



Introduction
Bats (Chiroptera) represent the second most diverse order of mammals after rodents, constituting approximately 22% of all mammalian species, with around 1420 species documented globally (Burgin et al., Reference Burgin, Colella, Kahn and Upham2018; Letko et al., Reference Letko, Seifert, Olival, Plowright and Munster2020). They inhabit all continents except Antarctica (Letko et al., Reference Letko, Seifert, Olival, Plowright and Munster2020), with particularly high densities noted in Mediterranean regions, including Southern Europe and North Africa (Rose, Reference Rose2006; Aulagnier et al., Reference Aulagnier, Haffner, Mitchell-Jones and Moutou2017). While bats have been associated with various disease outbreaks, their ecological importance is significant, as they fulfil critical roles in ecosystems, such as regulating arthropod populations and facilitating plant pollination (Aggrey et al., Reference Aggrey, Rwego, Sande, Khayiyi, Kityo, Masembe and Kading2024).
Despite their ecological benefits, bats are recognized as natural reservoirs for several zoonotic pathogens, including viral, bacterial, parasitic and fungal agents (Smith and Wang, Reference Smith and Wang2012; Allocati et al., Reference Allocati, Petrucci, Di Giovanni, Masulli, Di Ilio and De Laurenzi2016). These pathogens can be disseminated over considerable distances, aided by the migratory behaviour of certain bat species (Messenger et al., Reference Messenger, Rupprecht, Smith, Kunz and Fenton2003). Algeria, with its favourable ecological conditions (Yeddou and Lammari, Reference Yeddou and Lammari2017), hosts a remarkable diversity of bat species, ranking second in Africa after Sudan. The bat fauna of Algeria includes 25 species across 7 families, with 6 species from the Rhinolophidae family and 13 species from the Vespertilionidae family (Ahmim, Reference Ahmim2013, Reference Ahmim2017, Reference Ahmim2019).
Research on chiropteran parasites, particularly ectoparasites and endoparasites, has garnered increased interest in recent years due to their relevance in modelling the emergence and transmission of diseases (Dick and Patterson, Reference Dick and Patterson2007; Wibbelt et al., Reference Wibbelt, Moore, Schountz and Voigt2010; Attia-Hili, Reference Attia-Hili2015). However, studies on bat parasites in Algeria remain limited and fragmented (Bendjeddou et al., Reference Bendjeddou, Khelfaoui, Abiadh, Mechouk, Mihalca and Sándor2024). Notable efforts include investigations into the seasonal infestation of Myotis punicus Felten, 1977 by Ixodes vespertilionis Koch, 1844 (Bendjeddou et al., Reference Bendjeddou, Bouslama, Amr and Bani Hani2016) and inventories of bat ectoparasites (Ter Hofstede and Fenton, Reference Ter Hofstede and Fenton2005; Bendjeddou et al., Reference Bendjeddou, Bitam, Abiadh, Bouslama and Amr2013, Reference Bendjeddou, Bouslama, Amr and Bani Hani2016, Reference Bendjeddou, Loumassine, Scheffler, Bouslama and Amr2017).
Bats are known to host a wide variety of ectoparasites (Anciaux de Faveaux, Reference Anciaux de Faveaux1965, Reference Anciaux de Faveaux1976; Amr and Qumsiyeh, Reference Amr and Qumsiyeh1993; Stanyukovich, Reference Stanyukovich1997; Imaz et al., Reference Imaz, Aihartza and Totorika1999; Bitam et al., Reference Bitam, Dittmar, Parola, Whiting and Raoult2010; Camilotti et al., Reference Camilotti, Graciolli, Weber, Arruda and Cáceres2010; Orlova, Reference Orlova2011, Reference Orlova2013; Tsai et al., Reference Tsai, Chang, Chuang and Chomel2011; Krištofík and Danko, Reference Krištofík and Danko2012; Morse et al., Reference Morse, Olival, Kosoy, Billeter, Patterson, Dick and Dittmar2012; Klimpel and Mehlhorn, Reference Klimpel and Mehlhorn2014; Morand, Reference Morand2014; Burazerović et al., Reference Burazerović, Ćakić, Mihaljica, Sukara, Ćirović and Tomanović2015; Sponchiado et al., Reference Sponchiado, Melo, Martins, Krawczak, Labruna and Căcere2015; Leulmi et al., Reference Leulmi, Aouadi, Bitam, Bessas, Benakhla, Raoult and Parola2016; Khelfaoui et al., Reference Khelfaoui, Kebaci and Benyacoub2018). In Algeria’s Chrea National Park (CNP), 2 significant studies on bat ectoparasites were conducted in the El Hamdania sector (Yeddou and Lammari, Reference Yeddou and Lammari2017; Bendjoudi et al., Reference Bendjoudi, Yedou, Beneldjouzi, Mechouk and Bendjeddou2019). In addition to ectoparasites, bats harbour various endoparasites, including cestodes, trematodes and nematodes (Theodorides, Reference Theodorides1953; Combes and Clerc, Reference Combes and Clerc1970; Gardner and Jiménez-Ruiz, Reference Gardner and Jiménez-Ruiz2009; Horvat et al., Reference Horvat, Čabrilo, Paunović, Karapandža, Jovanović, Budinski and Čabrilo2015; Singh et al., Reference Singh, Barolia and Sharma2015; Hadjouti and Mamouni, Reference Hadjouti and Mamouni2016; Webber and Willis, Reference Webber, Willis and Ortega2016; Léger, Reference Léger2019). However, knowledge regarding the endoparasites of Algerian bats remains limited.
