



Introduction
Global chicken meat production was projected to increase from 105.6 million tonnes in 2020 to 181.3 million tonnes by 2050(Reference Alexandratos and Bruinsma1). According to the Food and Agriculture Organisation of the United Nations and Organization for Economic Co-operation and Development (OECD), chicken meat production already accounts for half of all global meat production, and it will be responsible for more than half (55%) of its growth from 2024 to 2033(2). Broiler chickens have the most efficient conversion rate of feed into meat for human consumption coupled with the smallest environmental footprint in terms of energy and water used across terrestrial food-producing animals, due in part to significant efficiency gains(Reference Havenstein, Ferket and Qureshi3–Reference Vaarst, Steenfeldt and Horsted5). The improvements in efficiency are largely attributed to genetics, but husbandry and nutrition have also contributed. Recent advances in broiler nutrition include refinements to ideal amino acid (AA) ratios(Reference Maharjan, Martinez and Weil6,Reference Wu7) , the increasing availability of supplemental (synthetic, crystalline) AA(Reference Selle, de Paula Dorigam and Lemme8), and improvements in exogenous feed enzymes such as phytase(Reference Selle, Macelline, Chrystal and Liu9), which have the potential to impact least-cost feed formulation practices by expanding the feed ingredient portfolios. In the development and use of broiler diets, the availability and cost of ingredients, chicken growth, and feed conversion ratios, remain major considerations. To meet anticipated global demand for chicken-meat in a sustainable manner; however, the formulation of broiler diets will need to advance beyond the present, simple ‘least-cost’ approach to embrace ecological factors (e.g., adding least environmental-costs by employing multi-objective optimization)(Reference Kleyn and Ciacciariello10,Reference Usigbe, Uyeh and Park11) .
The supply of feedstuffs and feed ingredients has a greater overall environmental impact than other stages of chicken-meat production(Reference Bengtsson and Seddon12–Reference Wiedemann, McGahan and Murphy14). Comparing different feed ingredients, the highest overall contribution to environmental impact is from soybean meal (SBM)(Reference Bengtsson and Seddon12,Reference Copley and Wiedemann13) , although the estimated magnitude of the SBM impact depends on how impacts are calculated and attributed(Reference Cappelaere, Le Cour Grandmaison, Martin and Lambert15), and on the origin of SBM and other feed ingredients(Reference de Boer, Van Krimpen, Blonk and Tyszler16). In many parts of the world, a reduction in SBM dietary inclusion has the potential to attenuate the environmental impact and carbon footprint of chicken-meat production(Reference Copley and Wiedemann13–Reference Cappelaere, Le Cour Grandmaison, Martin and Lambert15). This requires the use of alternative, locally available and hence context-specific feedstuffs and may result in trade-offs between the direct environmental benefits of use of alternative feedstuffs, and their indirect impact on sustainability through reduced crop yield or growth performance. Thus, further research into efficient alternatives continues to be needed, e.g. pursuing avenues as discussed in this review. Importantly, lower demand for SBM could help to limit the deforestation of subtropical rainforests in South America(Reference Gasparri, Grau and Angonese17,Reference Liu, Macelline, Chrystal and Selle18) . In recent years, the desire to reduce the reliance of poultry production on imported ingredients has been strengthened by the disruption of global trade such as during the COVID-19 pandemic and various geopolitical events that have affected the production or shipping of feed ingredients, leading to increased interest in the use of local agricultural byproducts in poultry diets(Reference Attia, Ashour and Nagadi19). Reduced SBM inclusions are common in reduced crude protein (rCP) diets in poultry(Reference Kandel, Liu and Toghyani20,Reference Siegert, Omotoso, Hofmann and Rodehutscord21) . Such diets may have additional advantages, such as attenuated nitrogen (N) and ammonia (NH3) emissions, and improved litter quality and chicken welfare.
A broad interpretation of sustainability requires that broiler production meets societal needs beyond availability, affordability and environmental sustainability, and includes welfare and health of chickens as well as public health. The welfare of broilers is developing into an increasingly important issue for supermarkets, consumers, legislators and producers(Reference Bessei22). Whilst traditional macro-environmental and sustainability factors (including eutrophication), water and energy efficiency, carbon footprint, aerial emissions, waste production, and antimicrobial resistance (AMR) keep increasing their importance, growing interest in in chicken welfare sharpens the focus on the in-house micro-environment provided by commercial grow-out farms. Reducing dietary crude protein (CP) concentrations whilst maintaining meeting amino acid requirements, is a promising nutritional pathway to alleviate both macro-and micro-environmental concerns(Reference Georgitzikis23). At the same time, concerns about the impact of AMR on humans, animals, and the global economy, have created a growing demand for reductions in the use of in-feed antimicrobials, both at supranational level (e.g., World Health Organisation, Food and Agriculture Organisation of the United Nations, World Organisation for Animal Health, and United Nations Environment Programme) and in individual countries. This may be desirable from a public health perspective but could potentially create a challenge to the maintenance of poultry ‘gut health’, referred to here as ‘the numerous complex and subtle interactions between the host mucosa, the intestinal content and all organisms residing in the intestinal tract’(Reference Ducatelle, Goossens, Eeckhaut and Van Immerseel24). Focus areas to enhance sustainable chicken-meat production include use of alternative feed ingredients, with consideration of protein and starch contents, and their interrelationship, increasing the use of supplemental AAs, strengthening of gut health through a balanced microbiome to improve performance and resistance to pathogens and environmental stressors and improved monitoring and management of the poultry house environment.
The purpose of this review is to describe opportunities in each of those areas, starting with alternative protein sources and rCP diets, improved understanding of the main constraints around digestion of starch and non-starch polysaccharides, and finishing with the interplay between diet and the internal (microbiota) and external (poultry house) environment of broilers. As each of these topics could be the subject of an entire review, our aim is to illustrate challenges and concepts using specific examples, and to provide a broad introduction to the depth of research that exists in these areas.
Replacement of SBM or reduction of protein content in broiler chicken diets
Replacement of SBM using vegetable sources
The identification of alternative, non-soybean protein sources, and overcoming the nutritional challenges associated with them, are complex issues that will require multi-factorial strategies(Reference Abdollahi and Ravindran25). Alternative protein feedstuffs include plant-based proteins, e.g., canola, grain legumes (notably chickpeas, faba beans, field peas, lentils and lupin), dried distiller grains or sunflower seeds, and non-vegetable proteins, such as insect meal and single cell protein (SCP) such as algal, yeast, fungal and bacterial protein. Selection of ingredients will be driven, in large part, by availability and cost in different parts of the world, which is beyond the scope of the current review. Here, we illustrate the challenges associated with alternative protein sources, as well as approaches to overcoming such challenges, using canola, pulses, and non-vegetable protein as examples.
Canola
Canola (Brassica napus) is a variety of rapeseed selected for low erucic acid and glucosinolates. Dietary protein derived from canola can be supplied in the form of whole ‘full-fat’ seed, partially defatted seeds in the form of cakes or meals, or protein isolates/concentrates and may partially replace dietary SBM when locally available such as in Europe or Australia(Reference Mulvenna, McCormack and Magowan26). Currently, inclusion of canola in broiler diets is usually restricted to less than 150 g/kg, which does not fully replace SBM as the principal protein source. This restriction has been attributed to lower available energy content and the presence of antinutritive factors (ANF) in canola(Reference Khajali and Slominski27). Canola meal has a N-corrected apparent metabolizable energy (AMEn) density of 8.37 MJ/kg in comparison to 9.37 MJ/kg for SBM(Reference Khajali and Slominski27). In addition to erucic acid and glucosinolates, canola meal may contain other relevant ANF including tannins and phytate (Table 1). The ‘as-is’ CP content of canola meal (365 g/kg) is less than SBM (456 g/kg). Differences in composition also exist at absolute AA level: canola meal contains more methionine (7.4 versus 6.5 g/kg) and cysteine (8.5 versus 6.7 g/kg) than SBM, but less arginine (20.4 versus 32.3 g/kg) and lysine (20.0 versus 28.6 g/kg)(Reference Khajali and Slominski27). Canola meal contains more neutral detergent fibre (260 versus 120 g/kg), acid detergent fibre (182 versus 75 g/kg) and total dietary fibre (317 versus 218 g/kg) than SBM, causing canola meal’s lower energy density(Reference Mejicanos, Sanjayan, Kim and Nyachoti28).
Key aspects of nutritional value and antinutritive factors of major current or potential plant-based protein sources for poultry diets (271–Reference Santiago, Teixeira and Calderano274)

Canola meal usage could be increased by breeding better canola varieties, improving canola oil extraction procedures, and formulating diets with appropriate enzyme inclusions to enhance canola digestion(Reference Kandel, Macelline and Toghyani29). For example, a yellow-seeded canola has been developed that has a 19.9% (241 versus 301 g/kg) lower total dietary fibre content than conventional, black-seeded canola varieties. The reduction in fibre content stemmed from larger seed sizes, relatively less hull fractions and lower lignin contents with associated polyphenols in the hulls(Reference Slominski, Jia and Rogiewicz30). In a comparison of black-versus yellow-seeded Brassica napus meals in broilers, apparent metabolizable energy (AMEn) of the yellow-seeded variety was significantly higher (9.54 versus 8.21 MJ/kg dry matter). Also, the mean apparent digestibility of total AAs was significantly higher (0.888 versus 0.834) for the yellow-seeded variety(Reference Jia, Mikulski and Rogiewicz31). Thus, canola with lower total fibre contents and providing higher AMEn in broilers can be developed and may contribute to the sustainability of chicken meat production. Protein digestibility may also be enhanced through means other than crop breeding. For example, in sorghum, RNAi silencing can improve protein digestibility through downregulation of genes that affect grain size and kafirin protein folding. Use of this transgenic sorghum allowed for a 20.5% reduction in SBM inclusion compared to commercial sorghum varieties, without compromising feed conversion efficiency(Reference Macelline, Godwin and Liu32). In any development of improved crops, quality needs to be balanced with yield, whereby varieties with high yields, disease resistance, and with high oil and protein contents are preferred(Reference Kandel, Macelline and Toghyani29).
In addition to improved genetics, improved processing may enhance the nutritional value of a crop. As an example of the role of processing, heat treatment (115°C for 25 minutes) of an expeller-pressed yellow canola meal reduced AMEn values and digestibility of some AAs(Reference Bryan, Abbott, Van Kessel and Classen33). Comparison of toasted and non-toasted canola meal also showed negative impacts of heat treatment, whereby the average ileal digestibility coefficient of 16 AAs was superior by 14.5% (0.875 versus 0.764) in non-toasted meals(Reference Newkirk, Classen, Scott and Edney34). Heat treatment reduces protein solubility and AA digestibility through numerous complex interactions including Maillard reactions(Reference Mauron35). These examples illustrate the importance of fine-tuning breeding and processing systems for canola meal production and other SBM alternatives in the quest to improve their value as a feedstuff for poultry.
