



Introduction
Schistosomiasis is the second most important human parasitic disease globally (Mawa et al., Reference Mawa, Kincaid-smith, Tukahebwa, Webster and Wilson2021), as measured by its impact on public health, and a major waterborne disease globally (Steinmann et al., Reference Steinmann, Keiser, Bos, Tanner and Utzinger2006). The causative agents of schistosomiasis are several species within the phylum Platyhelminthes, class Trematoda, family Schistosomatidae and genus Schistosoma, a group of parasitic flatworms that infect birds and mammals, where they inhabit the vascular system (Anderson and Enabulele, Reference Anderson and Enabulele2021). Three species of schistosomes are of great concern to humans: Schistosoma mansoni, S. japonicum and S. haematobium, of which S. mansoni and S. haematobium account for 99% of human infections (Keitel et al., Reference Keitel, Potter, Diemert, Bethony, El Sahly, Kennedy, Patel, Plieskatt, Jones, Deye, Bottazzi, Hotez and Atmar2019).
Globally, more than 251 million people require treatment for schistosome infection and 779 million people across 75 countries are at risk of infection (Steinmann et al., Reference Steinmann, Keiser, Bos, Tanner and Utzinger2006; World Health Organization, 2026), of which 90% of the disease burden and risk are concentrated in Africa (Okwori et al., Reference Okwori, Sidi, Ngwai, Obiekezie, Makut, Chollom, Okeke and Adikwu2014). Recent estimates from the 2021 Global Burden Disease (GBD) Study place the number of direct deaths from schistosomiasis at approximately 12 800 per annum and 1.75 million disability-adjusted life years (DALYs; Shen and Luo, Reference Shen and Luo2025). Importantly, both the World Health Organization (WHO) and the GBD acknowledge that even these revised figures are likely an underestimate due to persistent underdiagnosis, non-specific clinical symptoms, limited surveillance and inadequate funding and health resources (World Health Organization, 2026). Earlier burden estimates were substantially higher, including up to 280 000 deaths annually in sub-Saharan Africa and as high as 56 million DALYs globally when broader assumptions about chronic morbidity and attributable mortality were applied (King, Reference King2010). This burden is compounded by the association between schistosomiasis and increased susceptibility to HIV infection (Brown et al., Reference Brown, Miiro, Nkurunziza, Watera, Quigley, Dunne, Whitworth and Elliott2006), malaria (Booth et al., Reference Booth, Vennervald, Butterworth, Kariuki, Amaganga, Kimani, Mwatha, Otedo, Ouma and Dunne2004; Sokhna et al., Reference Sokhna, Le Hesran, Mbaye, Akiana, Camara, Diop, Ly and Druilhe2004), salmonellosis (Melhem and LoVerde, Reference Melhem and LoVerde1984) soil-transmitted helminth infections (Molvik et al., Reference Molvik, Heiland, Zulu, Kleppa, Lillebo, Gundersen, Kvalsvig, Taylor, Kjetland and Vennervald2017), and tuberculosis (Brown et al., Reference Brown, Miiro, Nkurunziza, Watera, Quigley, Dunne, Whitworth and Elliott2006). Additionally, schistosomiasis disproportionately affects marginalized and socioeconomically disadvantaged communities, especially children and pregnant women from these populations, reinforcing the cycle of poverty (Pinto-Almeida et al., Reference Pinto-Almeida, Mendes, Ferreira, Abecasis, Belo, Anibal, Allegretti, Galinaro, Carrilho and Afonso2021).
Treatment for S. mansoni infection is limited to oxamniquine and praziquantel, with the latter being the only drug used in mass drug administration (MDA) campaigns due to broad activity against multiple Schistosome species and its ease of availability. Praziquantel is only active against adult S. mansoni and has an extremely short half-life (<3 h) (Kuevi et al., Reference Kuevi, Acquah, Amuquandoh and Abbey2023), leaving the host vulnerable to reinfection, especially from immature S. mansoni parasites still within the host. The lack of effective long-term chemotherapy highlights the as yet unmet need for a vaccine to control S. mansoni. Although vaccine development strategies must ultimately address all major schistosome species, most vaccine research has concentrated on S. mansoni. This species serves as the principal model for understanding schistosome biology and host–parasite interactions as the lifecycle is easily maintained compared to other species (Anderson and Enabulele, Reference Anderson and Enabulele2021) and was one of the first schistosomes to have a complete genome sequence and assembly published (Berriman et al., Reference Berriman, Haas, LoVerde, Wilson, Dillon, Cerqueira, Mashiyama, Al-lazikani, Andrade and Ashton2009).
S. mansoni life cycle
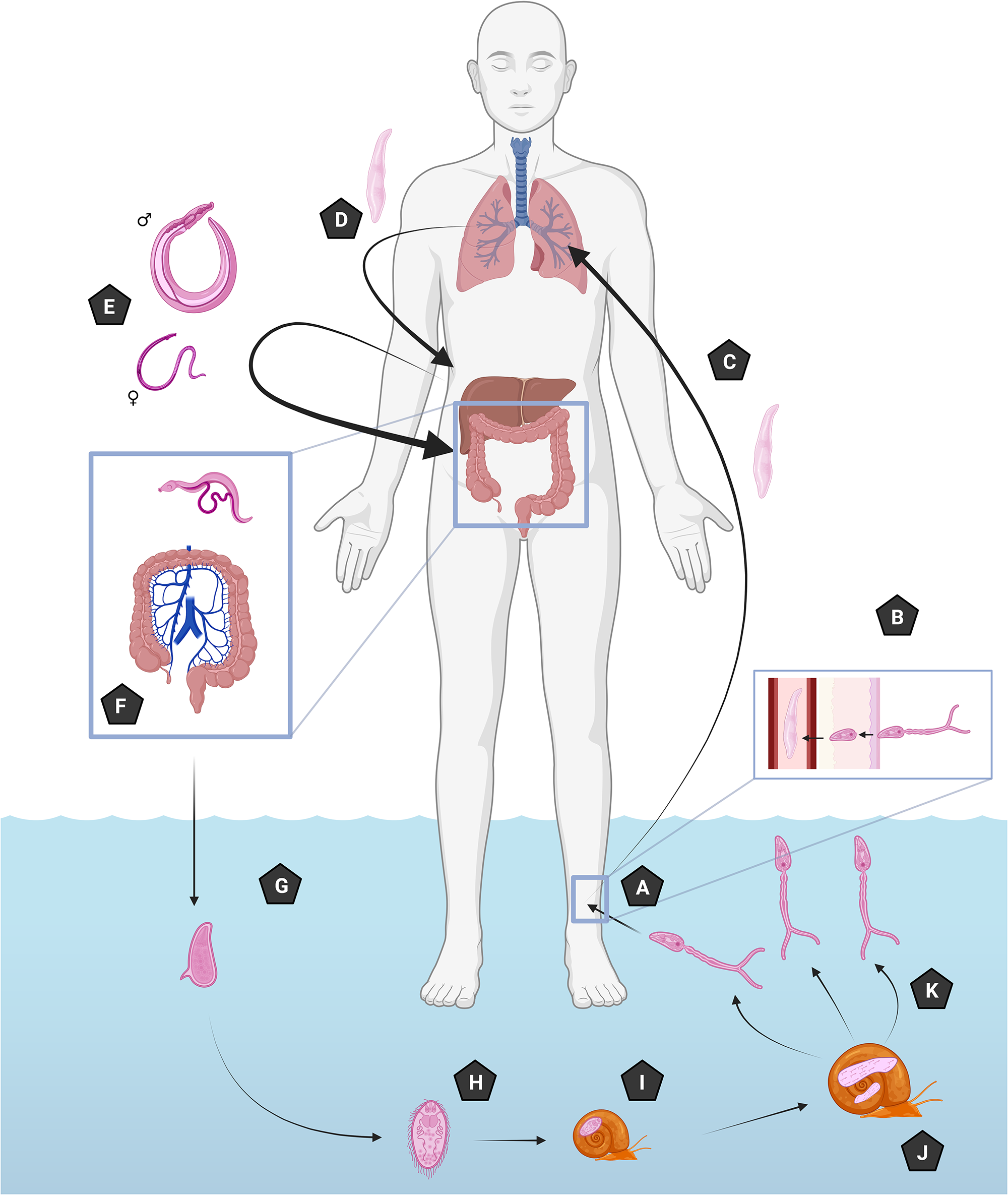
Infection with S. mansoni occurs when cercariae at depths of 25–30 cm in warm, transparent freshwater contact human skin (Panzner et al., Reference Panzner, Utzinger and Keiser2025), taking between 12 and 72 h to penetrate the dermis after initial contact and attachment by the cercariae (Bartlett et al., Reference Bartlett, Brown, Marriott and Whitfield2000) (Figure 1A). After penetration, the cercariae immediately begin to morphologically change into schistosomula (Figure 1B) and migrate to the lungs via the circulatory or lymphatic system to partially mature (Figure 1C). Partially mature schistosomula subsequently migrate again to the liver to undergo further maturation between 3 and 6 days (Figure 1D) before migrating again (Figure 1E) to their final destination in the mesenteric veins of the large intestine (Nation et al., Reference Nation, Da’dara, Marchant and Skelly2020) (Figure 1F). During these migration and maturation stages, hosts can experience moderate to severe flu-like symptoms of fatigue, fever, night sweats or headache, between 2 and 7 weeks after exposure, known as acute schistosomiasis (Langenberg et al., Reference Langenberg, Hoogerwerf, Koopman, Janse, Kos-van Oosterhoud, Feijt, Jochems, de Dood, van Schuijlenburg, Ozir-Fazalalikhan, Manurung, Sartono, van der Beek, Winkel, Verbeek-menken, Stam, Van leeuwen, Meij, van Diepen, van Lieshout, van Dam, Corstjens, Hokke, Yazdanbakhsh, Visser and Roestenberg2020). Unusal for Trematoda infecting humans, schistosomes are dioecious, with larger males permanently enclosing females within their gynaecophoric canal for copulation (Colley et al., Reference Colley, Bustinduy, Secor and King2014; Jamieson, Reference Jamieson2016). Schistosomes live between 5 and 15 years, but in some cases more than 30 years within the human host (Gryseels et al., Reference Gryseels, Polman, Clerinx and Kestens2006). During this period, females produce hundreds of eggs per day (Gryseels et al., Reference Gryseels, Polman, Clerinx and Kestens2006). Schistosome eggs are excreted from the host via faeces (Figure 1G), hatching once in freshwater and releasing ciliated miracidia (Figure 1H). In turn, these miracidia infect a suitable snail host (Figure 1I) and undergo asexual replication through the mother and daughter sporocyst stages for 4–6 weeks (Figure 1J). Eventually, the snails shed tens of thousands of cercariae into the surrounding freshwater that remain infectious for 1–3 days to continue the transmission cycle (Colley et al., Reference Colley, Bustinduy, Secor and King2014) (Figure 1K). Eggs that fail to penetrate the vessel wall and fail to escape the host are transported via the hepatic portal vein to the liver and spleen, where they become trapped, leading to symptoms known as schistosomiasis (chronic infection) and are the primary determinant of schistosomiasis pathology in hosts (Costain et al., Reference Costain, MacDonald and Smits2018) (Figure 1).