This study aimed to document the ectoparasites and endoparasites associated with bats in CNP, thereby contributing valuable insights into the parasitic fauna of Algerian bats and their ecological interactions. Specifically, we sought to: (i) identify the bat species present in tunnels of CNP and confirm potential new records for the park and the region; (ii) inventory the ectoparasitic Diptera and Acari species colonizing bat hosts and assess their relative abundance (RA) across different bat species; (iii) conduct coprological analyses of guano samples to identify helminth and arthropod endoparasites, determine their prevalence and quantify their RA; (iv) evaluate whether ectoparasite composition is associated with host species using statistical methods; and (v) provide the first baseline data for ectoparasite–host associations in CNP, thereby establishing a foundation for future faunistic, epidemiological and ecological studies on the role of bats in regional biodiversity and potential pathogen transmission.
Materials and methods
Study area
The CNP, located between latitudes 36°19′ N–36°30′ N and longitudes 02°38′ E–03°02′ E (Figure 1), encompasses an area of 26 587 hectares (BNEF, 1984). The park is recognized for its rich biodiversity, hosting a diverse range of flora and fauna, including a notable variety of chiropteran species. In the western region of the park, bats have been observed inhabiting disused railway tunnels and abandoned mines (Figure 2) along the Chiffa River (36°24′0.01″ N, 2°46′0.01″ E; elevation: 239 m a.s.l.) (Meddour, Reference Meddour2002). CNP is predominantly characterized by montane forests and shrublands, dominated by Atlas cedar (Cedrus atlantica (Endl.) Manetti ex Carrière), holm oak (Quercus ilex L.) and mixed broadleaved woodlands. The main woodland areas are distributed across Oued El Marja, Tibenkent, Beni-Selmen, Tamesguida and the central Chréa region (Bendjoudi et al., Reference Bendjoudi, Marniche, Takdjout, Epalanga and Chenchouni2023).
Vegetation map and geographic location of Chrea National Park in Algeria.

Figure 1 Long description
The map illustrates the vegetation distribution within Chrea National Park, located in Algeria. The park is highlighted in green on the inset map, showing its position relative to nearby cities such as Blida and Medea. The main map details various vegetation types, with a legend indicating categories: cultivated lands, Quercus suber forests, Quercus ilex forests, Cedrus atlantica forests, Pinus halepensis forests and herbaceous vegetation. The Chiffa Tunnel is marked in solid red circle. The distribution of vegetation types is shown across different regions of the park, with cultivated lands and various forest types spread throughout, indicating the park's diverse ecological zones.
Photos of Chiffa Tunnel at the Chrea National Park in Algeria. Left: outside the tunnel, right: inside the tunnel, near the entrance.

Figure 2 Long description
The left image shows two persons walking on a narrow bridge made of metal beams, situated over a river witin a rocky terrain outside the Chiffa Tunnel. The background includes a mine road. The right image depicts the interior of the Chiffa Tunnel, featuring railway tracks leading into the darkness of the tunnel. The tunnel walls are made of stone.
Bat sampling
Field sampling was conducted between May and June 2019 in the El Hamdania tunnel (Figure 2). Bats were captured using hand nets (Figure 3), a method compliant Algerian Law and preferred in chiropteran research (Kunz and Kurta, Reference Kunz, Kurta and Kunz1988; Mitchell-Jones and Robertson, Reference Mitchell-Jones, Robertson, Mitchell-Jones and McLeish2004). Captured bats were placed in thick, aerated capture bags and transported to the Zoology Laboratory of the National Veterinary School of El Alia (Algiers) for further examination.