Appropriate inclusions of feed enzymes in broiler diets containing SBM-alternatives such as canola provide a third pathway to enhance growth performance. There is more phytate in canola meal than SBM, both in absolute terms and relative to protein. Canola meal contained between 5.2 and 8.8 (an average of 6.69) g/kg phytate-P (76.4% of total P) as opposed to between 4.0 and 5.4 (average 4.53) g/kg phytate-P (68.3% of total P) in SBM in an Australian survey(Reference Selle, Walker and Bryden36). Phytate has the capacity to interfere with both protein digestion and intestinal uptakes of AAs, which are essentially consequences of de novo binary protein-phytate complex formation in the crop and gizzard(Reference Selle, Macelline, Chrystal and Liu9). The digestibility of AA in SBM can be improved through use of phytate, whereas only modest improvements were obtained in canola meal AA digestibility(Reference Ravindran, Cabahug, Ravindran and Bryden37). Olukosi et al. (Reference Olukosi, Kasprzak and Kightley38) and Moss et al. (Reference Moss, Chrystal and Dersjant-Li39) investigated inclusions of phytase and xylanase, individually and in combination, in broiler diets containing canola meal as the only protein source. Supplemented together, the enzymes increased average AA digestibility coefficients by 4.31% (0.799 versus 0.766) in the first and by 20.9% (0.832 versus 0.668) in the second study. Moreover, for all the AAs assessed, the responses in both studies appeared to be synergistic in comparison to the individual enzyme responses. One hypothesis is that the degradation of non-starch polysaccharides (NSP), and perhaps other fibre components, by xylanase is providing phytase with greater access to its substrate, facilitating the rapid degradation of phytate and amplifying the positive impacts of exogenous phytase on protein digestion and intestinal uptake of AAs. Other enzymes may also help to improve the utilisation of the fibre component of canola meal in broilers, as reviewed by Lannuzel et al. (Reference Lannuzel, Smith and Mary40). Dietary inclusions of multi-carbohydrase enzymes (e.g., pectolytic and hemicellulolytic enzymes), or combinations of enzymes with synergistic activities, were considered most promising.
In summary, as illustrated here for canola, a combination of breeding better varieties, improving processing systems and the judicious application of feed enzymes offer real potential to increase usage as a substitute for SBM. A similar approach, albeit with different actualisation and weighting of the three elements, could be applied to other plant-based alternative protein sources, with grain legumes or pulses providing additional examples as discussed next.
Pulses
Pulses are the dried edible seeds of leguminous crops that are also called grain legumes. The Food and Agriculture Organisation of the United Nations has distinguished eleven groups of pulses that belong to the family of Leguminosae, including beans (Phaseolus spp.); broad bean (Vicia faba: horse bean, broad bean and field bean); peas (Pisum spp.); chickpea (Cicer arietinum); cowpea (Vigna unguiculata); pigeon pea (Cajanus cajan); lentils (Lens culinaris); Bambara beans (Vigna subterranea); vetch (Vicia sativa); lupins (Lupinus spp.); soybean (Glycine max) and minor pulses such as jack bean, winged bean and yam bean. Pulses have N-fixing ability in their root nodules and require reduced fertiliser application, which contributes directly to their reduced carbon footprint, as fertiliser production is a significant contributor to emission of greenhouse gases(Reference Menegat, Ledo and Tirado41,Reference Virk, Lin, Kan and Sparks42) . Due to their high protein and energy contents and high digestibility, they are an excellent source of both for poultry. Like canola meal (see section Canola), locally grown grain legumes may partially replace dietary SBM to offset price volatility and supply-chain disruptions in some regions, depending on availability. However, the potential of pulses has not been effectively studied and their application in poultry diets has been limited due to the lack of data on digestible AAs and available energy content of pulses for poultry. Lack of data could drive lack of interest in using legumes as poultry feed ingredient, limiting its production, availability and the motivation to generate nutritionally relevant information, potentially creating a vicious cycle. Here, we briefly describe progress that has already been made in understanding the nutritional value of pulses in broiler chicken feeds.
Legumes make an attractive alternative to SBM because of the relatively high protein content of, e.g., chickpea (182–290 g/kg), faba bean (226–332 g/kg), field pea (215–307 g/kg), lentils (206–314 g/kg), and lupin (223–424 g/kg)(Reference Nalle43,Reference Urbano, Porres, Frías, Vidal-Valverde, Yadav, McNeil and Stevenson44) (Table 1). Additional advantages include the high digestibility of protein from chickpea and lupin seeds(Reference Nalle43) and the fact that the lysine content of beans is high (59.6–69.0 g/kg), similar to that of SBM and higher than in cereal grains, particularly maize grain(Reference Duranti and Gius45,46) . However, beans are deficient in tryptophan and sulphur-containing AAs like methionine (6.9–9.4 g/kg)(46,Reference Pol, Hristov, Zaman and Delano47) . Several legumes combine high protein content with high starch content, e.g. chickpeas (300–584 g/kg), faba beans (413–422 g/kg) and field peas (334–511 g/kg)(Reference Nalle43). In addition to similarities between legumes, there are differences between species. Chickpeas, for example, are richer in phosphorus and calcium than other pulses and faba beans are rich in lipids (10 g/kg), with a high proportion of linoleic and linolenic acids, whereas fat content in field peas is very low(Reference Nalle43,Reference Blair48) .
Legumes may contain a variety of ANF, however, most grain legumes grown currently have been selected for low levels of tannins and enzyme inhibitors(Reference Ravindran, Nalle, Molan and Ravindran49). Recent preliminary work found that trypsin inhibitor activity, phytate-P and phytic acid levels in raw chickpeas, faba beans, field peas, lentils, and lupins to be near or below that observed in solvent extracted SBM(Reference Hamungalu, Abdollahi and Morel50). This suggests that, at least in certain cultivars, plant breeding has already succeeded in overcoming many problems associated with ANFs in grain legumes. However, current lupin varieties still contain significant levels of alkaloids of which the quinolizidine alkaloids are the most relevant ANF, as well as trypsin inhibitor, phytate and flatulence-inducing oligosaccharides(Reference Helsper, Balkema-Boomstra and Ribot51). Thermal treatment during the pelleting process of poultry feed provides sufficient heat to inactivate most of the proteinaceous ANFs in lupins(Reference Sipsas, Mackintosh and Petterson52), illustrating that composition and processing of feed ingredients may need to be considered in tandem. Likewise, the effect of ANF in faba beans, such as protease inhibitors, tannins, and lectins, can be overcome through plant breeding and processing approaches, such as use of low tannin cultivars, dehulling(Reference Nalle, Ravindran and Ravindran53), and heat treatment in the case of proteinaceous ANF(Reference Hejdysz, Kaczmarek and Rutkowski54). The ANFs present in peas are haemagglutinins, trypsin inhibitors, chymotrypsin inhibitors, amylase inhibitors, tannins, oxalate, phenolic acid and phytic acid(Reference Savage55), while lentils contain phytic acid, protease inhibitors, tannins, oligosaccharides, and lectins(Reference Urbano, Porres, Frías, Vidal-Valverde, Yadav, McNeil and Stevenson44). Chickpeas are relatively free from ANFs(Reference Sipsas, Mackintosh and Petterson52), although they do contain some oligosaccharides, phytate, tannins and trypsin inhibitors(Reference Bampidis, Christodoulou and Nistor56). It can be surmised that various pulses are attractive but currently understudied and probably underutilized poultry feed ingredients and alternatives to the soya bean (Glycine max), which is itself a leguminous crop.
Replacement of protein using non-vegetable sources
While there are differences between countries, the main animal sourced proteins currently in use as feed ingredients in commercial poultry production include meat and bone meal, feather meal and fish meal(Reference Abun, Rusmana, Haetami and Widjastuti57–Reference Salehizadeh, Tajabadi Ebrahimi and Mousavi59). For example, the European Union (EU) currently allows the use of processed animal proteins of porcine origin in poultry feed, but not the use of meat and bone meal. Fish meal or fish waste has been evaluated as alternative protein source in poultry feed(Reference Macelline, Chrystal, Selle and Liu60,Reference Sydenham, Truong and Moss61) while poultry by-product meal is being considered as alternative protein source in fish feed(Reference Hasan, Rimoldi and Chiofalo62,Reference Marzouk, Gaber and Ahmad63) , reflecting sustainability and disease transmission concerns in different animal protein production sectors. Here, we briefly introduce alternative protein sources that have gained research momentum in recent years but that are not in widespread commercial use, including insects, worms, and SCP. Most of these are the subject of dedicated nutritional reviews, so our main aim here is to signpost the variety of alternative proteins under consideration.
Insects can be used whole or processed, whereby whole insects can be live, frozen or dehydrated and processed products may include full-fat insect meal, defatted insect meal(Reference Cullere, Tasoniero and Giaccone64,Reference Khan65) , or insect oil(Reference Somparn, Pootthachaya and Puangsap66). Insect protein is receiving considerable attention as a potential protein source for commercially produced poultry, in part because the protein content of insects (40–60%) is similar to that of SBM or fish meal(Reference Makkar, Tran, Heuzé and Ankers67). Evolutionarily, this is back to front as insects were part of poultry diets prior to large-scale production and use of SBM in poultry feed, and free-ranging and backyard chickens will voluntarily consume a wide variety of non-vegetable protein sources, including worms, insects and small amphibia, reptiles, birds, or mammals(Reference Selaledi, Mbajiorgu and Mabelebele68). The argument as presented here focuses on current commercial realities. There is also a perception that the use of insects would not create competition between food and feed, in contrast to the situation for SBM and fish meal(Reference Parlar and Ustundag69). Insect and worm species of interest for poultry nutrition include flies, mealworms, superworms, silkworms and other caterpillars, earth worms, termites, grasshoppers (various species) and locusts (Schistocerca gregaria)(Reference Cullere, Tasoniero and Giaccone64,Reference Khan65) (Table 2). As of 2021, eight insect species were authorised for use in poultry feed in the EU(Reference Meijer, Safitri, Tao and Hoek-Van den Hil70). Nutritional, environmental and commercial aspects of the production and use of insects in animal feed have been reviewed recently in a special issue edited by Veldkamp and Gasco(Reference Veldkamp and Gasco71). In addition to production effects, the use of insect products may have health benefits and improve litter quality, at least in experimental settings(Reference Ipema, Bokkers and Gerrits72). The consistency, volume and safety of worm or insect production on food waste or other materials need further consideration if it is to support broiler production at a commercial scale(Reference Parodi, Ipema and Van Zanten73,Reference van Huis and Oonincx74) .
