relative abundance of empty phragmocone fragments preserved at some localities (Pohle et al. Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). Alternatively, cameral deposits have been suggested to potentially improve growth rates by sealing apical chambers and reducing the pumping energy required to prevent cameral liquid refill (Pohle et al. Reference Pohle, Hoffmann, Nützel, Seuss, Aubrechtová, Kröger, Stevens and Immenhauser2025). For cephalopods with larger body chambers, cameral deposits may have served as buoyancy-reducing ballasts (Crick Reference Crick1988) rather than counterweights, allowing larger conch sizes relative to the soft body. If natural selection favored either of these alternate possibilities, they would only indirectly influence hydromechanics at sufficiently low BCRs (around 25%).
Conclusions
A variety of theoretical morphologies with different soft-body proportions and cameral deposit distributions were used to explore a broad range of swimming constraints for orthocone and cyrtocone cephalopods. We introduce the dimensionless lever arm ratio (distance between the source of jet thrust and the center of mass to the distance between the hydrostatic centers), which is used as a proxy for maneuverability. We find that cameral deposits do not considerably influence the mass distribution with BCRs (body chamber length to total conch length) around or above 30%. Cephalopods with these proportions are highly stable while assuming a downward-facing posture. However, as BCR approaches 25%, a larger proportion of organismal mass can be dedicated to cameral deposits, reducing hydrostatic stability, improving maneuverability, and allowing a broader range of orientations. A stable condition is better suited for life above the seafloor, because such cephalopods could translate laterally with a downward-facing soft body. This condition is also favorable for faster and more efficient vertical movement (including diurnal migration in the water column). While all orthocones can accelerate more rapidly in the apical direction, the unstable yet maneuverable orthocones can more easily steer their shells in a broader range of directions while swimming and access prey by rotating the entire conch. These differences in swimming capabilities may reflect the high diversity, morphological disparity, and varied environmental settings occupied by these animals during the Paleozoic. In the context of systematics, the Orthoceratoidea may have displayed a larger degree of functional and/or physiological differences compared with some other orthocone clades. Finally, these swimming constraints are highly sensitive to BCR, highlighting the importance of knowing these proportions in cameral deposit–bearing cephalopods.
Acknowledgments
We thank A. Pohle (Ruhr-Universität Bochum) for discussions about orthoceratoid body chamber lengths and cameral deposit morphogenesis. We appreciate B. Hussaini and H. Ketchum (American Museum of Natural History) for their help with the measured specimens. We thank L. Fuelling for assistance sectioning and photographing the material. We thank T. Yancey (Texas A & M University, College Station, Texas) for donating the orthoceratoid specimens for this study to the AMNH. Finally, we appreciate the constructive and thoughtful comments of K. De Baets and P. Wagner during manuscript revision.
Competing Interests
The authors declare no competing interests.
Data Availability Statement
All 43 theoretical morphologies are housed in an online repository (Supplementary Dataset S1; https://doi.org/10.5281/zenodo.17829437). Models of each component used to compute hydrostatic properties are included for each experimental condition. This link also contains a PDF document with supplementary figures, tables, and text.











 Open access
Open access