Capture bats with a hand net.

Figure 3 Long description
The left image shows a close-up of a bat in a tunnel, highlighted by a circle and an arrow pointing to it. The right image depicts a person (a technician-forester from the CNP) using a hand net to capture bats perched on the tunnel ceiling. The person is positioned at the bottom, holding the net upward toward the bats in the ceiling, which are indicated by an arrow and label.
Bat species identification was conducted using the determination keys of Dietz and von Helversen (Reference Dietz and von Helversen2004) and Dietz (Reference Dietz2005), by Prof Faiza Marniche at National Veterinary School of El Alia (Algeria) and Prof Djamel Bendjoudi at the Laboratory of Biotechnology, Environment, and Health in Blida 1 University. Following identification and examination, all bats were released back into their natural habitats by the end of the same day.
Collection and identification of endoparasites in guanos
For parasitological analysis, 150 g of guano was collected from each individual of 2 Rhinolophidae species (Rhinolophus blasii and Rhinolophus ferrumequinum). Parasites were identified using the flotation technique (Sivajothi et al., Reference Sivajothi, Reddy and Rayulu2014; Hendrix and Robinson, Reference Hendrix and Robinson2023) and the direct smear method. Microscopic examination was conducted at magnifications of 10×, 40× and 100×, with parasite identification guided by standard taxonomic keys (Sonin, Reference Sonin1975; Bray et al., Reference Bray, Gibson and Jones2008; Anderson et al., Reference Anderson, Chabaud and Willmott2009; Léger, Reference Léger2019; Zajac et al., Reference Zajac, Conboy, Little and Reichard2021).
Collection and identification of ectoparasites on bats
Each bat was individually examined for ectoparasites, which were carefully removed with fine forceps following macroscopic examination of all body regions (Hutson et al., Reference Hutson, Mickleburgh and Racey2001). Collected ectoparasites were preserved in 70% ethanol and processed through a series of preparatory steps: immersion in a 10% potassium hydroxide (KOH) solution for 24 h, rinsing in distilled water for 30 min and subsequent dehydration in 100% ethanol. Prepared specimens were mounted on slides and examined under a Leica DM500 optical microscope at 100× magnification.
Ectoparasite identification was performed using established taxonomic references (Arthur, Reference Arthur1956; Hopkins and Rothschild, Reference Hopkins and Rothschild1956; Rudnick, Reference Rudnick1960; Dusbabek, Reference Dusbabek1962; Radovsky, Reference Radovsky1967; Theodor, Reference Theodor1967; Hůrka, Reference Hůrka1982; Dietz and von Helversen, Reference Dietz and von Helversen2004). Additional verification was provided by Prof Faiza Marniche at the National Veterinary School (Algiers, Algeria) and Prof Carl Dick (Department of Biology, Western Kentucky University, Kentucky, USA), utilizing dichotomous keys.
Data analysis
The results obtained from our study on ectoparasites and endoparasites were analysed using the positivity index applied to ectoparasite samples collected from bats and guano. The positivity index represents the percentage of samples containing a parasite, calculated as P% = (P+ /Pt) × 100, where P% is the positivity index, P+ is the number of positive samples and Pt is the total number of samples (El Guamri et al., Reference El Guamri, Belghity, Achicha, Tiabi, Aujjar and Barkia2009). Parasite abundance (A) is defined as the ratio of the total number of individuals of a parasite species (n) to the total number of individuals host examined (H) (Margolis et al., Reference Margolis, Esch, Holmes, Kuris and Schad1982). In this context, H denotes the number of host samples examined, and A is calculated as A = n/H, where n represents the number of individuals of the parasite species. Finally, statistical analyses were conducted using the software Quantitative Parasitology V 3.0 (Rózsa et al., Reference Rózsa, Reiczigel and Majoros2000) to process prevalence and other relevant parameters. The package {ggalluvial} version 0.12.5 under the statistical environment R was used to generate to generate alluvial diagram depicting the distribution of dipteran ectoparasites across their bat hosts.