Worms and insects under consideration as poultry feed ingredients. EU: insect species authorised for use in poultry feed in the European Union

In addition to plant-and animal proteins, microbial protein is of growing interests as a source of food or feed, with many species already in commercial production(Reference Ritala, Häkkinen, Toivari and Wiebe75). Microbes can be multicellular (e.g. fungi and filamentous algae such as spirulina-Limnospira spp.-), or unicellular (e.g., bacteria, yeasts and microalgae)(Reference Cheng and Kim76). Regardless of their structure, SCP is commonly used to refer to protein of microbial origin(Reference Ritala, Häkkinen, Toivari and Wiebe75). The idea of using SCP as poultry feed is not new: SCP for use in poultry production were already available in the 1960s(Reference Amara and El-Baky77). Since then, the global human population and commercial poultry production have grown dramatically, and one recent estimate is a market value for poultry feed of USD 1.7 billion or more by 2032 for yeast alone(Reference Amara and El-Baky77). Microbial protein or SCP is of interest because of its nutritional value, its contribution to waste recycling, and its efficiency. For example, bacterial protein for use in poultry, pig or fish feed can be produced using less land and less water than needed for SBM production, and bacteria can capture carbon dioxide (CO2) or methane in the process(Reference Salehizadeh, Tajabadi Ebrahimi and Mousavi59,Reference Woolley, Chaklader and Pilmer78) . Fungal protein can be grown on poultry litter, turning a waste product into a renewable resource(Reference Amara and El-Baky77). Algae and other SCP are of interest both as source of nutrition and as functional feedstuffs for broilers as reviewed recently by others(Reference Cheng and Kim76,Reference Abdel-Wareth, Williams and Salahuddin79) . When developing and evaluating new feed ingredients, the consumer must not be forgotten. Whilst the society is concerned about sustainable production, consumer drivers to maintain high chicken meat consumption such should also be kept in mind. Thus, consumers may have concerns about the diet of the poultry they consume, its appearance, and its flavour. For example, consumers in the EU did not necessarily like the idea of poultry being fed with insects(Reference Menozzi, Sogari and Mora80), the taste of insect-fed chicken meat(Reference Shaviklo81), or the appearance of the meat from spirulina-fed chicken(Reference Altmann, Anders, Risius and Mörlein82). Although these aspects are not addressed in our current review, the examples indicate the need to consider the education and preferences of consumers as part of the economic, environmental and social sustainability of poultry production and alternative feed ingredients.
Reduction of dietary protein
In addition to the replacement of SBM with alternative ingredient combinations that provide similar protein/AAs levels, a reduction in CP in broiler diets has the potential to diminish the chicken-meat industry dependence on imported SBM in many parts of the world. In rCP diets, SBM is not replaced with an alternative protein source, but with an alternative energy source, whereby the preferred alternative (e.g., wheat, barley, sorghum, maize, rice, potato) will depend on local availability. As a typical example, a CP reduction from 210 to 180 g/kg of CP in maize-soy diets decreased SBM inclusions by 28.7% (253 versus 355 g/kg) in Chrystal et al. (Reference Chrystal, Greenhalgh and McInerney83). In this study, dietary non-bound amino acid (NBAA) inclusions were elevated from 4.5 to 14.0 g/kg to meet targeted essential AA specifications, and maize inclusions were elevated from 543 to 654 g/kg, resulting in a 16% (362 versus 312 g/kg) increase in analysed dietary starch concentrations. However, dietary CP reductions of this magnitude (30 g/kg) often compromise growth performance, and the successful development of rCP diets is proving a real challenge. The specific feed grain used as the basis of rCP diets has a substantial bearing on the capacity of boiler chickens to accommodate dietary CP reductions, and in direct comparisons, maize(Reference Chrystal, Greenhalgh and McInerney83,Reference Greenhalgh, Chrystal and Lemme84) and sorghum(Reference Wang, Macelline and Toghyani85,Reference Macelline, Chrystal and Inanan86) were superior to wheat.
Numerous factors could be contributing to the shortcomings of wheat-based rCP diets(Reference Selle, Macelline, Chrystal and Liu87,Reference Selle, Macelline and Greenhalgh88) , where digestion rates seemed to be a main constraint (Table 3). The digestion of wheat starch is more rapid than the digestion of maize or sorghum starches, both in vitro (Reference Giuberti, Gallo, Cerioli and Masoero89) and in vivo (Reference Selle, Moss and Khoddami90). Starch content is higher in rCP than in standard diets, thus, resulting in a higher amount of glucose delivered by the former. Glucose stimulates the secretion of the anabolic hormone insulin(Reference Chang, Chiang and Saltiel91). In chickens offered diets with standard CP levels (ca 200 g/kg), maize-based diets supported 27.4% (12.1 versus 9.5 mIU/l) higher plasma insulin concentrations than wheat-based diets at 42 days post-hatch(Reference Kulcsár, Mátis and Molnár92). Moreover, Petrilla et al. (Reference Petrilla, Mátis and Kulcsár93) compared standard (220 g/kg) to rCP (188 g/kg) diets based on maize or wheat. At 42-days post-hatch, plasma insulin concentrations were numerically elevated by 16.7% (6.081 versus 5.209 ng/ml) following the dietary CP reduction with maize but were depressed by 15.9% (4.785 versus 5.688 ng/ml) in chickens offered wheat-based diets. In this study, chickens fed maize-based rCP diets performed better than those fed wheat-based rCP diets. The role of insulin in regulating glucose metabolism in broilers remains elusive(Reference Gomez-Capilla and Langslow94) and a recent review of the impact of feed grains on insulin and growth performance was described as ‘inherently speculative’(Reference Selle, Macelline, Wang and Liu95). From in vitro data(Reference Ebsim96), the starch digestion rate of field peas is slower than for either maize or wheat (Table 4). On average, field peas contain 236 g/kg CP and 413 g/kg starch(Reference David, Nalle, Abdollahi and Ravindran97), although both parameters are variable. The inclusion of field peas into wheat-based rCP diets could prove advantageous because their additions would retard overall dietary starch digestion rates. The latter, in turn, may enhance broiler growth performance(Reference Herwig, Abbott, Schwean-Lardner and Classen98), possibly through slower intestinal uptakes of glucose and more sustained insulin responses(Reference Weurding, Enting and Verstegen99).
Digestible starch fractions and starch digestion rates of three unprocessed feed grains with crude protein and starch concentrations expressed on a dry matter basis

a Starch composition categorised by digestion rate into rapid, slow or resistant (adapted from Giuberti et al. (Reference Giuberti, Gallo, Cerioli and Masoero89)).
Proportion (%) of starch digested in four feedstuffs in an in vitro model simulating small intestinal digestion. Adapted from Ebsim(Reference Ebsim96)

In addition to starch digestion rates, protein content differs between feed grains. In one Australian survey, the average protein content of wheat samples was 115.5 (ranging from 88.0 to 162.0) g/kg as opposed to 101.9 (ranging from 71.0 and 118.0) g/kg in sorghum and only 80.0 (ranging from 69.0 to 91.0) g/kg in maize(Reference Bryden, Li and Ravindran100). Paradoxically, the higher protein content of wheat compared to maize and sorghum demands elevated NBAA inclusions in wheat-based rCP diets (15.3 g/kg of NBAA on average in maize-based rCP diets versus 25.3 g/kg in wheat-based rCP diets). This is because higher SBM proportions are replaced with wheat than with the feed grains that have lower protein content. In addition, AAs digestibility is lower in wheat protein than in the SBM protein it replaces(Reference Bryden, Li and Ravindran100), and it is unlikely that non-bound and protein-bound AAs are completely bioequivalent(Reference Selle, Macelline, Chrystal and Liu101,Reference Baker102) .
Given that intestinal uptakes of NBAA are more rapid than uptakes of intact protein(Reference Wu103), excess NBAA in the diet may result in post-enteral AAs imbalances and the catabolism of potential AAs surpluses(Reference Brosnan104). The catabolism of AAs generates NH3, which is a toxic metabolite affecting avian more than mammalian species(Reference Stern and Mozdziak105,Reference Wilson, Muhrer and Bloomfield106) . It follows that inadequate NH3 detoxification, or high levels of circulating NH3, may depress broiler growth performance(Reference Aguihe, Ospina-Rojas and Sakamoto107–Reference Ospina-Rojas, Murakami and Moreira110). Collectively, glutamine, glutamate, glycine, and serine are crucial to NH3 detoxification and uric acid elimination. The subject of NH3 overload or inadequate NH3 detoxification in wheat-based diets has been covered in more detail by Selle et al. (Reference Selle, Macelline, Chrystal and Liu87).
The importance of glycine and serine (or glycine equivalents) concentrations in rCP diets has been previously demonstrated(Reference Dean, Bidner and Southern111). Siegert and Rodehutscord(Reference Siegert and Rodehutscord112) proposed that dietary glycine equivalent requirements range from 11 to 20 g/kg for young broilers. Recently, glutamine and asparagine were shown to facilitate high dietary NBAA inclusions without compromising growth performance or N accretion, which was attributed to enhanced acid–base homeostasis(Reference Ibrahim, Kenéz, Rodehutscord and Siegert113). Ideal AA ratios (IAAR) are a cornerstone of least-cost formulations of broiler diets in which AA requirements are expressed relative to lysine. The IAAR for rCP diets may differ from standard-CP diets with non-essential AAs playing a potential role. In addition, specific IAAR for wheat-based diets should be developed to minimise risks. These should include requirements for glycine, serine, glutamine, and glutamate (as distinct from the collective glutamic acid). In addition, proline, serine, and glutamic acid may need to be given consideration because they are abundant in feather protein(Reference Leme, Sakomura and Vargas114), and partitioning of AAs for feathering may be given priority in poultry(Reference Wylie, Robertson and Hocking115).
L-carnitine is protective against NH3 toxicity(Reference Kloiber, Banjac and Drewes116). In chickens offered 160 g/kg CP, sorghum-based diets, 75 mg/kg L-carnitine significantly improved weight gain (15.0%), feed intake (8.34%) and feed conversion ratio (FCR) (5.82%) from 7 to 33 days post-hatch. In contrast, L-carnitine responses were not observed in 190 and 220 g/kg CP diets(Reference Greenhalgh, Hamilton and Macelline117). These outcomes are consistent with L-carnitine counteracting NH3 overload in chickens offered 160 g/kg CP diets supplemented with 51.0 g/kg NBAA. L-carnitine is both present in feedstuffs and may be synthesised endogenously from its precursors, lysine and methionine(Reference Rebouche118). L-carnitine promotes fat mobilisation via the so-called ‘carnitine shuttle’, transferring long-chain fatty acids across the barrier of inner mitochondria membranes to undergo β-oxidation and fat is partitioned to energy production(Reference Eder119). In addition to benefits in preventing NH3 overload, L-carnitine has displayed promise in limiting lipid deposition in some feeding studies as the addition of 50 mg/kg L-carnitine to 180 g/kg CP diets reduced relative abdominal fat-pad weights by 20.5%(Reference Rabie, Szilágyi and Gippert120). Although L-carnitine can mitigate some of the risks of wheat-based diets, its use alone is not sufficient to meet the limitations of wheat in rCP diets.
Blends of wheat and other grains such as barley, maize, rice, or sorghum, depending on local availability, may offer a promising alternative to rCP diets containing wheat as the only grain. Diets formulated to 215, 190 and 165 g/kg CP based on an equal blend of wheat and maize were offered to straight-run chickens from 1 to 41 days post-hatch(Reference Maynard, Ghane and Chrystal121). Following the CP reduction from 215 to 165 g/kg, weight gain and feed intake was maintained. This contrasted with results obtained for the wheat-based diet. When using the blend, FCR was compromised by 3.43% (1.597 vs 1.544), but SBM consumption was reduced by 63% (500 vs 1339 g/chicken). Compared with wheat-only diets, an equal wheat and maize blend is probably advantaged by a lower protein content and decreased NBAA inclusions, coupled with slower starch digestion rates. Where maize is not a practical option, sorghum may offer a viable alternative.