S. Mansoni life cycle: (A) Cercariae attach to and progress through the dermis, during this process they transform into schistosomula. (B) Schistosomula migrate to the lungs and undergo partial maturation. (C) Schistosomula migrate to the liver to mature into adult worms. (D) Adult worms migrate from the liver to their final destination. (E) Primarily, the mesenteric venules of the large intestine and monogamous pairs where the females produce approximately 300 eggs per day. (F) Eggs are excreted out of the host and hatch in freshwater, (G) releasing the miracidia. (H) Miracidia seek out and penetrate suitable snail hosts (Biomphalaria species). (I) Miracidia transform into sporocysts within the snail. (J) After several cycles of asexual reproduction infective larvae are released as cercariae into the surrounding water. (K) Cercariae seek out and penetrate human skin.

S. mansoni treatment and control
Only 2 chemotherapeutic anthelminthics, oxamniquine and praziquantel, are approved for treating schistosomiasis caused by S. mansoni. Praziquantel has been the mainstay of MDA campaigns for decades in endemic areas because it is inexpensive, widely available and active against all schistosome species (Hagan et al., Reference Hagan, Appleton, Coles, Kusel and Tchuem-tchuenté2004; Secor and Montgomery, Reference Secor and Montgomery2015), whereas the utility of oxamniquine is limited to S. mansoni and additionally may have limited utility due to reports of pre-existing natural resistance in S. mansoni populations (Valentim et al., Reference Valentim, Cioli, Chevalier, Cao, Taylor, Holloway, Pica-mattoccia, Guidi, Basso, Tsai, Berriman, Carvalho-queiroz, Almeida, Aguilar, Frantz, Hart, LoVerde and Anderson2013). The mechanism of action for praziquantel occurs through the activation of the transient receptor potential melastatin ion channel (TRPMPZQ) in schistosomes, resulting in Ca2+ ions entering the worm uncontrollably, leading to paralysis and death (Park et al., Reference Park, Friedrich, Yahya, Rohr, Chulkov, Maillard, Rippmann, Spangenberg and Marchant2021).
The efficacy of anthelminthic drugs against S. mansoni is assessed by the egg reduction rate (ERR) with a threshold of 90% indicating optimal efficacy (World Health Organization, 2013). Disappointingly, praziquantel is not fully effective, with cure rates reported ranging from 86.6% to 97.9% (Tesfie et al., Reference Tesfie, Getnet, Abere, Yihenew, Belete, Kassa and Gize2020; Berhanu et al., Reference Berhanu, Atnafie, Ali, Chekol and Kebede2022; Gebreyesus et al., Reference Gebreyesus, Makonnen, Tadele, Mekete, Gashaw, Gerba and Aklillu2023; Kabatende et al., Reference Kabatende, Ntirenganya, Mugisha, Barry, Ruberanziza, Bienvenu, Bergman and Aklillu2023). Contributing to this fact is that praziquantel loses effectiveness in moderate and heavy infections (Raso et al., Reference Raso, N’goran, Toty, Luginbühl, Adjoua, Tian-bi, Bogoch, Vounatsou, Tanner and Utzinger2004; Gebreyesus et al., Reference Gebreyesus, Makonnen, Tadele, Mekete, Gashaw, Gerba and Aklillu2023) and infections in school age children who have been treated >7 times in repeated MDA rounds display a decreased praziquantel ERR (Crellen et al., Reference Crellen, Walker, Lamberton, Kabatereine, Tukahebwa, Cotton and Webster2016). Paradoxically, repeated treatments of praziquantel in mice mimicking repeated infection-treatment cycles of MDA programmes has demonstrated a relative reduced susceptibility of hosts to reinfection by S. mansoni with elevated cytokine levels in the liver and reduced egg accumulation (Nono et al., Reference Nono, Mpotje, Mosala, Aziz, Musaigwa, Hlaka, Spangenberg and Brombacher2021).
In endemic regions, the WHO has set guidelines for MDA, aiming to eliminate schistosomiasis as a public health concern. In populations with infections ≥10%, the WHO recommends annual single dose (40 mg kg−1 body weight) of praziquantel with ≥75% coverage for all age groups ≥2 years including pregnant women after first trimester (World Health Organization, 2022). If these endemic regions fail to show a response to annual praziquantel treatment with adequate coverage of age groups the WHO recommends twice yearly dose of praziquantel (World Health Organization, 2022). Some countries, such as Zimbabwe, have shown the effectiveness of praziquantel MDA in school-aged children with a 6-year survey demonstrating near elimination of schistosomiasis in this cohort initially but rebound infections within the following years (Mduluza et al., Reference Mduluza, Jones, Osakunor, Lim, Kuebel, Phiri, Manangazira, Tagwireyi and Mutapi2020). This rebound can be explained due to absence of efforts to break the transmission cycle of S. mansoni in Zimbabwe, additionally the exclusion of preschool and adults from this MDA programme allows for the transmission cycle to remain in the community. Modelling of MDA effects on schistosomiasis reveals there is a positive relationship between prevalence levels and the required coverage and time required to eliminate the disease (Danso-Appiah et al., Reference Danso-Appiah, Eusebi, Lo, Orso, Akuffo, Fleming, Jiangang, Mwinzi, Colley and Hagan2021; Kura et al., Reference Kura, Ayabina, Hollingsworth and Anderson2022), which may not be economically or logistically feasible for countries where schistosomiasis is endemic.
Multi-year MDA programmes have failed to reduce transmission and infection rates among populations in numerous endemic regions (Campbell et al., Reference Campbell, Binder, King, Knopp, Rollinson, Person, Webster, Allan, Utzinger and Ame2020). This lack of progress can be attributed to praziquantel lasting for only hours in the human body and only being able to target adult schistosomes, leaving juvenile worms and eggs unaffected (Zhang et al., Reference Zhang, Le, Ahmad, Molehin, Siddiqui, Torben, Karmakar, Rojo, Sennoune, Lazarus, Khatoon, Freeborn, Sudduth, Rezk, Carey, Wolf, Papin, Damian, Gray, Marks, Carter and Siddiqui2020a). The WHO acknowledges that MDA programmes alone are insufficient to eliminate schistosomiasis (World Health Organization, 2020). Concerns have also been raised regarding MDAs leading to positive selective pressure on S. mansoni, which are increasingly resistant to praziquantel (Berger et al., Reference Berger, Crellen, Lamberton, Allan, Tracey, Noonan, Kabatereine, Tukahebwa, Adriko, Holroyd, Webster, Berriman and Cotton2021). While no definitive mechanism of resistance to praziquantel has been established, laboratory studies investigating praziquantel resistant S. mansoni show 2.25-fold lower expression of TRPMPZQ (Clec’h et al., Reference Clec’h, Chevalier, Mattos, Strickland, Diaz, McDew-White, Rohr, Kinung’hi, Allan, Webster, Webster, Emery, Rollinson, Djirmay, Mashikhi, Yafae, Idris, Moné, Mouahid, LoVerde, Marchant and Anderson2021), increased expression of P-glycoprotein and multi-drug resistance-associated protein efflux pumps (Pinto-Almeida et al., Reference Pinto-Almeida, Mendes, Armada, Belo, Carrilho, Viveiros and Afonso2015).