Results
Bat species
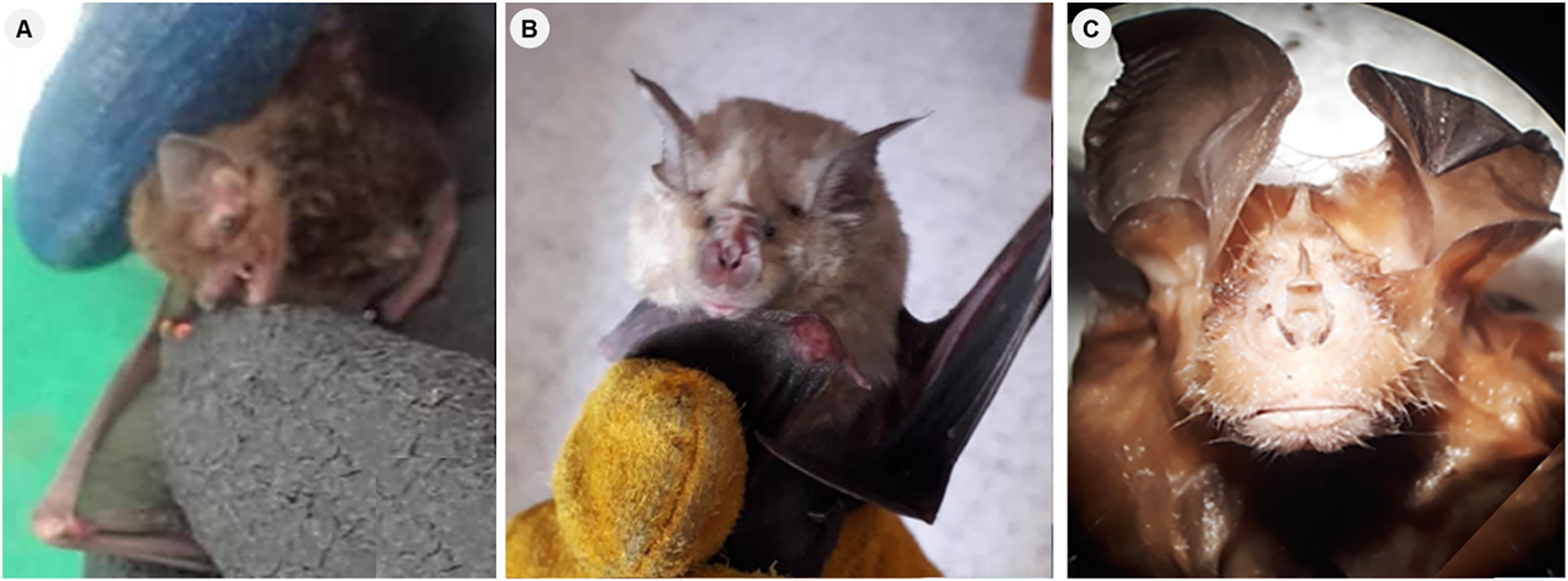
During field sampling events around the El Hamdania tunnels in April and June 2019, we collected 20 bats from 2 different families: 2 species from the Rhinolophidae family and 1 species from the Vespertilionidae family. The captured bat species included Blasius’s Horseshoe Bat (Rhinolophus blasiiblasii Peters, 1866) with 18 individuals (13 males and 5 females), the Greater Horseshoe Bat (Rhinolophus ferrumequinum (Schreber, 1774)) which comprised 1 male and Leisler’s Bat (Nyctalus leisleri (Kühl, 1817)) also represented by 1 male (Figure 4). It is noteworthy that N. leisleri (Vespertilionidae) is a new species that has not been previously reported in the CNP.
Bat species found in the Chiffa tunnel around the Chrea National Park. (A) Leisler’s Noctule Nyctalus leisleri, (B) Blasius’s horseshoe bat Rhinolophus blasii, and (C) greater horseshoe bat Rhinolophus ferrumequinum.

Figure 4 Long description
A composite photograph with three separate photographs labeled A, B and C. A: An individual of Leisler’s Noctule bat held in hand, with two gloved fingers visible wearing a rubber glove. B: Frontal view of an individual Blasius’s horseshoe bat held in a hand wearing a yellow leather glove, with the bat’s wings partly spread. C: Frontal view of an individual Greater horseshoe bat with both ears spread around the face. Visible fur covers the head, while part of a wing appears in the background.