As dietary CP decreases, the ratio between dietary starch and dietary protein increases. Capping that increase may be beneficial in wheat-or maize-based rCP diets. In wheat-based diets, limiting starch:protein ratios in 197.5 g/kg CP significantly increased weight gain by 10.4% and numerically improved FCR by 4.04% from 7 to 35 days post-hatch(Reference Greenhalgh, Chrystal and Lemme84,Reference Greenhalgh, Chrystal, Selle and Liu122) . In maize-based diets, capping ratios in 175 g/kg CP diets improved weight gain by 3.45% and FCR by 3.75% from 7 to 35 days post-hatch. The overall impacts of capping ratios at three dietary CP levels were significant for weight gain (P = 0.004) and FCR (P < 0.001)(Reference Greenhalgh, Chrystal and Lemme84,Reference Greenhalgh, Chrystal, Selle and Liu122) . In both studies, the capping of starch:protein ratios was achieved by partially substituting SBM (505 g/kg CP) with ‘full-fat’ soy (362 g/kg CP). A similar effect may be achievable through using canola meal.
Improvement of carbohydrate use in broiler chicken diets
Importance of particle size in starch digestibility
Growth of broilers, which is primarily skeletal muscle protein synthesis, is dependent on the balanced provision of dietary AAs and energy(Reference Moughan123). For the accretion of 1 g of body protein, 5.35 kJ energy input is required(Reference Aoyagi, Tasaki, Okumura and Muramatsu124–Reference Selle and Liu126). The bulk of this energy is provided by glucose, which is derived from dietary starch as the major component in feed grains. In general, starch shows high digestibility rates in chickens associated with a significant variability. This variability, in turn, may cause lack of synchronisation with protein digestion decreased efficiency of use of the energy for growth. Starch digestion is influenced by the chemical properties of the starch itself (e.g., degree of branching) and the physical and chemical properties of the grains, the latter being an aspect rarely considered in the context of protein digestion. There is abundant evidence that both particle size and chemical composition of grains have major effects on the rate of starch digestion in vitro (Reference Al-Rabadi, Gilbert and Gidley127) translating to in vivo consequences on feed efficiency which have been well established in pigs(Reference Al-Rabadi, Hosking and Torley128) and to a certain extent in chickens(Reference Amerah, Lentle and Ravindran129,Reference Amerah, Ravindran, Lentle and Thomas130) . In addition, particle size distributions and structural features of undigested feed in the small intestine of chickens have been investigated(Reference Yang, Flanagan and Selle131).
When grains are hammer-milled, disc-milled, or ground in the chicken gizzard(Reference Amerah, Lentle and Ravindran129), a wide range of different particle sizes are produced, with the overall starch digestibility representing a weighted average of digestion across the particle size distribution. Particle size has a major effect on the rate of starch digestion(Reference Heaton, Marcus, Emmett and Bolton132). In vitro digestion of milled grain fractions ranging from 1.1 mm to 0.16 mm in size follow classical 1st order reaction kinetics(Reference Al-Rabadi, Gilbert and Gidley127) so they can be linearised by plotting the natural logarithm of (1-starch fraction digested) against time. The slope of these lines gives the 1st order rate coefficient, a fundamental characteristic of the digestion process. Good correlations between in vitro and in vivo rates of starch digestion in broilers have been reported(Reference Weurding, Veldman and Veen133). The most likely explanation for the marked size dependence of starch digestion rate for milled grains is that α-amylase is impeded from passing into and through the grain particles(Reference Al-Rabadi, Gilbert and Gidley127). In other words, the digestion rate of starch is influenced by enzyme diffusion to the starch substrate. If valid, this leads to a prediction that the rate coefficients should scale inversely with the surface area (related to particle size squared) of the particles. This is indeed what is found for a range of milled grains examined to date. Furthermore, the slope of the inverse relationship between rate coefficient and particle size squared can be related to an apparent diffusion coefficient that represents the intrinsic feature of a particular grain that controls digestibility at a given particle size(Reference Ratanpaul, Williams, Black and Gidley134). Reducing the proportion of larger particles in a milled grain sample will lead to faster starch digestion. How feed grain particle size affects passage rate, gut motility or gut microbiota is largely unknown, and could be explored in tandem with its effect on digestion.
The most likely reason for the apparent barrier to amylase diffusion within milled grains is the presence of intact cell walls within particles(Reference Grundy, Edwards and Mackie135). Milled grains contain intact cell walls surrounding starch in pieces of endosperm tissue as well as fragments of other (non-starch containing) grain tissues such as aleurone and bran layers. Endosperm cells are typically of the order of 0.1 mm in size, considerably smaller than the particle sizes of milled grains, so cells containing starch with intact cell walls are abundant in larger particle sizes and therefore provide a major barrier to digestive enzyme action(Reference Grundy, Moughan and Wilde136). For this reason, heat treatment and/or cell wall acting enzymes can be used in feed formulation to provide (in theory) improved access of digestive enzymes to starch and increase energy delivery to the chicken. Experimental demonstration of the ability of cell wall acting enzymes to break wheat, barley and maize endosperm cells has been reported, providing demonstrated benefits on chicken performance(Reference Liu, Flanagan, Roura and Gidley137). To provide predictive tools to modify digestion characteristics, further work is needed to identify what influence grain particle size and apparent amylase diffusion properties have on starch digestion in chickens. A major challenge in this type of study in chickens is the location of particle size reduction. Unlike the mouth and stomach in pigs and humans, the gizzard in poultry is likely to have a major effect on particle size before starch digestion is accelerated in the small intestine(Reference Amerah, Ravindran, Lentle and Thomas130). More studies on the effect of grain type and particle size distribution on size breakdown in the gizzard(Reference Amerah, Lentle and Ravindran129) are needed to identify how to influence subsequent starch digestion through manipulation of feed.
It is worth noting that a similar effect of particle size (at a shorter length scale) is also seen for some isolated starch granules(Reference Dhital, Shrestha and Gidley138). For maize starch granules (and all other common cereals except for high amylose variants), there are ‘pinhole’ channels leading from the outside to the interior of the granule resulting in very limited particle size dependence on digestion rate. However, potato starch has no such channels and shows a strong size dependence that scales with the particle size squared(Reference Dhital, Shrestha and Gidley138). The ‘pinhole’ channels are a characteristic of cereal starches(Reference Huber and BeMiller139), but most other starch sources are expected to show granule size dependence. Given that cereals provide starch in broiler diets, granule size effects are unlikely to be important in poultry nutrition.
Starch digestion and starch:protein dynamics
In chickens, digestion of starch and absorption of glucose is rapid, especially in maize-based diets(Reference Moran140). Starch is enzymatically converted into glucose by the combined action of pancreatic alpha-amylase in the gut lumen and gut wall amyloglucosidases(Reference Moran140). Subsequent intestinal uptake of glucose occurs primarily via the Na+-dependent transporter, SGLT-1(Reference Chen, Tuo and Dong141). The reliance of both glucose and AA transport on co-absorption with sodium may create competition for sodium and could contribute to the apparent reduction in AA digestibility coefficients observed with high dietary starch concentrations(Reference Moss, Sydenham and Khoddami142). Once in the bloodstream, plasma glucose concentrations are relatively high in broilers but tightly controlled. For example, chickens offered diets based on maize, wheat or rice had mean plasma glucose concentrations of 12.72 mmol/L for up to 420 minutes post-prandially, with a range of 11.10 to 14.52 mmol/L(Reference Li, Li and Liu143). The relationship between glucose and insulin in poultry differs from that of mammals, as broilers have naturally high levels of plasma glucose despite the presence of endogenous insulin circulating at seemingly normal concentrations(Reference Dupont, Tesseraud and Simon144). The conversion of glucose to lactic acid in the intestinal mucosa may be important in maintaining glucose homeostasis in poultry(Reference Riesenfeld, Geva and Hurwitz145).
Average dietary inclusions of 535 g/kg semi-purified starch in starter, grower, and finisher broiler diets as slowly digestible pea starch and/or rapidly digestible wheat starch were assessed by Herwig et al. (Reference Herwig, Abbott, Schwean-Lardner and Classen98). The balance of 25% slowly and 75% rapidly digestible starch maximised feed efficiency and linearly improved breast meat yield. These researchers suggested that gradual glucose absorption from slowly digestible starch may better match the physiological energy requirements of broilers by allowing the immediate utilisation of glucose for muscle deposition instead of energy storage.
The relevance of starch:protein disappearance rate ratios was investigated in Sydenham et al. (Reference Sydenham, Truong and Moss61) based on fishmeal and corn starch inclusions in sorghum-SBM broiler diets. Proximal jejunal starch:protein disappearance rate ratios were quadratically related to weight gain (r = 0.849; P < 0.001) and FCR (r = 0.838; P < 0.001), from 15 to 28 d post-hatch. It may be deduced that a disappearance rate ratio of 3.59:1 would generate the maximum weight gain of 1265 g/bird and a similar ratio of 3.88:1 would generate the minimum FCR of 1.287. The impact of protein sources on starch:protein disappearance rates in wheat-based broiler diets has also been investigated(Reference Macelline, Chrystal, Selle and Liu60). Lower ratios were obtained with mixed protein sources than with a single protein source and resulted in improved FCR. These examples show that starch and protein digestive dynamics should be considered in tandem and support the concept that the provision of slowly digestible starch coupled with rapidly digestible protein in broiler diets is beneficial. A better understanding of starch and protein digestive dynamics including heat processing will assist the chicken-meat industry in reaching more sustainable production.
Non-starch polysaccharides
Dietary fibre or NSP provide the structural framework to feed grains. The capacity of NSP leached from grain cell walls to increase digesta viscosity in chickens offered barley-and wheat-based diets is established; consequently, NSP-degrading enzymes are routinely added to appropriate diets(Reference Chesson146). The inclusion of NSP-degrading enzymes in broiler diets has been extensively researched(Reference Kim, Morgan and Moss147) and reviewed(Reference Choct148) and is not within the ambit of the present review. Development of transgenic crops may be another avenue to offering potential solutions to some of these long-known problems(Reference Macelline, Godwin and Liu32). NSP also plays a positive role in broiler nutrition. Together with resistant starch, that is, starch that is not digested along the small intestine due to a combination of enzyme digestion and passage rate effects (Reference Dhital, Warren and Butterworth149,Reference Svihus and Itani150) , NSP are available for fermentation by hindgut (predominantly the caeca in chickens) microbiota. This results in the conversion of polysaccharides through monosaccharides such as glucose to short chain fatty acids (SCFA), primarily acetate, propionate, and butyrate, that are available for uptake by the chicken intestinal epithelia. SCFA can provide energy to the chicken (in principle 40% of that of glucose) but are also important modulators of the immune system and support hind-gut development. However, it has been estimated that the chicken only gets 3-5% of its energy from SCFA(Reference Svihus, Choct and Classen151), so caecal fermentation may be relatively limited. This may be because only residual feed particles smaller than about 200 µm can enter the caeca(Reference Vanderghinste, Bautil and Bedford152). It will therefore be important to determine (a) the proportion of NSP at the ileum that is < 200 µm in size, and (b) the opportunity to control caecal fermentation through using finely milled fibre particles.