To date, there has been no successful vaccine for S. mansoni infection. Given the significant global toll of infections, an effective vaccine that can provide long-lasting immunity and reduce dependency on environmental and chemotherapeutic campaigns is critically needed. Expert working groups have proposed the preferred product characteristics for an effective prophylactic vaccine, which should achieve a minimum of 75% reduction in adult worm burden and 75% reduction in egg excretion, provide protection for 2–3 years, be suitable for school-aged children and adults and be compatible with co-administration alongside MDA or other control measures (Molehin et al., Reference Molehin, Rojo, Siddiqui, Gray, Carter and Siddiqui2016). While these targets are ambitious, it has been argued that even a first-generation vaccine conferring 50% protection against worm and egg burden in humans could still substantially reduce schistosomiasis-associated morbidity and mortality (Siddiqui et al., Reference Siddiqui, Siddiqui and Ganley-leal2011). In S. mansoni vaccination studies, vaccine efficacy is commonly reported as the percentage reduction in adult worms recovered and/or eggs collected from faeces or tissues in vaccinated animals relative to contemporaneous challenged unvaccinated or adjuvant/placebo-treated controls. Adult worm burden is commonly reported because adult worms are the source of egg production; therefore, reduction in adult worm survival is expected to reduce both egg deposition in host tissues and egg excretion into the environment. However, worm burden alone is an incomplete efficacy endpoint, because schistosomiasis pathology is driven primarily by tissue-trapped eggs (Costain et al., Reference Costain, MacDonald and Smits2018), while ongoing transmission depends on egg excretion and viability (Kifle et al., Reference Kifle, Kalyanasundaram, Molehin, Balkhi, Arya, Zhang, Ahmad, Torben, Carey, Papin, Jackson, Gray, Carter and Siddiqui2025).
Mathematical modelling further suggests that chemotherapy or vaccination alone is unlikely to eliminate S. mansoni in high-transmission regions and that concurrent vaccination programmes and MDA are needed, with outcomes dependent on vaccine efficacy, coverage and duration of protection (Stylianou et al., Reference Stylianou, Hadjichrysanthou, Truscott and Anderson2017; Kura et al., Reference Kura, Truscott, Toor and Anderson2019). If such an effective vaccine is developed, it would not only represent a leap forward in managing schistosomiasis but also align with the WHO global health goals of sustainable and humane disease control strategies (World Health Organization, 2026).
Historical efforts for the development of a S. mansoni vaccine
S. mansoni has been documented in Egyptian mummies dating back over 5000 years and in 1937 it was estimated to infect 60% of the population of the Nile Delta (modern Egypt) (Barakat, Reference Barakat2013). Work on the first vaccine for S. mansoni began in the 1950s after Olivier and Schneidermann (Reference Olivier and Schneidermann1953) were able to demonstrate the ability to acquire partial protection to S. mansoni infection in 4 strains of mice (Swiss, C57BL/6JN, BALB/cAN, A/LN) through prior exposure and that single sex (male) priming (a model of infection where only a single sex of S. mansoni are used to eliminate the risk of egg production) failed to improve survival (Olivier and Schneidermann, Reference Olivier and Schneidermann1953). Initial vaccine experiments in the 1950–70s primarily focused on irradiation-attenuated cercariae that were exposed to X- or gamma radiation (Radke and Sadun, Reference Radke and Sadun1963; Li Hsü et al., Reference Li Hsü, Hsü and Osborne1969). Low-dose radiation (<20 krad/<200 Gy) prevents cercariae from completing maturation but allows the cercariae to survive to a stunted adulthood (Fukushige et al., Reference Fukushige, Mitchell, Bourke, Woolhouse and Mutapi2015), whereas high-dose radiation prevents cercariae from being able to penetrate the skin or mature (Smithers, Reference Smithers1962; Radke and Sadun, Reference Radke and Sadun1963). Animal studies of irradiated cercariae have demonstrated great efficacy in reducing worm burden in mice by 60–80% (Sher et al., Reference Sher, Coffman, Hieny and Cheever1990; Smythies et al., Reference Smythies, Coulson and Wilson1992; Wynn et al., Reference Wynn, Oswald, Eltoum, Caspar, Lowenstein, Lewis, James and Sher1994) and >50% in non-human primates (Stek et al., Reference Stek, Minard, Da and Hall1981) but never progressed further due to safety concerns of incomplete attenuation of the cercariae and the difficulty of delivering live, irradiated cercariae (Coulson, Reference Coulson, Baker, Muller and Rollinson1997). Additionally, whole worm homogenate (Sadun and Bruce, Reference Sadun and Bruce1964; Murrell et al., Reference Murrell, Dean and Stafford1975) and putative membrane antigens (Maddison et al., Reference Maddison, Slemenda, Chandler and Kagan1978) demonstrated ineffective levels of protection in mouse models and never progressed.
These vaccine candidate studies laid the groundwork for understanding protective immunity and identifying specific antigen molecules that could be targeted in vaccine formulations. Specific antibodies generated by mice to the irradiated cercariae (15- and 50-krad) antigens were later shown to recognize a 23 kDa surface-exposed integral membrane protein (Sm23), a 28 kDa glutathione S-transferase (Sm28-GST), a 30-32 kDa cathepsin B (SmCB), asparaginyl endopeptidase (Sm32), heat shock protein 70 (HSP70), triosephosphate isomerase (SmTPI) and a 97 kDa paramyosin (Sm97) (Richter and Harn, Reference Richter and Harn1993; Richter et al., Reference Richter, Reynolds and Harn1993). This demonstrates that not all antigens on the irradiated cercariae were immunogenic. Furthermore, evidence indicated that immunising mice with irradiated cercariae elicited a delayed-type hypersensitivity response responsible for clearing subsequent challenge cercariae, prompting concerns that a similar hypersensitivity reaction could occur in humans (Ratcliffe and Wilson, Reference Ratcliffe and Wilson1991).
By the 1980s, irradiated cercariae had demonstrated efficacy in animal models, with radiation dose, cercariae number and immunization schedule being optimized, as outlined in the review paper by Richter et al. (Reference Richter, Harn and Matuschka1995), though these results did not progress through to human trials due to failure to reach the necessary reduction in worm burden (Houlder et al., Reference Houlder, da Silva, van Diepen, Amaral, Wilson, Hokke, Roestenberg and Bakker2025). During this time, alternative approaches to inducing immunity were explored in the mouse model, such as using unirradiated adult single-sex schistosome infections (Bickle et al., Reference Bickle, Bain, McGregor and Doenhoff1979; Christensen et al., Reference Christensen, Monrad, Nansen and Frandsen1980), and irradiated adult worms (3–4 weeks old) (Bickle et al., Reference Bickle, Bain, McGregor and Doenhoff1979), which failed to induce any protection against infection. The identification of specific antigens from irradiated cercariae vaccination experiments shifted the field away from expensive whole parasite vaccines to antigen and subunit vaccine research.
Also in the 1980s, the WHO, World Bank and United Nations Development Programme for Research and Training in Tropical Diseases (TDR/WHO) committee funded and fast-tracked research into 6 promising vaccine candidate antigens for S. mansoni: Sm28-GST, Sm97, 62-kDA fragment of myosin (Ir-V5), SmTPI, Sm23 and fatty acid–binding protein (Sm14) (Bergquist and Colley, Reference Bergquist and Colley1998). By the late 1990s, funding for this programme was exhausted with a shift in WHO focus (Tendler and Simpson, Reference Tendler and Simpson2008). Of the 6 fast-tracked antigens, only Sm14 has entered clinical development (Phase 2; NCT05658614) and continues to be supported by Oswaldo Cruz Foundation (FIOCRUZ) and Brazil’s Ministry of Health. The remaining candidates did not consistently reach the ≥40% worm-burden reduction benchmark in mouse models, with reported efficacies for Sm23 25–44% (Da’dara et al., Reference Da’dara, Skelly, Fatakdawala, Visovatti, Eriksson and Harn2002), SmTPI 30–60% (Harn et al., Reference Harn, Mitsuyama, Huguenel, Oligino and David1985, Reference Harn, Gu, Oligino, Mitsuyama, Gebremichael and Richter1992), Ir-V5 75% (35% in baboons) (Soisson et al., Reference Soisson, Masterson, Tom, McNally, Lowell and Strand1992, Reference Soisson, Reid, Farah, Nyindo and Strand1993), Sm97 39% (Pearce et al., Reference Pearce, James, Hieny, Lanar and Sher1988) and Sm28-GST 34–45% (Xu et al., Reference Xu, Verwaerde, Grzych, Fontaine and Capron1991).