Ectoparasites
The identification of ectoparasites found in the 2 species of Rhinolophidae revealed 30 ectoparasites were collected from 18 R. blasii (14 males and 4 females) and 1 R. ferrumequinum (male) between April and June 2019. We noted the presence of a single class, a single order with 2 families, namely 20 Streblidae flies including Brachytarsina (Brachytarsina) flavipennis Macquart, 1851 [= Nycteribosca africana] and 10 Nycteribiidae flies including Phthiridium biarticulatum Hermann, 1804 (n = 5 flies) and Nycteribia schmidlii Schiner, 1853 (n = 5 flies) (Figure 5).
Microscopic views (40× magnification) of Nycteribia (Listropoda) schmidlii schmidlii Schiner, 1853 (Diptera: Nycteribiidae): (A) female ventral view; (B) male ventral view; (C) male ventral extremity; (D) dorsal view of Nycteribosca africana = Brachytarsina (Brachytarsina) flavipennis Macquart, 1851 (Diptera: Streblidae) and (E) ventral view of Phthiridium biarticulatum (Diptera: Nycteribiidae) Hermann, 1804.

Figure 5 Long description
Five microscope photographs are arranged in a composite figure and labeled A, B, C, D and E. A shows a ventral view of a female of Nycteribia (Listropoda) schmidlii schmidlii. B shows a ventral view of a male of Nycteribia (Listropoda) schmidlii schmidlii. C shows a close-up of the posterior end of a male of Nycteribia (Listropoda) schmidlii schmidlii. D shows a dorsal view of Nycteribosca africana = Brachytarsina (Brachytarsina) flavipennis. E shows a ventral view of Phthiridium biarticulatum. A measurement scale with tick marks is present along the lower edge of the frame.
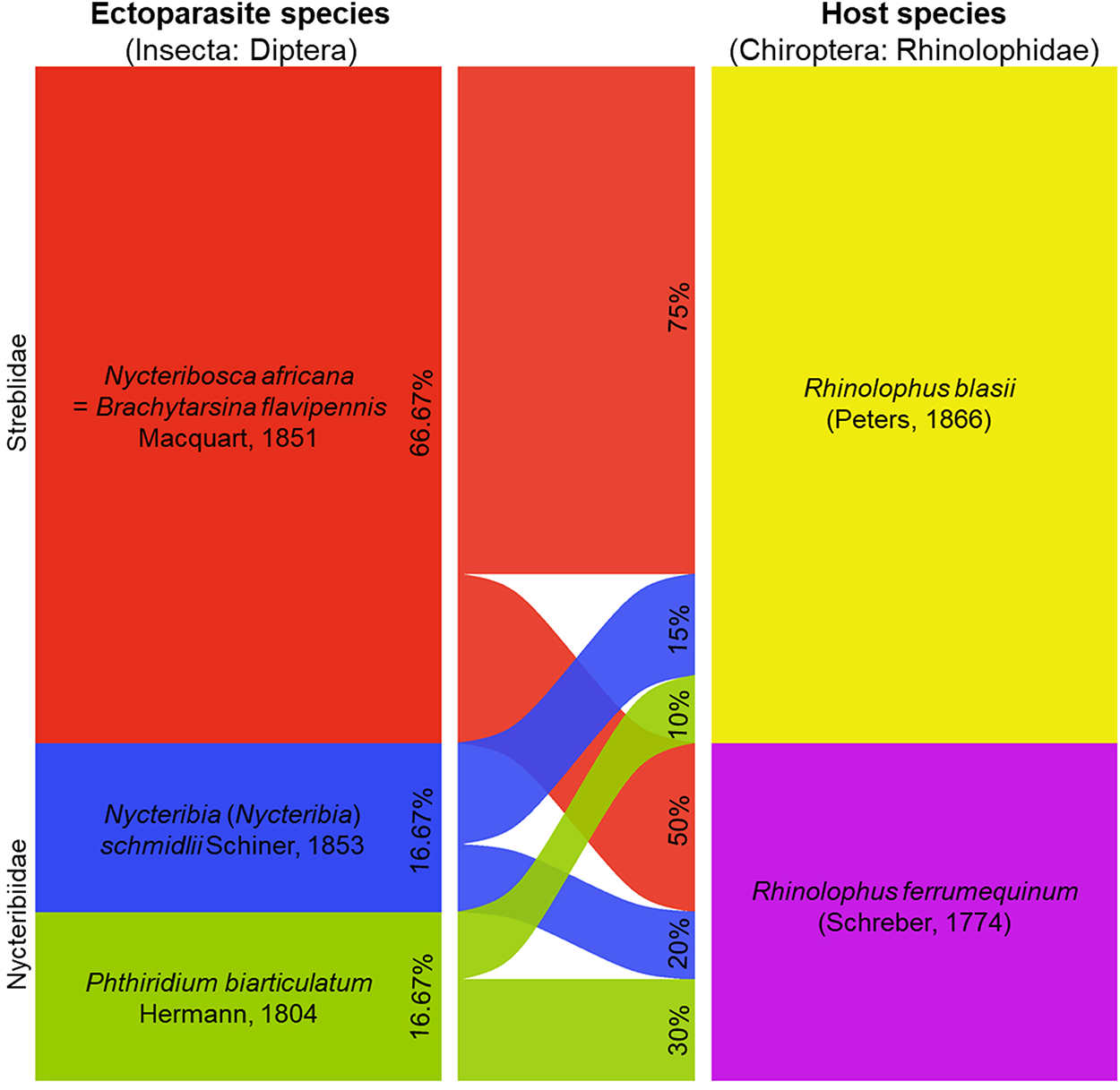