The use of carbohydrate, rather than protein, as major energy source for the gut microbiota has several benefits for the chicken. The production of SCFA reduces caecal pH, which supports tissue development, promotes a healthy immune system, and provides some protection against opportunistic pathogens(Reference Ali, Ma and La153). In the absence of fermentable carbohydrate, the microbiota use protein as an energy source, resulting in a negligible pH reduction arising from the production of potentially toxic alkaline metabolites from microbial transformations of some AAs. The benefit of availability of cell wall fibres/polysaccharides for fermentation is accompanied by risks, notably the potential for absorbing water and promoting passage without complete fermentation. This is particularly acute for soluble polysaccharides that can result in unwanted excretion of water(Reference Choct, Hughes and Wang154). One consequence of the widespread use of cell wall-degrading enzymes in broiler rations could be to reduce water holding in the caeca and large intestine by depolymerising cell wall polysaccharides. Although this principle is established, there may be opportunity for optimisation through tailoring enzyme selection and levels for specific feed components and formulations.
In the current environment of reduced use of antimicrobials, there is an opportunity to tailor chicken diets to nourish a gut microbiota that maintains chicken health and reduces levels of not only avian pathogens such as strains of E. coli, Enterococcus cecorum and Enterococcus faecalis but also food safety risk organisms such as Campylobacter and Salmonella species (see subsequent section). In the future, the choice of grain source for chicken diets should consider not only the starch and protein digestion dynamics but also the effects of residual undigested fractions on the gut microbiota in the caeca and colon. Several tools exist to potentially manipulate these features including choice of ingredient, extent of grinding and particle size distribution, addition of enzymes, and interactions with other feed ingredients (Table 5).
Features of carbohydrate nutrition in broilers and opportunities for improvement

Nutrition and microbes
Beneficial microbes: gut microbiota
Recent advances in sequencing methodologies and the rapid growth of biomarker databases opened a new horizon in our understanding of the complex microbial communities that shape and are shaped by the organs they inhabit. This sparked major interest in this field, which in recent years has been dominated by 16S amplicon sequencing for the analysis of bacterial communities(Reference Bajagai, Trotter and Williams155). The abundance of the data collected via 16S amplicon sequencing reveals the importance of the gut microbiota in various processes and functions critical for the host’s health and well-being. Although this varies across gut sections, the gut microbiota is generally very diverse, even when accounting for possible overestimation resulting from the 16S methodology limitations(Reference Bajagai, Trotter and Williams155). This diverse community brings a diverse metagenome (collection of all genes from the microbiota), and a range of functions and processes that occur in the gut, such as nutrient absorption and vitamin production, metabolising dietary compounds that the host cannot digest, and helping develop the immune system. The microbiota also includes pathogens (see section Harmful microbes: human and animal pathogens), thus, host health depends on the balance between beneficial and pathogenic microbes and their functional capabilities(Reference Pickard, Zeng, Caruso and Núñez156).
As a very complex and still largely unpredictable system, the intestinal microbiota composition is not only shaped by the feed and its nutritional composition alone; initial inoculum, the environmental microbiome, bird characteristics, and production system (open range, barn, cage) all have a significant impact on the presence and abundance of specific microbial groups (Reference Feye, Baxter and Tellez-Isaias157,Reference Ricke and Rothrock158) . Even birds from the same parent genetic stock grown in the same facility on the same batch of feed can have unrelated microbial communities dominated by different phyla(Reference Stanley, Geier and Hughes159). In addition, each intestinal section has a unique and distinguishable microbiome due to differences in the role, chemistry, oxygen availability and remaining feed composition in that gut section. On the other hand, there is high reproducibility in the composition of microbiota that naturally colonise and utilise certain raw feed ingredients such as grains, soy, meat-and-bone meal or fish meal(Reference Haberecht, Bajagai and Moore160), and different bacterial genera play recognizably distinct roles in the gut microbiome. For example, fibrolytic genera, including Bifidobacterium, Lactobacillus, Faecalibacterium, and Ruminococcus, as well as Prevotella and Anaerostipes, play a role in fibre fermentation (Reference Cronin, Joyce, O’Toole and O’Connor161,Reference Erkmen162) . These genera are commonly found in grains and seeds(Reference Haberecht, Bajagai and Moore160). Among the proteolytic bacteria, the most prolific genera are Alcaligenes, Bacillus, Micrococcus, Staphylococcus, Clostridium, Pseudomonas, Flavobacterium, and Brevibacterium (Reference Erkmen162). Some bacteria, such as Akkermansia muciniphila, are known for their involvement in both lipid breakdown and the regulation of fatty acid synthesis(Reference Rodrigues, Elias-Oliveira and Pereira163). High-throughput microplate assays measuring the rate of utilisation of common poultry feed ingredients by major intestinal bacterial species could be helpful in optimising feed, but such simplified in vitro systems might not capture the complexity of the in vivo system, which includes cross-feeding between bacterial species as well as strain-level differences within bacterial species. Both phenomena are critical for species survival, influencing their physiology, pathogenesis, feed preferences and host interactions (Reference Nie, Utter and Kerns164,Reference Smith, Shorten and Altermann165) . Bacterial responses to feed ingredients may not be reproduced in feed mixtures, further complicating the idea of precise control of nutrient-microbiome interactions, although there is growing interest in AA-microbiome interactions (see section Aminobiotics).
Unlike other agricultural animals, commercially produced poultry are generally fully separated from the maternal transfer of microbiota and are hatched in commercial hatcheries that operate under high biosecurity regulations to remove all significant poultry pathogens from the microbiota-naïve hatchlings. While this practice is effective in preventing the spread of infectious diseases, it also opens a range of gut health issues as natural colonisation is no longer an option. In the absence of maternal transmission, the chicken gut gets colonised by the environmental microbiota including from human handlers(Reference Stanley, Geier and Hughes159). In addition, poultry feed carries a microbial community that remains viable regardless of feed processing and storage(Reference Li, Meng and Wang166). As hatchlings transition from yolk sacs to solid feed, the gut microbiota undergoes dramatic changes in response to dietary macro-and micro-nutrients, and the first feed offered to hatchlings combined with the first bacterial species that enter the gut will play a key role in shaping the new microbial community.
The hatchery, transport and poultry house environments also significantly impact gut microbiota colonisation processes(Reference Pottenger, Watts and Wedley167). Factors such as litter quality(Reference Wang, Lilburn and Yu168), poultry house and transport(Reference U. Bello, Idrus and Yong Meng169) biosecurity measures, previous outbreak history, biofilms, and rodent control all(Reference Wickramasuriya, Park and Lee170) contribute to the initial inoculation of the gut. This type of colonisation is opportunistic and would not be reproducible even if we used the same batch of hatchlings on the same farm multiple times due to extreme spatial and temporal fluctuations in this environment(Reference Stanley, Geier and Hughes159). Major fluctuations of the hatchling microbiota often stabilise during the first week, depending on the environmental microbial load, diet, and many other variables. However, the maturation of the community continues throughout the production cycle, whereby certain bacterial groups slowly become dominant and new ones emerge while others decline(Reference Li, Meng and Wang166). This process is cut short in broilers, but major weekly microbiota fluctuations have been reported(Reference Donaldson, Stanley, Hughes and Moore171).While it is impossible to completely prevent colonisation by pathogens from the environment, farmers often use probiotics very early to shift the balance between pathogenic and beneficial microbiota. These probiotics are often mixes of multiple species and are comprised of poultry-specific strains(Reference Krysiak, Konkol and Korczyński172). Another approach used is a predesigned poultry caecal microbiota mix to mimic maternal microbiota(Reference Kayal, Stanley and Radovanovic173). Early gut colonisation in chickens is a research frontier with many mechanisms and treatments yet to be discovered.
Future research into precision probiotic and prebiotic supplementation and controlled gut colonisation will extend from productivity and health-oriented studies towards optimisation of nutrient utilisation to improve sustainability of production, and decreasing greenhouse gas emissions, prevalence of antimicrobial resistance genes, or decreased litter moisture (see section Associations between broiler nutrition and the internal and external environment). In addition, advances in the underexplored realm of intestinal fungal, archaeal, and viral communities (Reference Borrel, Brugère and Gribaldo174,Reference Salahi and Abd El-Ghany175) , and the increasing availability and affordability of metagenomic and metabolomic data are likely to bring a second wave of development in gut health understanding.
Harmful microbes: human and animal pathogens
While many members of the gut microbiota contribute to digestion, gut health and performance (see section Beneficial microbes: gut microbiota), other members may cause disease in broiler chickens or chicken meat consumers (Table 6). Sustainable nutrition would ideally include prevention of animal and human disease, and evaluation of alternative feed ingredients or feed processing methods should consider their impact on pathogen loads. Here, we summarize examples and knowledge gaps around the relationship between feed ingredients or processing methods and animal and/or human pathogens. In doing so, we illustrate the importance of an integrated evaluation of performance and health impacts of current and alternative feed ingredients.
Reported associations between feed ingredients or processing methods and health outcomes in broiler chickens or consumers associated with specific groups or species of pathogens. BCO = bacterial chondronecrosis with osteomyelitis (including femoral head necrosis), D = decrease, I = increase, S = ingredient is a potential source, nd = no data

Table 6. Long description
The table has 6 columns and 17 rows including the header row. The columns are labeled as Nutrition and health aspects, Campylobacter spp., Clostridium perfringens, Eimeria spp., Enterobacteriaceae, Enterococcus cecorum, and Salmonella spp. The rows are grouped under Ingredients, Amino acids, Starches, Antimicrobials, and Processing methods. Each cell contains data indicating whether a specific ingredient or processing method increases (I), decreases (D), or is a potential source (S) of a pathogen, or if no data (nd) is available. The table also includes a legend explaining the symbols used. Row 1: Ingredients, Protein source, Animal, nd, I (186, 196), nd, nd, nd, S (176). Row 2: Insect, S (179), I, S (179, 180), nd, S (179), nd, S (179). Row 3: Soybean meal, nd, D (186), nd, nd, nd, S (176). Row 4: Corn gluten, nd, D (186), nd, nd, nd, nd. Row 5: Potato, nd, I (186), nd, nd, nd, nd. Row 6: Canola meal, I (177, 178), nd, nd, nd, nd, D, S (176, 178, 183). Row 7: Amino acids, Arginine, I (194), D (190), D (192), nd, nd, D (193). Row 8: Glycine, nd, I (186, 188), nd, nd, nd, nd. Row 9: Starches, Wheat, nd, I (195, 196), nd, nd, nd, nd. Row 10: Maize, nd, nd, nd, I (199), nd, nd. Row 11: Sorghum, D (206), D (207), nd, D (199), nd, nd. Row 12: Antimicrobials, nd, nd, D (209), nd, nd, D (210). Row 13: Processing methods, Fermentation, nd, nd, nd, nd, nd, D (183). Row 14: Particle size (small), nd, nd, I (204), nd, nd, D (184). Row 15: Exogeneous enzymes use, nd, nd, nd, D (195, 200), nd, nd. Row 16: Health impact, Broiler chickens, Disputed, Necrotic enteritis, Coccidiosis, BCO (due to E. coli) (201), BCO (279), Serotype dependent. Row 17: Consumers, Food poisoning, None, None, Antimicrobial resistance genes (203), Extremely rare (280), Food poisoning.