Current S. mansoni vaccine status
The development of vaccines against S. mansoni has faced numerous challenges, resulting in a complex landscape of research and candidate formulations. Currently, no effective vaccine is available; however, 3 promising candidates, Sm-TSP-2, Sm14 and Sm-p80/SchistoShield, are undergoing clinical evaluation.
Schistoshield (Sm-p80 + GLA-SE)
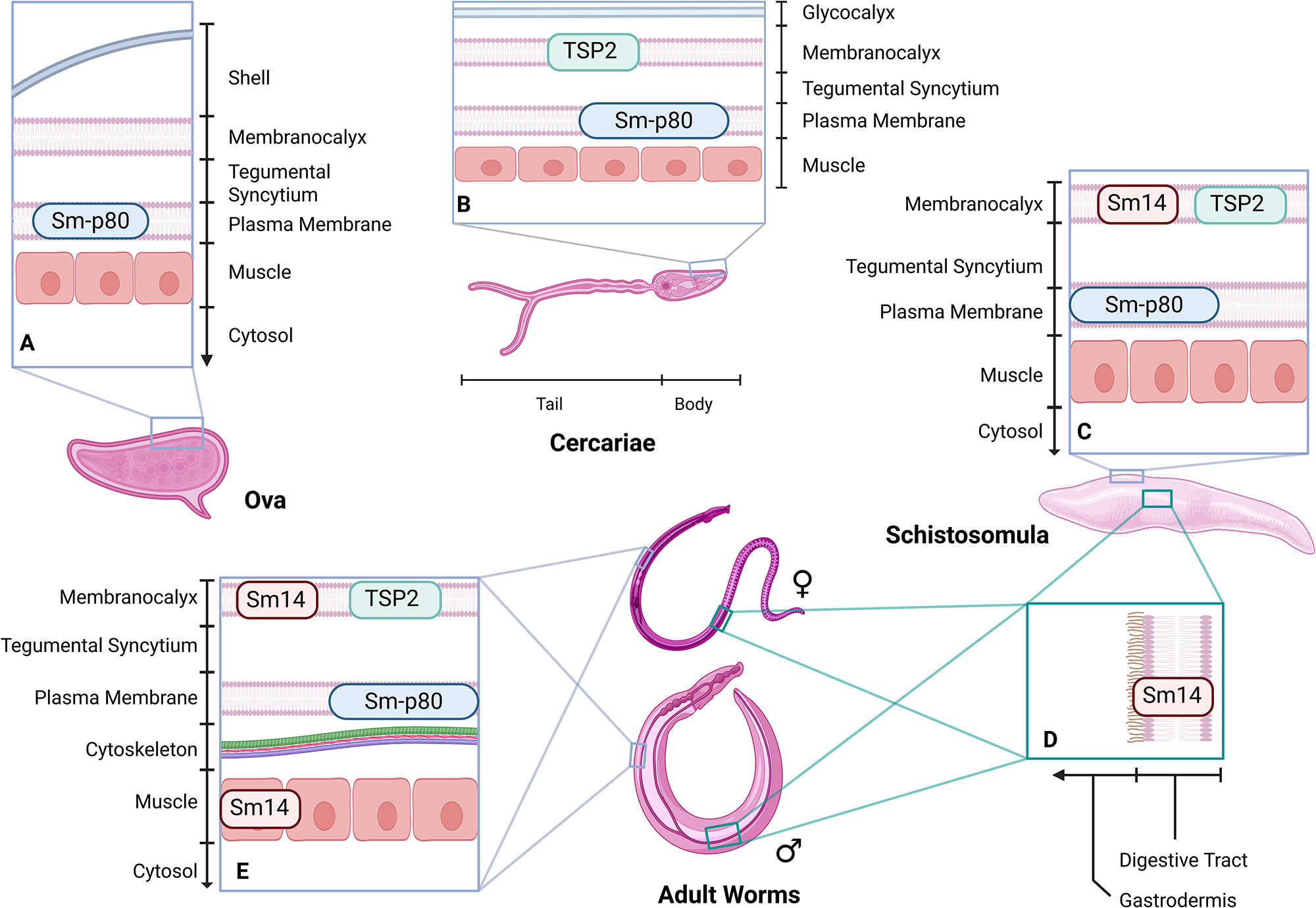
The recombinant protein vaccine candidate of Texas Tech University, ShistoShield, is based on the 5β subunit of the 20S proteasome (Sm-p80), a multi-subunit enzyme involved in tegument membrane turnover (tegument is the collective term for 3 layers: the surface membranocalyx, the underlying tegumental syncytium and the innermost plasma membrane) (Jackson et al., Reference Jackson, Coler, Deye, Carter, Gray, Pecor, Davis, Larsen, Posavad, Cox, Watanabe, Lundeen, Gill, Kalyanasundaram and Siddiqui2025). The 20S proteasome enables schistosomes to shed the tegument to evade the host’s immune system and is central to maintaining homeostasis (Podesta et al., Reference Podesta, Karcz, Ansell and Silva1987). Sm-p80 is found in the plasma membrane of ova, apical point of the cercariae body and ducts, schistosomula and adult worms (Figure 2A–C,E) and is also expressed in the reproductive organs of adult worms (Molehin et al., Reference Molehin, Sennoune, Zhang, Rojo, Siddiqui, Herrera, Johnson, Sudduth, May and Siddiqui2017).
Localization of current vaccine proteins in S. mansoni. (A) Sm-p80 in plasma membrane of ova. (B) TSP-2 in the membranocalyx and Sm-p80 in the plasma membrane of the cercariae. (C) Sm14 and TSP-2 in membrancalyx and Sm-p80 in the plasma membrane of the schistosomula. (D) Sm14 in the gastrodermis of schistosomula and adult worms. (E) Sm14 in the membranocalyx and muscle, TSP-2 in membrancalyx and Sm-p80 in the plasma membrane of adult worms.

Figure 2 Long description
The diagram illustrates the localization of proteins Sm-p80, TSP-2 and Sm14 in different life stages of Schistosoma mansoni. In ova, Sm-p80 is located in the plasma membrane. In cercariae, TSP-2 is in the membranocalyx and Sm-p80 is in the plasma membrane. In schistosomula, Sm14 and TSP-2 are in the membranocalyx, while Sm-p80 is in the plasma membrane. In adult worms, Sm14 is found in the gastrodermis and muscle, TSP-2 in the membranocalyx and Sm-p80 in the plasma membrane. The diagram includes cross-sectional views of the tegument layers, showing the membranocalyx, tegumental syncytium and plasma membrane, along with muscle and cytosol layers.
SchistoShield combines Sm-p80 with glucopyranosyl lipid adjuvant–stable emulsion (GLA-SE), an adjuvant formulation containing a potent toll-like receptor 4 agonist in a squalene oil-in-water emulsion (Behzad et al., Reference Behzad, Huckriede, Haynes, Gentleman, Coyle, Wilschut, Kollmann, Reed and McElhaney2012; Kim et al., Reference Kim, Davis, Lee, Cho, Yang, Yang, Bae, Son, Kim, Whittington, Siddiqui, Carter and Gray2024; Jackson et al., Reference Jackson, Coler, Deye, Carter, Gray, Pecor, Davis, Larsen, Posavad, Cox, Watanabe, Lundeen, Gill, Kalyanasundaram and Siddiqui2025). The vaccine has completed phase 1 dose-escalation trial in healthy adults in the US (NCT05292391), where adjuvant formulations were demonstrated to be well tolerated and immunogenic (Jackson et al., Reference Jackson, Coler, Deye, Carter, Gray, Pecor, Davis, Larsen, Posavad, Cox, Watanabe, Lundeen, Gill, Kalyanasundaram and Siddiqui2025; Kifle et al., Reference Kifle, Kalyanasundaram, Molehin, Balkhi, Arya, Zhang, Ahmad, Torben, Carey, Papin, Jackson, Gray, Carter and Siddiqui2025). A phase 1b trial in Burkina Faso and Madagascar (NCT05762393) is evaluating safety, tolerability and immunogenicity in adults from endemic populations, with results still to be reported. A phase 2 controlled human infection study (NCT05999825) designed to assess the safety, immunogenicity and preliminary efficacy of 30 µg Sm-p80 plus 5 µg GLA-SE is currently underway using a single-sex challenge model with male cercariae. The controlled human infection methodology for the phase 2 trial and good manufacturing practices for ShistoShield have been published (Abaasa et al., Reference Abaasa, Egesa, Driciru, Koopman, Kiyemba, Sanya, Nassuuna, Ssali, Kimbugwe, Wajja, van Dam, Corstjens, Cose, Seeley, Kamuya, Webb, Yazdanbakhsh, Kaleebu, Siddiqui, Kabatereine, Tukahebwa, Roestenberg and Elliott2023; Kim et al., Reference Kim, Davis, Lee, Cho, Yang, Yang, Bae, Son, Kim, Whittington, Siddiqui, Carter and Gray2024).
Schistovac (Sm14)
The FIOCRUZ’s Sm14/GLA-SE vaccine, is based on the 14.8-kDa fatty acid–binding protein that is expressed in the gut epithelium and tegument of S. mansoni schistosomula, with adult worms also displaying Sm14 in muscle (Figure 2C-E) (Moser et al., Reference Moser, Tendler, Griffiths and Klinkert1991; Brito et al., Reference Brito, Oliveira, Oliveira, Street, Riengrojpitak, Wilson, Simpson and Correa-oliveira2002). Fatty acid–binding proteins are critical to the survival of schistosomes, and S. mansoni is known to be unable to synthesize fatty acids and sterols de novo but can create phospholipids and triacylglycerols from precursors obtained from the host (Brouwers et al., Reference Brouwers, Smeenk, van Golde and Tielens1997). These proteins are crucial for schistosomes, as they rely entirely on their host for sterols and fatty acids because of the absence of oxygen-dependent synthesis mechanisms that facilitate the uptake, transport and compartmentalization of these essential molecules (Tendler and Simpson, Reference Tendler and Simpson2008), which are required for energy production and membrane synthesis (Moser et al., Reference Moser, Tendler, Griffiths and Klinkert1991; Perally et al., Reference Perally, Geyer, Farani, Chalmers, Fernandez-fuentes, Maskell, Hulme, Forde-thomas, Phillips, Farias, Collins and Hoffmann2021). Another key function of these fatty acid–binding proteins is to incorporate host lipids into the outer membrane to mask and dampen the inflammatory immune response produced by the schistosomula and adult worm (Chayé et al., Reference Chayé, Gasan, Ozir-Fazalalikhan, Scheenstra, Zawistowska-deniziak, van Hengel, Gentenaar, Manurung, Harvey and Codée2023).
Sm14/GLA-SE has completed phase 1a trials (NCT01154049) in healthy Brazilian adults and demonstrated safety, tolerability and the ability to induce significant IgG antibody and Th1/Th2 cytokine responses without increasing IgE levels (Santini-Oliveira et al., Reference Santini-Oliveira, Coler, Parra, Veloso, Jayashankar, Pinto, Ciol, Bergquist, Reed and Tendler2016). A phase 1b trial (NCT01154049) of the Sm14/GLA-SE schistosomiasis vaccine in healthy young women demonstrated that the vaccine was safe, well-tolerated and induced marked induction of anti-Sm14 IgG1, IgG2 and IgG3 responses but no increase in IgE, and increases in tumour necrosis factor-α, interferon (IFN)-γ and interleukin (IL)-2 CD4+ T-cell cytokine responses (Santini-Oliveira et al., Reference Santini-Oliveira, Machado Pinto, Santos, Vilar, Grinsztejn, Veloso, Paes-de-almeida, Amaral, Ramos, Marroquin-Quelopana, Coler, Reed, Ciol, Savino, Parra, Almeida and Tendler2022). Phase 2 trials (NCT05658614, NCT03799510) have been completed with published results demonstrating an acceptable safety profile and a robust immunogenicity in both adults (Phase 2a) and school children (Phase 2b) (Ly et al., Reference Ly, Diop, Diop, Schacht, Mbengue, Diagne, Guisse, Dompnier, Messias, Coler, Ramos, Tendeng, Ndiaye, Marroquin-Quelopana, de Carvalho Parra, Dos Santos, Sirianni Dos Santos Almeida, Mendes-da-cruz, Reed, Savino, Riveau and Tendler2025). These 2 cohorts developed an antigen-specific immune response after vaccination with elevated IgG levels reported; allowing for the vaccine to progress through to phase 3 trails, which at time of writing has not been reported.
Baylor College of Medicine (Sm-TSP-2)
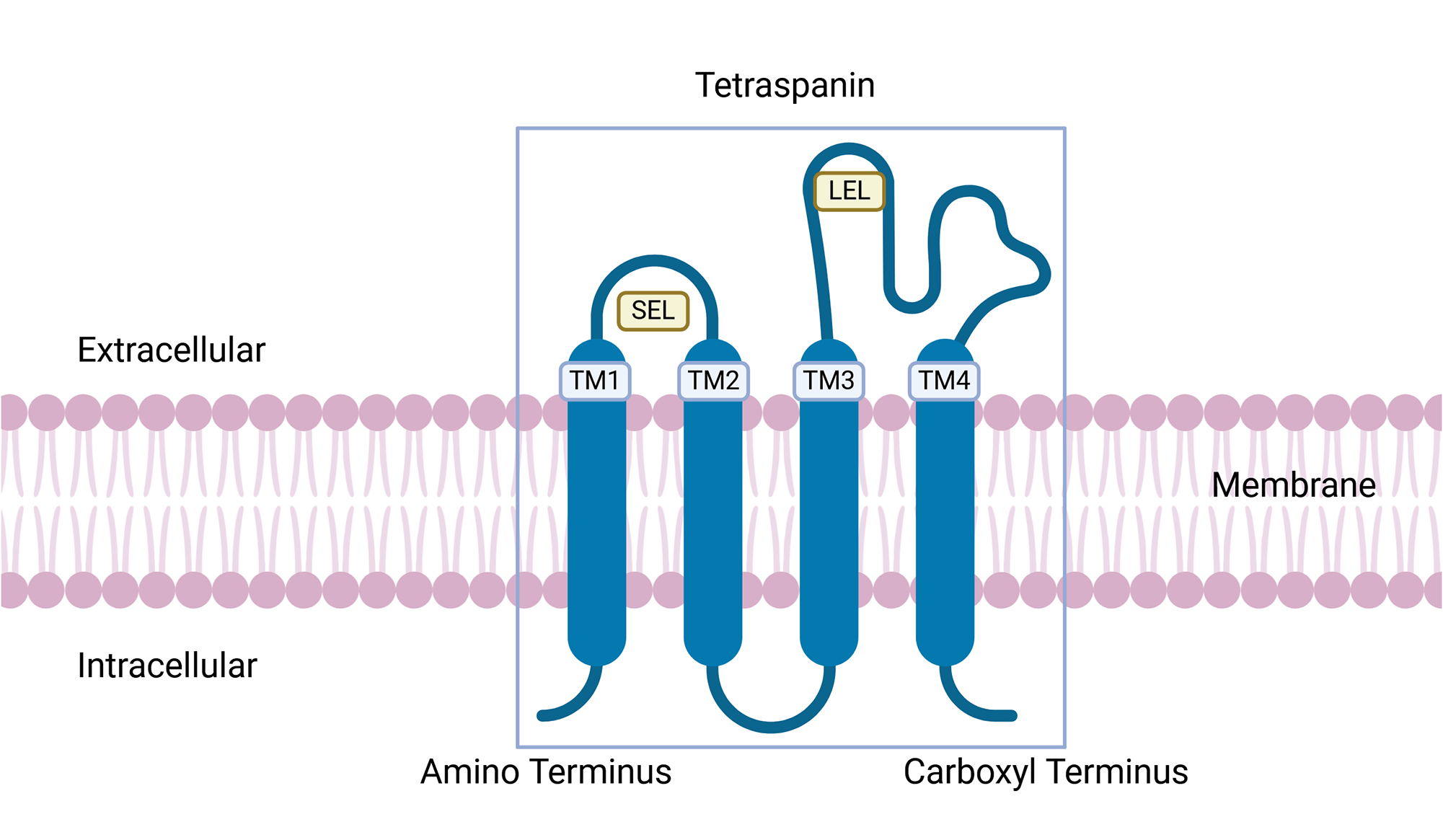
The Baylor College of Medicine’s Sm-TSP-2/Alhydrogel (Al) vaccine targets the tetraspanin 2 (TSP-2) proteins, a member of a group of evolutionarily conserved surface proteins (Boucheix and Rubinstein, Reference Boucheix and Rubinstein2001), that are abundant in the tegument of cercariae, schistosomula and adult worms (Figure 2B,C,E; Cai et al., Reference Cai, Bu, Wang, Wang, Zhong and Wang2008; Jia et al., Reference Jia, Schulte, Loukas, Pickering, Pearson, Mobli, Jones, Rosengren, Daly, Gobert, Jones, Craik and Mulvenna2014). The structure of tetraspanin is characterized by the presence of 4 transmembrane domains, where domains 1 and 2 connect to a small extracellular loop and domains 3 and 4 connect to a large extracellular loop (Figure 3). TSP-2 functions to support maturation, development and maintenance the structural integrity of the schistosomes tegument (Tran et al., Reference Tran, Freitas, Cooper, Gaze, Gatton, Jones, Lovas, Pearce and Loukas2010). Silencing the gene for TSP-2 leads to a thinner tegument that is more vacuolated compared to controls. In murine studies, vaccination experiments showed a 83% reduction in adult worm survival rates and large-scale degradation of the tegument (Tran et al., Reference Tran, Freitas, Cooper, Gaze, Gatton, Jones, Lovas, Pearce and Loukas2010).