Dietary protein sources can affect animal health directly by acting as a source of pathogens, or indirectly, through its impact on the gut microbiome or host immune system. Protein rich feed ingredients of animal and plant origin, including SBM and alternative plant-based protein sources, notably canola meal, may serve as a source of Salmonella (Reference Reiter, Adler and Kolar176). When free of pathogens, some of those ingredients may have beneficial effects on the intestinal pathogen load in broilers. For example, whilst canola meal can be a source of Salmonella, its use has been associated with reduced Salmonella levels in other studies (Table 6). To complicate the situation further, multiple microbial outcomes need to be considered when evaluating the impact of alternative dietary protein sources since the use of canola meal reduced Salmonella levels but increased Campylobacter colonisation of the broiler gut(Reference Visscher, Abd El-Wahab and Ahmed177,Reference Yadav, Teng and Choi178) . Insect larvae have their own gut microbiome and pathogen profiles, which depend on their feed sources and rearing conditions. For example, black soldier fly larvae may harbor viable Campylobacter, Clostridium, Enterobacteriaceae, and Salmonella. Microbiological risk assessments for use of insect larvae should consider primary and opportunistic pathogens as well as bacteria carrying AMR determinants(Reference Raimondi, Spampinato and Macavei179). The super worm Zophobas morio has been associated with increased levels of C. perfringens (Reference Benzertiha, Kierończyk and Rawski180). Conversely, the meal worm Tenebrio molitor was not associated with direct increase in C. perfringens (Reference Benzertiha, Kierończyk and Rawski180). Its use, however, led to the loss of beneficial gut microbes (Bacteroides), which is a potential risk factor for C. perfringens infection(Reference Biasato, Ferrocino and Biasibetti181), demonstrating again that both direct and indirect effects of alternative feed ingredients on the gut microbiome need to be considered. Supplementation of lactobacillus-fermented rapeseed meal in broiler diets reduced Campylobacter jejuni cecal colonization(Reference Khattak, Galgano and Pedersen182). In addition to the type and hygienic status of ingredients, the preparation of alternative protein sources, e.g., fermentation of canola meal or particle size of wheat bran, may influence colonisation and infection in broilers, as shown for Salmonella (Reference Ashayerizadeh, Dastar and Shams Shargh183,Reference Vermeulen, Verspreet and Courtin184) .
High dietary protein levels and poor dietary protein digestibility stimulate hindgut protein fermentation by the gut microbiome and overgrowth of C. perfringens (Reference Lee and Rochell185). This effect is not uniform across protein sources. Diets containing animal proteins (e.g., meat and bone meal, feather meal, and fish meal) or potato protein concentrate led to higher intestinal C. perfringens counts than diets with the same protein content derived from SBM, pea protein concentrates or corn gluten(Reference Wilkie, Van Kessel and White186). The impact of individual ingredients may be minimised by the composition of the complete diet. For example, whilst fish meal can be associated with an increase in C. perfringens, reduced CP diets with fish meal as protein source led to lower levels of C. perfringens. A similar reduction was not observed in diets based on SBM, which is already associated with low C. perfringens loads(Reference Drew, Syed and Goldade187). The impact of different protein sources may be mediated, in part, by their specific AA (see also section Aminobiotics). Animal source proteins are higher in glycine than plant-derived proteins, and potato-derived protein is higher in glycine than other plant-based proteins. Glycine stimulates Clostridium growth in vitro and is associated with ileal Clostridium levels and the severity of the disease it causes, necrotic enteritis (NE)(Reference Wilkie, Van Kessel and White186,Reference Dahiya, Hoehler, Van Kessel and Drew188) . Thus, high glycine levels may jeopardize broiler health. Again, the impact of the feed ingredient may depend on diet composition, as the addition of glycine to low protein diets may be beneficial for performance and bone strength(Reference Yuan, Karimi and Zornes189). Reduced CP diets have been associated with higher levels of circulating arginine in plasma(Reference Chrystal, Greenhalgh and McInerney83) and arginine supplementation reduces the severity of experimentally induced NE(Reference Zhang, Gan and Shahid190). The effect of arginine on protecting the gut mucosa and alleviating immunosuppression seemed to extend beyond NE into infectious bursal disease, coccidiosis, and a challenge with Salmonella enterica Serovar Typhimurium(Reference Aguzey, Gao and Haohao191–Reference Zhang, Li and Shahid193). In contrast, Campylobacter has been suggested to benefit from intestinal arginine, using it as a nutrient(Reference Awad, Smorodchenko and Hess194). Thus, it would be prudent to assess the impact of dietary arginine or other amino acids across the wider range of pathogen species.
Among the cereals used in poultry feeds, wheat is the most notorious in relation to pathogen load. As described in the ingredient-focussed sections, high wheat feeds have been associated with increases on digesta viscosity, prolonged transit time, and delayed digestion and absorption of nutrients. In addition, it alters the gut microbiome in favour of pathogens, including C. perfringens (Reference Raza, Bashir and Tabassum195,Reference Shojadoost, Vince and Prescott196) . If those traits are specific to wheat, it stands to reason that diets based on other sources of starch, e.g., those based on maize, have comparatively lower C. perfringens levels and lower risk of NE. Nutrition trials, such as comparisons of wheat-and maize-based diets generally assess outcomes in terms of performance parameters(Reference Chrystal, Greenhalgh and McInerney83), without considering pathogen loads. As methods for targeted (e.g., animal or human pathogen focussed) and untargeted (gut microbiome) detection of bacteria are increasingly available and affordable, it would be opportune to use nutrition trials to evaluate both production and microbial outcomes. Examples of such integrated studies include analyses of alternative energy sources (almond hulls, and distillers dried grains) that specifically considered the impact of diet on Clostridium levels to show the absence of adverse microbial consequences(Reference Loar, Moritz, Donaldson and Corzo197,Reference Wang, Singh, Kong and Kim198) . Cereal type (barley, rye, corn, and sorghum) and exogenous enzyme administration can also modulate populations of potentially pathogenic Enterobacteriaceae in the poultry caecum(Reference Ciurescu, Vasilachi, Idriceanu and Dumitru199,Reference Józefiak, Rutkowski and Kaczmarek200) . This includes E. coli, a potential contributor to femoral head necrosis in broilers(Reference Wideman201), and Klebsiella spp., which are of concern as ‘drug resistance gene traffickers’ between poultry and humans(Reference Wyres and Holt202,Reference Aguilar-Bultet, Bagutti and Egli203) . As for protein, both the source of carbohydrates and processing methods need to be considered. For example, increasing inclusion levels of coarse corn have been associated with decreased Clostridium levels(Reference Singh, Ravindran and Wester204). Sorghum, while initially problematic as a broiler feed ingredient due to anti-nutritional factors (e.g., condensed tannins) and protein resistance to digestion (e.g., kafirin matrix), continues to be of interest(Reference Selle, Cadogan, Li and Bryden205) due to potential anti-clostridial and anti-Campylobacter activities(Reference Hamad, Gerges and Mehany206,Reference Shields, Gang and Jordan207) . Genetic markers of antimicrobial activity have been identified in sorghum and their presence does not impact negatively on production traits, creating an opportunity to selectively breed for this trait in crops intended for use in animal feed(Reference Shields, Gang and Jordan207).
In a brief history of poultry nutrition over the last hundred years, Elwinger et al. (Reference Elwinger, Fisher and Jeroch208) concluded that ‘poultry nutrition is now a scientifically mature subject’. They postulate that changes in industrial practice will slow down. The imperative to develop broiler diets that are economically, environmentally, and societally sustainable suggests the opposite. A significant threat to broiler health, coccidiosis, has long been managed through inclusion of antimicrobial agents, notably coccidiostats, in poultry diets. Because of concerns about selection for antimicrobial resistance (AMR), there is pressure to control the disease through other means, even if the feasibility and value of such approaches is debated(Reference Kadykalo, Roberts and Thompson209). Reduced use of antimicrobials, whilst potentially beneficial for public health, has already contributed to the emergence of Enterococcus cecorum as a pathogen of concern in broiler production(Reference Schreier, Karasova and Crhanova210) and development of nutritional interventions to reduce this risk is in its infancy(Reference Hankel, Bodmann and Todte211). The growing availability of DNA-based technologies provides a unique opportunity to integrate the study of broiler nutrition, broiler health and public health. This integration enables us to detect and mitigate negative microbial impacts of dietary modifications whilst positive impacts can be identified and exploited. To date, such studies have largely focused on C. perfringens, as major cause of disease in broilers, and non-typhoidal Salmonella serotypes as major cause of disease in humans. Many gaps in our knowledge exist in relation to other pathogens, including those that have hitherto been controlled with antimicrobials (Table 6). Whilst it would be naïve to think that nutrition alone can solve the problem of known and (re-)emerging infectious diseases, its role in promoting or mitigating pathogen load should be considered more routinely so that, at the very least, we can abide by the ‘do not harm principle’ when introducing or scaling up the use of new feed ingredients. This concept could be extended beyond microorganisms found in the gut, as described here, and include the impact of nutrition on litter quality and its consequences for other health conditions such as footpad dermatitis (see section Associations between broiler nutrition and the internal and external environment).
Aminobiotics
Reducing the dependence on SBM in developing broiler chicken sustainable diets requires the use of alternative/local protein sources and crystalline AA supplementation (see 2.3). As highlighted in sections Beneficial microbes: gut microbiota and Harmful microbes: human and animal pathogens, the microbiota is responsive to changes in feed ingredients including AAs. Bacteria, like eukaryotic cells, use AA as building blocks of protein synthesis or as energy source(Reference Morowitz, Carlisle and Alverdy212). However, to date, little is known about essential AA requirements of commensal or pathogenic bacteria and potential implications of dietary AAs on the development of the chicken gut microbiota. The selective nourishment of the gut microbiota with AA may provide a novel prebiotic approach, also called ‘aminobiotics’(Reference Beaumont, Roura and Lambert213). Some of the main AA-fermenting bacteria in the small intestine are species belonging to the genus Clostridium, Bacillus, Lactobacillus, or Streptococcus, and the phylum Proteobacteria. These bacterial populations seem to utilise preferentially Lys, Met and Phe. In contrast, AA-fermenting species in the colon and caeca (e.g., Clostridium and Proteobacteria) seem to respond also to Arg, Gly and the branched-chain amino acids in addition to Lys(Reference Davila, Blachier and Gotteland214–Reference Dai, Wu and Zhu216).