Tetraspanin structure. Tetraspanins contain 4 transmembrane domains (TM1-4), a small and large extracellular loop (SEL and LEL, respectively). Sm-TSP-2/Al vaccine antigen.

The Sm-TSP-2/Al vaccine uses the large extracellular loop antigen (Figure 3A) and is formulated on the aluminium hydroxide adjuvant Al, which binds proteins to its surface without significant structural changes for efficient presentation to the immune system (Harris et al., Reference Harris, Soliakov, Lewis, Depoix, Watkinson and Lakey2012) and has been widely approved and used in several commercial vaccines (Fan et al., Reference Fan, Jin, Gilmartin, Toth, Hussein and Stephenson2022). The phase 1a clinical trial (NCT02337855) of Sm-TSP-2/Al vaccine in naïve adult participants from the USA demonstrated that the vaccine is well tolerated and safe in humans (Keitel et al., Reference Keitel, Potter, Diemert, Bethony, El Sahly, Kennedy, Patel, Plieskatt, Jones, Deye, Bottazzi, Hotez and Atmar2019). The phase 1b clinical trial (NCT03110757) of 60 healthy adults aged between 18 and 50 years from Americaninhas, Brazil, has already concluded with published results demonstrating that the vaccine is safe, minimally reactogenic and elicits significant IgG and IgG subclass responses against the vaccine antigen (Diemert et al., Reference Diemert, Correa-Oliveira, Fraga, Talles, Silva, Patel, Galbiati, Kennedy, Lundeen, Gazzinelli, Li, Hoeweler, Deye, Bottazzi, Hotez, El Sahly, Keitel, Bethony and Atmar2023). Sm-TSP-2/Al is currently undergoing phase 2b dose-escalation safety, efficacy and immunogenicity studies in endemic regions of Uganda (NCT03910972), but no results have been published at time of writing.
Lessons learned/Future prospects
Multi-epitope approach
As early as 1998, it has been suggested that a single-antigen approach will not achieve the required protection from schistosome infection and that a multiple antigen or epitope vaccine will be needed (Bergquist and Colley, Reference Bergquist and Colley1998). Furthermore, short peptide vaccines, which are easily manufactured, generally exhibit a well-tolerated safety profile and are less expensive to develop and commercialize. However, these vaccines suffer from short-lived biological activity. This is partly due to their susceptibility to enzymatic degradation and inefficient transport to lymph nodes (Chen et al., Reference Chen, Xu, Zhou, Xu, Varley, Golubovic, Lu, Wang, Yeganeh, Vosoughi and Li2023) precluding their development as an effective vaccine.
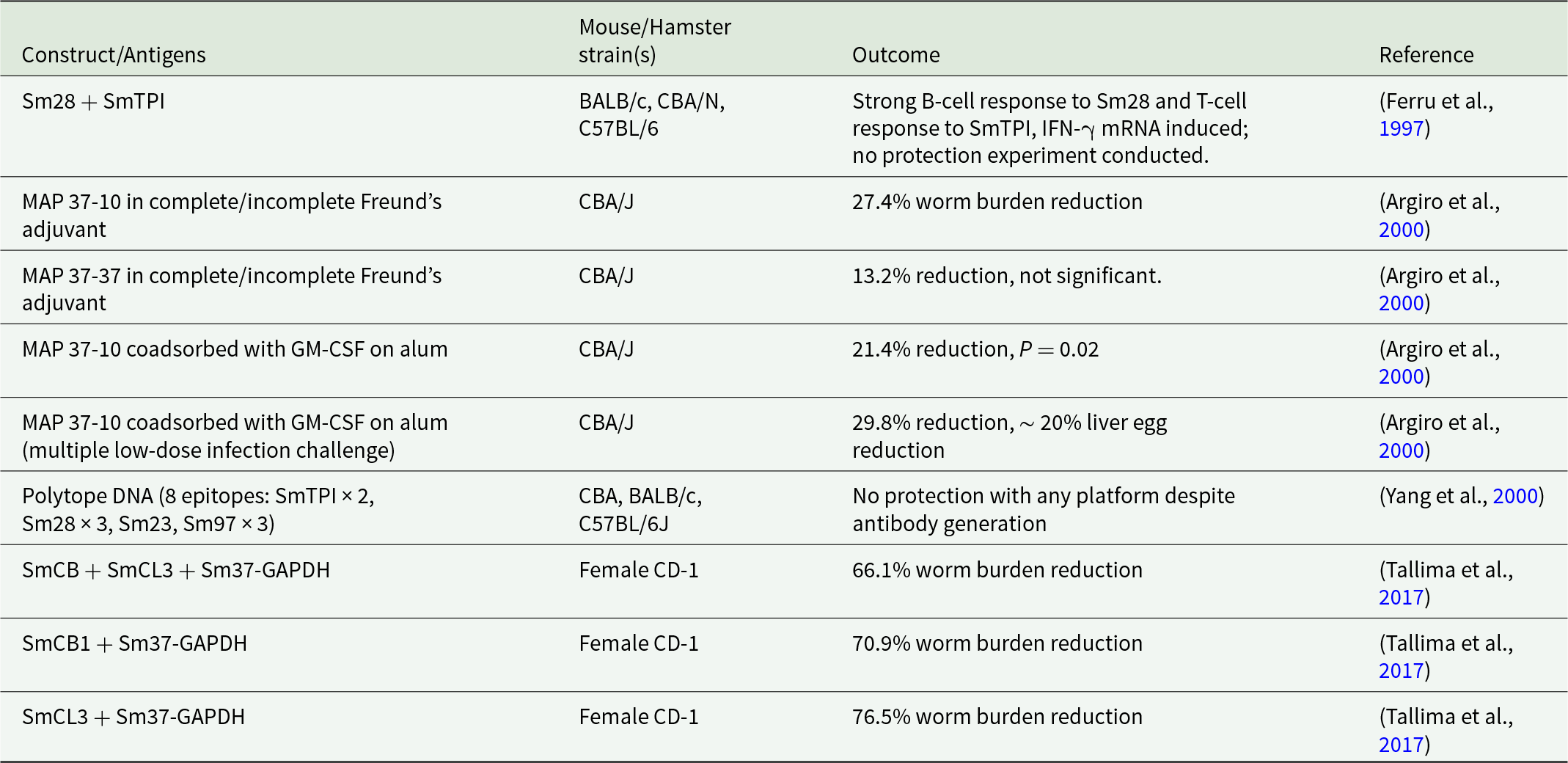
Multi-epitope, also termed multi-antigen or multiple antigen peptide (MAP) vaccines offer the possibility of targeting several antigens and epitopes conserved not just in S. mansoni but across multiple species of schistosomes (Sanches et al., Reference Sanches, Tiwari, Ferreira, Oliveira, Lopes, Passos, Maia, Taranto, Kato, Azevedo and Lopes2021). The first publications in this field began in 2000, investigating a broad range of epitopes and antigens including SmTPI, Sm28-GST, Sm97, Sm23, 37-kDa glyceraldehyde-3-phosphate dehydrogenase (SG3PDH/Sm37-GAPDH) and dynein light chain (Sm10-DLC) (Ferru et al., Reference Ferru, Georges, Bossus, Estaquier, Delacre, Harn, Tartar, Capron, Grassmasse and Auriault1997; Argiro et al., Reference Argiro, Henri, Dessein, Kouriba, Dessein and Bourgois2000; Yang et al., Reference Yang, Jackson, Zeng and McManus2000). These studies examined various epitope combinations: the Sm37-GAPDH major B-cell epitope 5 (SGPLLKGILEYTEDEVVSSDFCGDK, Sm37-5), the Sm37-GAPDH T-cell epitope 3 (GVFTTIDKAQAHIKN, Sm37-3) and the Sm10-DLC T-cell epitope (DKYEIEKDVAAYIKKEFDRKY, Sm10-T). Two key MAP constructs were developed: MAP 37-37, containing the B-cell epitope Sm37-5 and the T-cell epitope Sm37-3; and MAP 37-10, containing the Sm37-5 B-cell epitope paired with the Sm10-T T-cell epitope (Ferru et al., Reference Ferru, Georges, Bossus, Estaquier, Delacre, Harn, Tartar, Capron, Grassmasse and Auriault1997; Argiro et al., Reference Argiro, Henri, Dessein, Kouriba, Dessein and Bourgois2000; Yang et al., Reference Yang, Jackson, Zeng and McManus2000). The outcomes of these studies are summarized in Table 1. Some later publications proposed the design of novel multi-epitope vaccines using what was termed immunomics, a systematic approach to vaccine design that assesses each component and the whole construct of the multi-epitope vaccine against immunomic profiles of humans and animals to predict the immunogenicity and safety profile of the novel vaccine (Gaze et al., Reference Gaze, Driguez, Pearson, Mendes, Doolan, Trieu, McManus, Gobert, Periago, Oliveira, Cardoso, Oliveira, Nakajima, Jasinskas, Hung, Liang, Pablo, Bethony, Felgner and Loukas2014).