Microbial AA preferences may exist at genus and species level(Reference Yang, Pollard and Höfler217). For example, members of the genus Pseudomonas consume Ala, Glu, and/or Asp but not sulphur AA or Thr. Uniquely, Pseudomonas aeruginosa also uses Lys while other species in the genus cannot(Reference Sivolodskii218). Salmonella enterica requires Asp, Glu, Gly, Pro, Ser, Ala, Arg, and Asn as both carbon and nitrogen sources(Reference Kwan, Plagenz and Cowles219), while Pasteurella multocida requires sulphur AA, Glu and Leu for optimal growth(Reference Jablonski, Jaworski and Hovde220). Under anaerobic conditions, Ser, Asp, Cys and Asn are the sole carbon source for E. coli and they act as strong chemo-attractants for this species. In contrast, E. coli growth is inhibited by Val and Leu(Reference Yang, Pollard and Höfler217). Although there are conflicting preferences in this summary, such as for Leu which is required P. multocida whilst it inhibits E. coli, these findings suggest that certain dietary AA patterns have the potential to selectively suppress the growth of opportunistic pathogens in the intestine and to help prevent enteric bacterial diseases(Reference Beaumont, Roura and Lambert213). In addition, selection of commensal bacteria of known AA requirements to seed the gut early in the life of the chick might enable formulation of dietary AA patterns that specifically meet the requirements of both the chicken and its healthy microbiota.
The fact that the chicken microbiota is responsive to free AAs has several potential implications relevant to poultry nutrition: firstly, it offers a new dietary tool to influence gut health through selectively feeding the commensal microbiota while inhibiting pathogens; secondly, the findings imply that the microbiota itself has a selective appetite for some non-bound dietary AAs and may potentially compete with the host for limiting AAs. The latter opens a question about the potential impact of the gut microbiota on AA requirements in chickens. Moreover, different individuals develop different microbiota profiles, which may impact the efficiency of use of dietary AA and ultimately the requirements of the chicken. For example, Niknafs et al. (Reference Niknafs, Fortes and Cho221) showed that marginal AA deficiencies were associated with slow or fast growth. This study did not include characterisation of the microbiota so the nature of its relationship with deficiencies, if any, is unknown. Another significant gap in the application of the aminobiotic concept is the understanding if or how free dietary AAs may impact the microbiota before being absorbed in the proximal small intestine, and whether encapsulation might be required to create an in vivo aminobiotic effect. While this new concept has currently more questions than answers, the potential for future applications in poultry nutrition justify additional research.
Associations between broiler nutrition and the internal and external environment
The relationship between nutrition, sustainability and the environment is multifaceted and complex, both within and beyond the poultry house (Figure 1). The most direct connection is via ingestion of litter, which makes up about 5% of the total feed intake of broilers(Reference Malone, Chaloupka and Saylor222), creating multidirectional links between nutritional inputs, gastrointestinal microbial populations and litter conditions(Reference Wang, Lilburn and Yu168,Reference Bindari and Gerber223,Reference Collett224) . After introduction of broiler chicks to grow-out farms, bacteria ingested with the litter, like bacteria from feed and other elements of the environment (see section Beneficial microbes: gut microbiota), contribute to the establishment of the gut microbiome(Reference Plumblee Lawrence, Cudnik and Oladeinde225,Reference Cressman, Yu and Nelson226) . Subsequent feedback loops resulting from the recycling of excreta showed that ingesta can be beneficial, as seen in the development of protective immunity to Eimeria species and the optimisation of amino acid supplementation(Reference Williams, Johnson and Andrews227,Reference Hussein, Khattak and Vervelde228) . Alternatively, it can contribute to accumulation of avian or human pathogens. Despite the importance of pathogens for chicken and human health, there is limited understanding of the exact contribution of litter and litter conditions to pathogen loads, in part because pathogen loads(Reference Plumblee Lawrence, Cudnik and Oladeinde225) and litter condition are difficult to define(229–Reference van Harn, Dijkslag and van Krimpen235).
Choice of feed ingredients (top left) affects the environmental footprint of poultry production, as well as the quantity of solids, water and pathogens excreted by broiler chickens into the litter (centre). This, in turn, impacts the birds’ health and productivity through ingestion of litter-associated microbiome (bottom left) and moisture-related conditions such as footpad dermatitis (bottom right), whilst also affecting the environment within and beyond the shed, e.g., through gaseous emissions and odour (top right), and environmental release of nutrients and microorganisms (right).

A major contributor to litter quality is water. Fast-growing broilers contributed a large amount of water to the litter, with volumes estimated at 0.5 to 3.2 L/m2/day and a cumulative total of 104 L/m² during the grow-out periodDunlop, Blackall and Stuetz(Reference Dunlop, Blackall and Stuetz236). The daily amount of water added to the litter varies over the grow-out cycle due to variations in water and feed intake, feed quality, metabolic water production, chicken health and liveweight density. In addition, it varies spatially within the house. Improvement in broiler rearing over the past decade have occurred in terms of feed formulations (e.g. precision formulation and a reduced energy content), drinker management, stocking rates, and genetics. These have contributed to more efficient feed and water utilisation and a shorter grow-out period. Collectively, this has likely reduced the amount of water being applied to the litter by approximately 25% from the values previously estimated(229). Even so, in a typical modern broiler house with a floor area of 2500–3000 m², the amount of water added to the litter during the grow-out period was estimated to be 1000 to 7,200 L/day. This water needs to be removed by evaporation every day mostly through ventilation(Reference Pepper and Dunlop237). If the water additions are not removed daily, the litter moisture content increases resulting in ‘wet litter’, which has been defined as a litter having a moisture content greater than 25%(Reference Collett224). There is compelling evidence from both research and field experience of a proportional relationship between increasing litter moisture content and detrimental impacts on performance, pathogen loads, chicken health and welfare (e.g., footpad lesions and dermatitis), odour and ammonia(Reference Collett224,Reference van Harn, Dijkslag and van Krimpen235,Reference Dunlop, Moss and Groves238–Reference Brink, Janssens and Demeyer244) . When wet litter cannot be avoided with conventional management, growers may need to apply additional practices that accelerate water evaporation and removal from the poultry house such as by tilling the litter and increasing heat and air movement at the surface with high-powered air circulation fan(Reference Pepper and Dunlop237,Reference Mou, Czarick and Fairchild245,Reference Mou246) . Feed formulation and nutritional strategies, combined with management of drinkers(Reference Malone, Marsh Johnson and Owen242) and limited pathogen exposure, can help to reduce water excretion and prevention of such situations.
One strategy to decrease water intake in absolute terms and relative to feed intake is a reduction in CP(Reference Brink, Janssens and Demeyer244,Reference Alfonso-Avila, Cirot, Lambert and Létourneau-Montminy247,Reference Chrystal, Moss and Khoddami248,Reference Lemme, Hiller and Klahsen249) , adding to the potential benefits of rCP as discussed elsewhere in this review. Decreasing CP intake by 2–3 percentage points numerically reduces water intake by 8–23%(Reference Greenhalgh, Chrystal, Selle and Liu122,Reference Brink, Janssens and Demeyer244,Reference Alfonso-Avila, Cirot, Lambert and Létourneau-Montminy247,Reference Chrystal, Moss and Khoddami248,Reference Santonja, Georgitzikis and Scalet250) . On average, this translates into less water in excreta and litter(Reference Collett224,Reference van Harn, Dijkslag and van Krimpen235,Reference Alfonso-Avila, Cirot, Lambert and Létourneau-Montminy247,Reference Lemme, Hiller and Klahsen249,Reference Hejdysz, Bogucka and Ziółkowska251,Reference Jeon, Hong and Kang252) , although not all research shows a direct benefit in terms of drier litter during pen trials(Reference Brink, Janssens and Demeyer244,Reference Ferguson, Gates and Taraba253) . Some research studies reported litter moisture contents of 40–60%(Reference van Harn, Dijkslag and van Krimpen235,Reference Sonnabend, Spieß and Reckels254–Reference Son, Lee and Kim258) , which is not representative of typical litter conditions in commercial production, limiting the applicability of the findings. Other approaches to precision nutrition and improved gut health, such as those discussed in this review, may also reduce water excretion and hence decreases the reliance on ventilation to evaporate water and maintain dry and friable litter conditions. This improves the sustainability of chicken meat production both through improving broiler welfare and by reducing the energy footprint of electricity use.
Environmental benefits of precision nutrition may extend beyond the broiler house, as outlined in the introduction for inputs into broiler production (e.g., environmental impact of cultivation and transportation of feed ingredients) and illustrated below for unintended outputs from broiler production. For example, rCP diets decrease excretion and litter content of N. Some of the N excreted into the litter consists of uric acid, which is a precursor of atmospheric NH3 in broiler grow-out facilities(Reference Brink, Janssens and Demeyer244,Reference Groot Koerkamp259) . Reducing CP by 15 to 25 g/kg has been shown to lower total N in excreta or litter by 8 to 19%(Reference Santonja, Georgitzikis and Scalet250,Reference Ferguson, Gates and Taraba253) and NH3-N in litter by 13 to 22% in broiler breeders(Reference Brink, Janssens and Demeyer244,Reference van Emous, Winkel and Aarnink260) , which corresponded to 9 to 30% reductions in NH3 emission from the litter(Reference Brink, Janssens and Demeyer244,Reference Santonja, Georgitzikis and Scalet250,Reference van Emous, Winkel and Aarnink260) . Benefits to the natural environment of reducing NH3 emissions and the export of N in spent litter include reducing the risks of acidification, eutrophication, reduced biodiversity, and formation of particulates that may be inhaled by stockpersons(Reference van Emous, Winkel and Aarnink260,Reference Ogino, Oishi, Setoguchi and Osada261) . Nitrogen reduction is also required to meet increasingly stringent environmental regulations in some countries(Reference Lemme, Hiller and Klahsen249).
While reducing excreted N has some reported environmental benefits, it does not seem to contribute to a meaningful reduction in the emissions of nitrous oxide (N2O)(Reference de Rauglaudre, Méda and Fontaine262,Reference Wiedemann, Phillips and Naylor263) , which is considered to be an important greenhouse gas with a global warming potential that is 265 greater than CO2(Reference Myhre, Shindell, Bréon, Stocker, Qin, Plattner, Tignor, Allen, Boschung, Nauels, Xia, Bex and Midgley264). It has been postulated that the weak connection between N excretion and N2O emissions is due to the necessity for specific conditions that contribute to the nitrification and denitrification processes that produce N2O in litter(Reference de Klein and Eckard265–Reference Pereira, Martins and Bonifácio267). For other types of livestock manures, it has been shown that N content is less of a limiting factor to N2O formation and emission processes in manure compared to other factors including density, pH, temperature, manure mass, moisture content and organic carbon content(Reference Redding, Devereux and Phillips268). For this reasons, it has been suggested that reducing N2O emissions is more likely to be successful with urease and nitrification inhibitors(Reference de Klein and Eckard265,Reference Ledgard, Menneer and Dexter269,Reference Gerber, Hristov and Henderson270) than with dietary controls(Reference Wiedemann, Phillips and Naylor263).
Conclusions and outlook
The global demand for chicken-meat is increasing in a context where it is fundamental to produce economically while complying with societal imperatives of environmental sustainability. The nutrition of the broiler chicken is the main component of this complex challenge and requires novel approaches to feed formulation that target not only the chicken but also their internal and external environment. Improved use of locally produced ingredients with lower carbon footprint has the potential to reduce the reliance on imported ingredients, notably SBM, whereby the choice of alternative ingredients will vary with geographic locations and local availability. Overall, the future path for chicken-meat sustainability needs to look at improving feed formulations targeting higher efficiencies in the digestion of protein and carbohydrates. This requires a better understanding of the digestion dynamics of these nutrients (i.e., protein, polysaccharides and interactions), the inclusion of improved crop varieties (e.g., canola, sorghum) and feed processing (e.g., particle size), and the application of feed additives (i.e., enzymes, crystalline AAs, pre and pro-biotics). Reduced CP diets are likely to contribute to the solution towards improving environmental sustainability of chicken-meat production, including at the level of poultry houses through reducing the amount of water and N being excreted by the chickens. Finally, we need to continue to explore and harness the role of nutrition in strengthening gut health and improving gut microbiota function to improve performance while increasing resistance to pathogens and environmental stressors.