Early multi-epitope vaccine candidates and outcomes for S. mansoni

Table 1 Long description
The table summarizes early multi-antigen vaccine constructs tested against Schistosoma mansoni in mice, listing the animal strains used and reported immune or protection outcomes with citations. SmCL3 combined with Sm37-GAPDH in female CD-1 mice produced the largest worm burden reduction at 76.5%, followed by SmCB1 plus Sm37-GAPDH at 70.9% and SmCB plus SmCL3 plus Sm37-GAPDH at 66.1%. In CBA/J mice, MAP 37-10 delivered with Freund’s adjuvant reduced worm burden by 27.4%, while MAP 37-37 showed a 13.2% reduction that was not significant. Adding GM-CSF on alum to MAP 37-10 improved reductions to 21.4% with a reported p value of 0.02, and a multiple low-dose challenge yielded 29.8% worm reduction plus about 20% liver egg reduction. A Sm28 plus SmTPI construct elicited strong antibody responses to Sm28 and T-cell responses to SmTPI with induced interferon gamma messenger RNA, but no protection study was performed. A polytope DNA vaccine spanning eight epitopes generated antibodies yet did not protect across CBA, BALB/c, and C57BL/6J mice. Comparisons should be made cautiously because constructs, adjuvants, challenge methods, and mouse strains differ across studies.
More recently, studies on multi-epitope and multi-antigen vaccines have progressed to de novo designs of chimeric proteins that incorporate multiple epitope subunits from both identified and predicted proteins from S. mansoni (Pinheiro et al., Reference Pinheiro, Ribeiro, Cardoso, Martins, Figueiredo, Assis, Morais, Caliari, Loukas and Oliveira2014; Rahmani et al., Reference Rahmani, Baee, Rostamtabar, Karkhah, Alizadeh, Tourani and Nouri2019) that seek to provide a broader and more robust immune response. Almost all current multi-epitope vaccine designs have been limited to in silico designs and analysis, with designs combining Sm14, Sm21.7 (also known as tegument-allergen-like protein – SmTAL), Sm23, Sm29, Sm-p80, SmCB, HSP70 and Sm-TSP-2 epitopes into unique combinations but not progressing to laboratory experimentation and evaluation (Gaze et al., Reference Gaze, Driguez, Pearson, Mendes, Doolan, Trieu, McManus, Gobert, Periago, Oliveira, Cardoso, Oliveira, Nakajima, Jasinskas, Hung, Liang, Pablo, Bethony, Felgner and Loukas2014; Rahmani et al., Reference Rahmani, Baee, Rostamtabar, Karkhah, Alizadeh, Tourani and Nouri2019; Sanches et al., Reference Sanches, Tiwari, Ferreira, Oliveira, Lopes, Passos, Maia, Taranto, Kato, Azevedo and Lopes2021; Pandya and Kumar, Reference Pandya and Kumar2023).
While most multi-epitope vaccines published remain in the realm of in silico studies, some have progressed to animal studies, including MAP 37-37 multi-epitope vaccines paired with Freund’s adjuvant or aluminium hydroxide (Argiro et al., Reference Argiro, Henri, Dessein, Kouriba, Dessein and Bourgois2000). MAP 37-37 failed to induce any significant protection in vaccinated mice; however, MAP 37-10 vaccinated mice had a reduced worm burden of between 20% and 30% compared to controls (Argiro et al., Reference Argiro, Henri, Dessein, Kouriba, Dessein and Bourgois2000). Protection in MAP 37-10 vaccinated mice was similar for both Freund’s adjuvant and aluminium adjuvants.
Another multi-epitope vaccine that was used in animal studies combined cathepsin B1 (SmCB) and cathepsin L3 (SmCL3) proteins, both of which are expressed in the digestive tract of S. mansoni, and individually demonstrated reductions in worm burden of 58.7% and 65.4%, and a reduction in intestinal egg counts of 37.3% and 28.6% (Tallima et al., Reference Tallima, Dvořák, Kareem, Dahab, Aziz, Dalton and Ridi2017), respectively. Sm37-GAPDH has shown variable efficacy in different mouse strains (El Ridi et al., Reference El Ridi, Mahrous, Afifi, Montash, Velek and Ježek2001). Mice vaccinated with the SmCB + SmCL3 + Sm37-GAPDH vaccine demonstrated a reduction in worm burden by 76% and resulted in >80% dead ova in the intestines, vastly outperforming the individual proteins (Tallima et al., Reference Tallima, Dvořák, Kareem, Dahab, Aziz, Dalton and Ridi2017).
Work on these and other potential vaccine candidates has stagnated, with no new vaccine candidates reaching animal testing. This can be attributed to the complexity of S. mansoni, the cost involved in designing, synthesising and testing a novel vaccine and the limited funds available for neglected tropical diseases.
In silico advances
Much of what has enabled the design and creation of multi-epitope and antigen vaccines is the advancements in reverse vaccinology, also called immunoinformatics, a genome-based approach to vaccine design that makes use of in silico tools to identify potential antigenic targets directly from the genomic sequence. Central to the success of reverse vaccinology has been the rapid development and improvement of several key predictive models, ranging from modelling of discontinuous B- and T-cell epitopes (DiscoTope3 (Høie et al., Reference Høie, Gade, Johansen, Würtzen, Winther, Nielsen and Marcatili2024, 3), BepiPred 3 (Clifford et al., Reference Clifford, Høie, Deleuran, Peters, Nielsen and Marcatili2022), NetMHCIIPan (Reynisson et al., Reference Reynisson, Alvarez, Paul, Peters and Nielsen2020), etc.), protein structure and folding (AlphaFold 2 (Jumper et al., Reference Jumper, Evans, Pritzel, Green, Figurnov, Ronneberger, Tunyasuvunakool, Bates, Žídek, Potapenko, Bridgland, Meyer, Kohl, Ballard, Cowie, Romera-paredes, Nikolov, Jain, Adler, Back, Petersen, Reiman, Clancy, Zielinski, Steinegger, Pacholska, Berghammer, Bodenstein, Silver, Vinyals, Senior, Kavukcuoglu, Kohli and Hassabis2021), HelixFold (Fang et al., Reference Fang, Wang, Liu, He, Lin, Xiang, Zhang, Wu, Li and Song2022), ESMFold (Hie et al., Reference Hie, Candido, Lin, Kabeli, Rao, Smetanin, Sercu and Rives2022; Verkuil et al., Reference Verkuil, Kabeli, Du, Wicky, Milles, Dauparas, Baker, Ovchinnikov, Sercu and Rives2022)) to predicting toxicity (Toxinpred 3 (Rathore et al., Reference Rathore, Choudhury, Arora, Tijare and Raghava2024)), vaccine target identification in humans (Vaxign2 (Ong et al., Reference Ong, Cooke, Huffman, Xiang, Wong, Wang, Seetharaman, Valdez and He2021)) and immunogenicity in humans (C-ImmSim (Rapin et al., Reference Rapin, Lund and Castiglione2011)). These tools, among others, are used to design, test and improve possible vaccines rapidly before expensive and time-consuming production and laboratory testing is undertaken. Reverse vaccinology seeks to decrease the time and cost of vaccine development while simultaneously improving the efficacy of vaccines and search a broader range of data, the genome, for targets that can be missed using conventional methods (Mortazavi et al., Reference Mortazavi, Molaei and Fard2024).
Further enhancing the impact of these powerful models and artificial intelligence approaches to make the design and in silico testing of novel vaccines possible is the fact that most of these models are open source, hosted online and freely available to anyone. The Technical University of Denmark (DTU) hosts more than 75 bioinformatic tools and datasets (https://services.healthtech.dtu.dk/) that can be downloaded for local use or used via the web interface by anyone (DTU Health Tech, n.d..). This is just a single example from an array of hosted tools that can be used in the design and refinement of vaccines, which have dramatically reduced the cost, required coding and specialty knowledge, and local infrastructure to identify, design and test novel in silico uni- and multivalent vaccines for diseases that have been neglected by research funding.