Acknowledgement
This manuscript has been developed inspired by the AgriFutures Chicken Meat Consortium – Nutrition, Gut Health and Environment project (PRO-016111), which is a multi-institutional and multi-national collaboration, part of the AgriFutures Chicken Meat Program.
Financial support
The Consortium team acknowledges funding and support from all stakeholders. It includes AgriFutures Australia (PRO-016111) with its associated levy payers, The University of Queensland, The University of Sydney, Central Queensland University, Department of Primary Industries Queensland, Massey University, Aviagen, dsm-firmenich, Lucta, Lallemand, Poultry Research Foundation, World Poultry Science Association New Zealand Branch, and another nine international universities.
The authors are thankful to Ms Mila Meijer for her creative work on the visual abstract. The authors would also like to acknowledge the contribution of Dr Stanley Chen in the final formatting of the manuscript. In addition, the authors would like to acknowledge the intellectual contributions of the AgriFutures Chicken Meat Panel.
Competing interests
The authors declare no conflicts of interest.
Authorship
RNZ and ER conceptualized the review and provided supervision; all authors conducted the literature search, synthesized the data and drafted the original manuscript by sections of expertise; all authors contributed to the critical revision, review, and editing of the manuscript. All authors have read and agreed to the published version of the manuscript
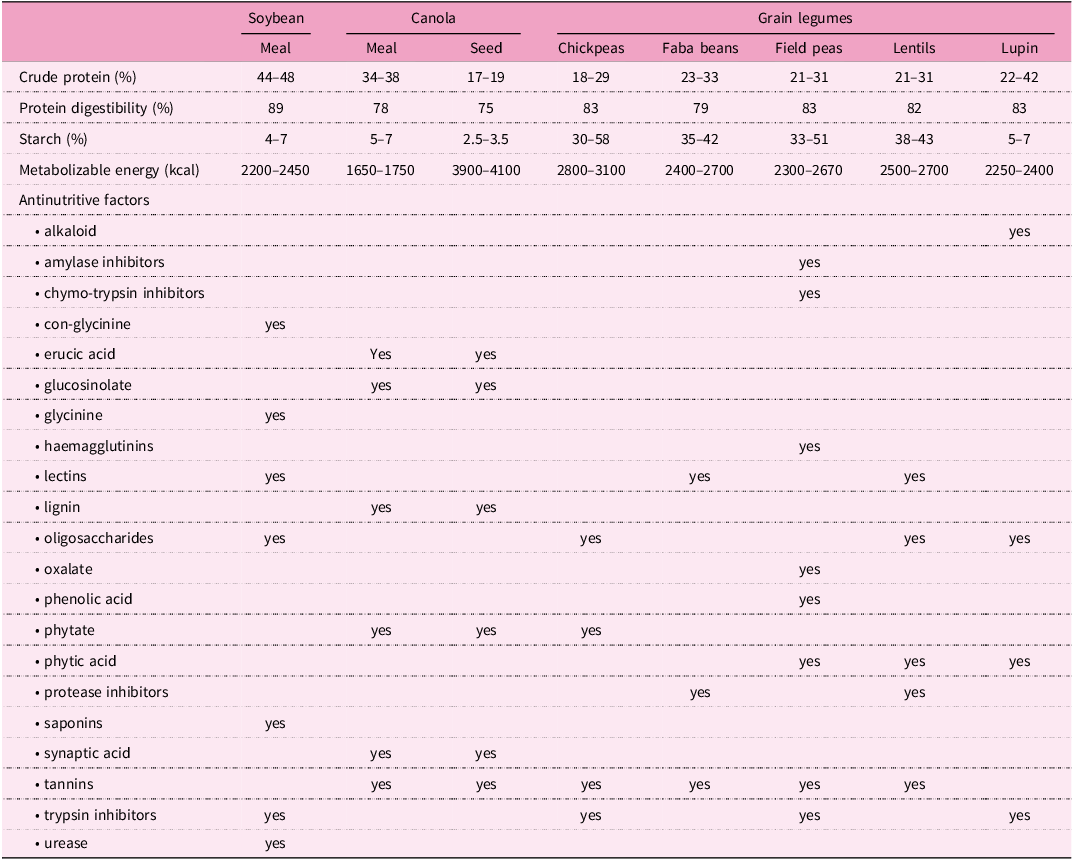
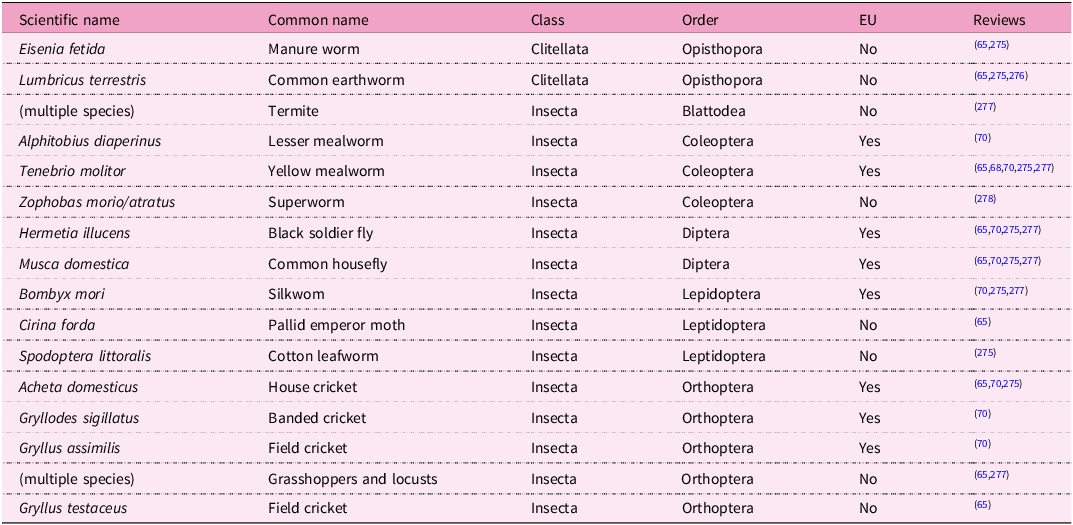
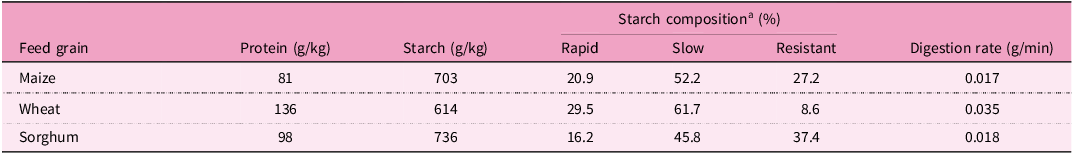
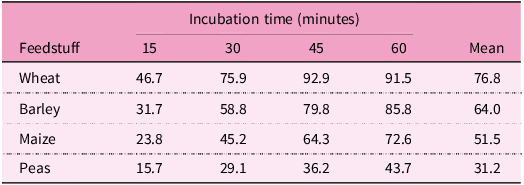
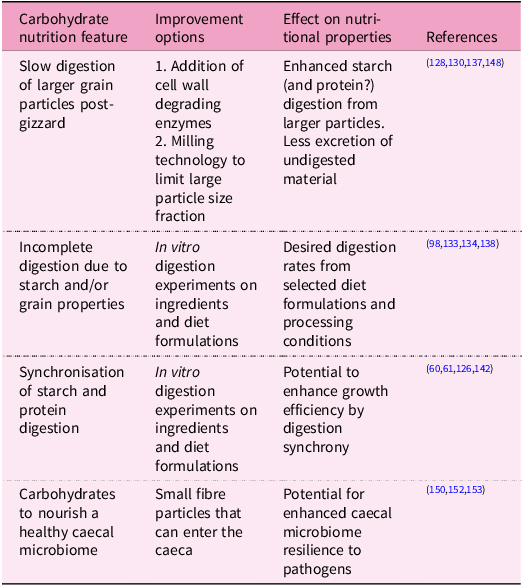
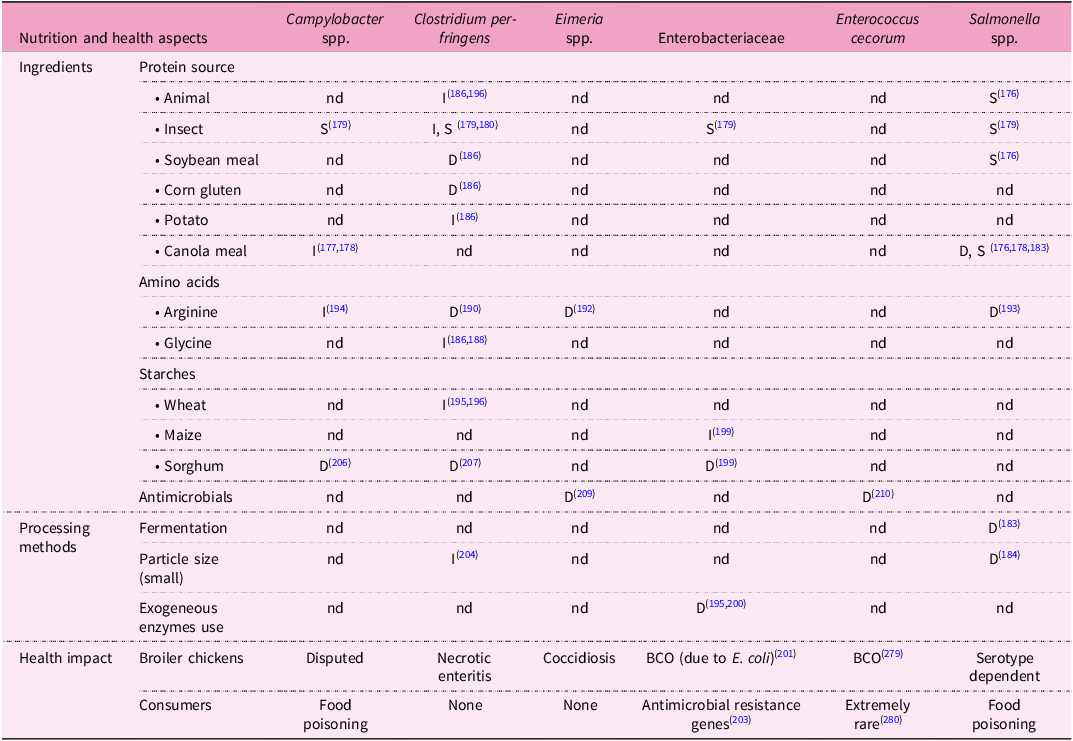

 Open access
Open access