De novo multi-epitope proteins designed using reverse vaccinology almost universally make use of linker motifs to connect the selected or screened epitopes, connecting individual epitopes or sections of discontinuous epitopes into a singular protein construct (Chen et al., Reference Chen, Zaro and Shen2013). Linker motifs utilized to join different epitopes can be categorized into flexible linkers, rigid linkers or in vivo cleavage linkers, with each linker influencing the overall structural integrity and shape, being used to optimize immunogenicity and immune processing and presentation through appropriate spacing of epitopes (Chen et al., Reference Chen, Zaro and Shen2013). The influence of each linker used in a multivalent vaccine design can now be rapidly tested to predict the structure and function using freely hosted in silico tools taking as little as a few minutes to produce a highly accurate model to further evaluate the theoretical vaccines, which has been employed in applications such as malaria vaccine research (Shamriz et al., Reference Shamriz, Ofoghi and Moazami2016).
There are multiple examples of reverse vaccinology in the design of novel vaccines for S. mansoni; Sanches et al. (Reference Sanches, Tiwari, Ferreira, Oliveira, Lopes, Passos, Maia, Taranto, Kato, Azevedo and Lopes2021) and Pandya and Kumar (Reference Pandya and Kumar2023), for example, have proposed radically different multi-epitope vaccine designs, employing some of the previously mentioned in silico models to narrow the selection of epitopes for inclusion, predict immunogenicity, select and test linkers and simulate the immune response to vaccination. Notably, there are endeavours to develop a pan-schistosome vaccine for all human infecting species utilising reverse vaccinology and evolutionarily conserved targets that are accessible to the host’s immune system (Gandvi et al., Reference Gandvi, Jyotishi, Patel and Gupta2025). While the proposed multi-epitope vaccines hold promise as novel and potentially more effective vaccines for S. mansoni, most of these recent designs have not been synthesized, expressed or tested in animal models to validate their predictions across all metrics used in the design phase. This lack of translation is unsurprising, the availability of robust and free in silico tools has only occurred in the past couple of years, coupled with the fact that S. mansoni is a neglected tropical disease with funding and support for translational research scarce.
Murine strain selection and discrimination
Across inbred mice, immune responses to S. mansoni antigens are strain dependent and antigen class specific. Comparative analysis of SmTPI-specific lymphokine responses have shown strain dependant immune variation with C57BL/6 demonstrating a Th1-biased response during in vitro restimulation of mesenteric lymph node cells from 7–8 week S. mansoni-infected mice showed SmTPI-specific lymphokine responses with elevated IL-2 and low level IFN-γ (Reynolds et al., Reference Reynolds, Dahl and Harn1994). In contrast, CBA/J mice exhibited a stronger SmTPI-specific Th1-type recall response than C57BL/6J mice, with mesenteric lymph node cells from infected animals producing substantially higher IL-2 and IFN-γ after in vitro SmTPI stimulation, while IL-4 and IL-5 remained undetectable (Reynolds et al., Reference Reynolds, Dahl and Harn1994). Compared with CBA/J mice, C3H/J mice showed a weaker SmTPI-specific Th1-type response, with no detectable IL-2, IL-4 or IL-5 and only low IFN-γ production by mesenteric lymph node cells after in vitro SmTPI stimulation (Reynolds et al., Reference Reynolds, Dahl and Harn1994). Epitope mapping between the strains showed recognition of different peptides and illustrated MHC-linked epitope selection. Studies into Sm23 have also found strain dependent immune response with T-cell recognition differing between C57BL/6J, CBA/J and BALB/c strains only sharing a single recognized epitope (Reynolds et al., Reference Reynolds, Shoemaker and Harn1992). The immune response differences between strains remains a gap in S. mansoni vaccine research and requires more studies to reach a consensus on the optimal murine model strain. Another important translational challenge is that standard laboratory strains of mice are highly inbred, whereas human populations are outbred and exhibit considerable genetic diversity, which can influence epitope recognition and vaccine responsiveness (Medetgul-Ernar and Davis, Reference Medetgul-Ernar and Davis2022). This genetic difference may contribute to the difficulty of replicating promising preclinical findings in human trials.
Standardization of the routes of infection in mice with cercariae between experiments has not been established (Houlder et al., Reference Houlder, da Silva, van Diepen, Amaral, Wilson, Hokke, Roestenberg and Bakker2025), with methods for tail exposure/immersion, percutaneous abdominal exposure using ring-based methods (Tucker et al., Reference Tucker, Karunaratne, Lewis, Freitas and Liang2013; Zhang et al., Reference Zhang, Molehin, Patel, Kim, Peña, Siddiqui and Timson2020b), and, in some cases, artificial subcutaneous inoculation of cercariae or schistosomula (Lombardo et al., Reference Lombardo, Pasche, Panic, Endriss and Keiser2019). The choice of infection route significantly influences the survival rates of cercariae and schistosomula. Vilar and Pinto (Reference Vilar and Pinto2005) showed that bypassing the skin phase by subcutaneous inoculation enhanced the recovery of adult worms relative to percutaneous abdominal infection, indicating that artificial routes may affect efficacy estimates and diminish the strength of comparisons to natural percutaneous exposure methods. Tendler et al. (Reference Tendler, Magalhães Pinto, Côrtes and Gebara1985) showed that both infection route and anatomical site of cercarial exposure influence S. mansoni infection efficiency in mice, noting that the percutaneous route of inoculation is preferential as it mimics natural infection and that skin exposure to S. mansoni may be an important trigger of host mechanism of immunity. This distinction is more than methodological preference as the skin is itself a major immune barrier and immune-inductive site which encounters cercariae and early schistosomula antigens, with Langerhans cells been shown to migrate from the epidermis to skin draining lymph nodes where the adaptive immune response is initiated (Mountford and Trottein, Reference Mountford and Trottein2004). Furthermore, per animal cercarial dosage used in infection experiments also lacks standardization, with cercarial dosage ranges proposed as 25–30 for chronic infection models, 100–150 for vaccine testing and 180–200 for maximum egg and adult worm recovery (Tucker et al., Reference Tucker, Karunaratne, Lewis, Freitas and Liang2013). Collectively, variation in both the infection route and challenge dosage adds complexity when comparing vaccine efficacy across S. mansoni studies.
The translation of protective efficacy from murine S. mansoni vaccine models to human protection remains a major unresolved challenge owing to substantial heterogeneity in animal models, limited predictive value for primate or human efficacy, the absence of validated correlates of protection and the inconsistent clinical performance of candidates with promising preclinical protective profiles (Panzner et al., Reference Panzner, Excler, Kim, Marks, Carter and Siddiqui2021). Further research is required to improve the predictive value of murine infection models, which would greatly enhance the discrimination of correlative protection factors and efficacy predictions in humans.
Concluding remarks
Previous decades of praziquantel MDA programmes have failed to reduce the global burden of S. mansoni induced schistosomiasis. This failure can be attributed to praziquantel’s limited bioavailability and narrow activity against mature S. mansoni worms, which allows for rapid reinfection, underscoring the crucial need for a vaccine. Historical vaccine attempts have failed to induce the required protection in either animal or human trials, and it remains critically important to continue research into a vaccine as improvements in efficacy, identified epitope targets, adjuvants and the possibility of pan-species protection would represent noteworthy progress towards a vaccine for S. mansoni. Furthermore, S. mansoni is a complex parasite that has likely co-evolved with humans to effectively evade the immune system, and there is no guarantee that any initially successful vaccine will not result in selective pressure, causing S. mansoni to once again adapt to overcome the vaccine. Therefore, continued investigation of diverse antigens and development of vaccines is crucial in mitigating such risks.
Advances in vaccine design due to the advent of in silico models allow for rapid identification of novel targets from large genomic datasets, and the design, refinement, structural analysis and simulation of immunization with improved accuracy and speed should be utilized in the development of novel vaccines for S. mansoni, reducing the cost of research while also increasing the speed of initial development. It is equally important that in silico work be translated into real-world experiments to validate predictions and contribute to the data to further inform and enhance current and future work in the pursuit of a truly effective vaccine for S. mansoni.
Future vaccine research should prioritize an iterative pipeline in which in silico designed and ranked candidates are experimentally tested for expression, antigenicity, immunogenicity and protective efficacy in a standardized methodology. This approach would align early-stage antigen discovery with translational vaccine development and may accelerate progress towards an effective vaccine for S. mansoni.
Author contributions
A.T. prepared the original manuscript draft and figures. All authors edited the manuscript and approved the final version.
Financial support
A.T. acknowledges funding support from the Rural Health Research Institute, Charles Sturt University.
Competing interests
The authors declare no competing interests.
Ethical standards
Not applicable.

 Open access
Open access