These ectoparasites were collected from the patagium of R. blasii females and R. ferrumequinum male. The ectoparasite community associated with the captured bats showed varying levels of RA among host species. In R. blasii, Nycteribosca africana was the dominant parasite (75%), followed by N. schmidlii (15%) and Phthiridium biarticulatum (10%). In R. ferrumequinum, N. africana also prevailed but at a lower proportion (50%), while P. biarticulatum (30%) and N. schmidlii (20%) were relatively more common compared to their occurrence in R. blasii (Figure 6). The Pearson chi-square test of independence revealed no significant association between bat host species and ectoparasite composition (χ 2 = 2.00, df = 2, p = 0.37), indicating that ectoparasite distribution was relatively similar between R. blasii and R. ferrumequinum.
Alluvial plot illustrating the distribution of dipteran ectoparasites across their bat hosts. The proportions (%) are relative abundances.

Figure 6 Long description
The alluvial plot illustrates the distribution of dipteran ectoparasites among bat host species of the Rhinolophidae family. On the left, ectoparasite species are listed: Nycteribosca africana Macquart, 1851 with 66.67% in red, Nycteribia schmidlii Schiner, 1853 with 16.67% in blue and Phthiridium biarticulatum Hermann, 1804 with 16.67% in green. On the right, bat host species are shown: Rhinolophus blasii (Peters, 1866) and Rhinolophus ferrumequinum (Schreber, 1774). The plot shows the flow of relative abundances between ectoparasite and host species, indicating the proportions of each ectoparasite species associated with each host species.
Endoparasites
Coprological analysis results
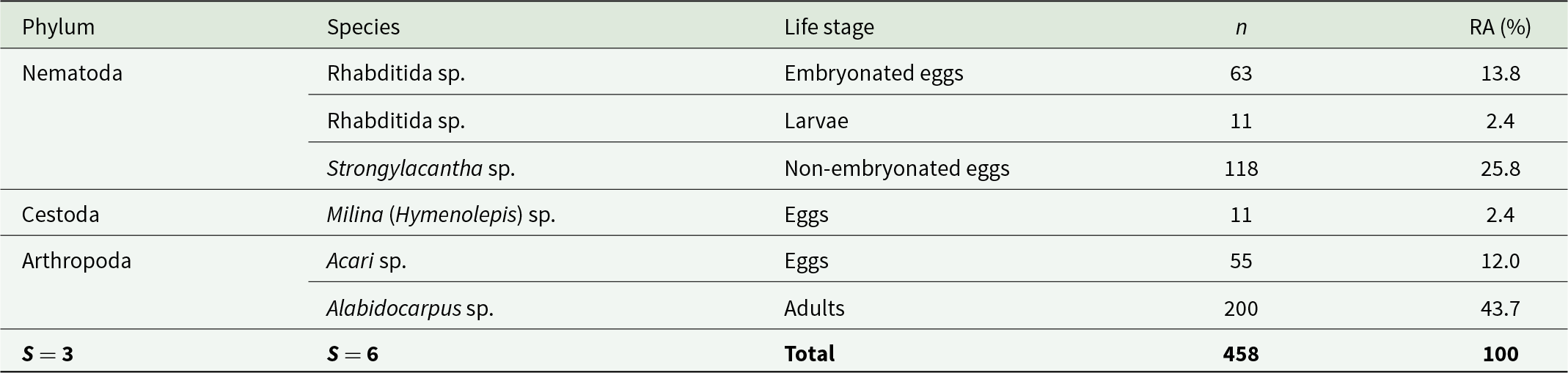
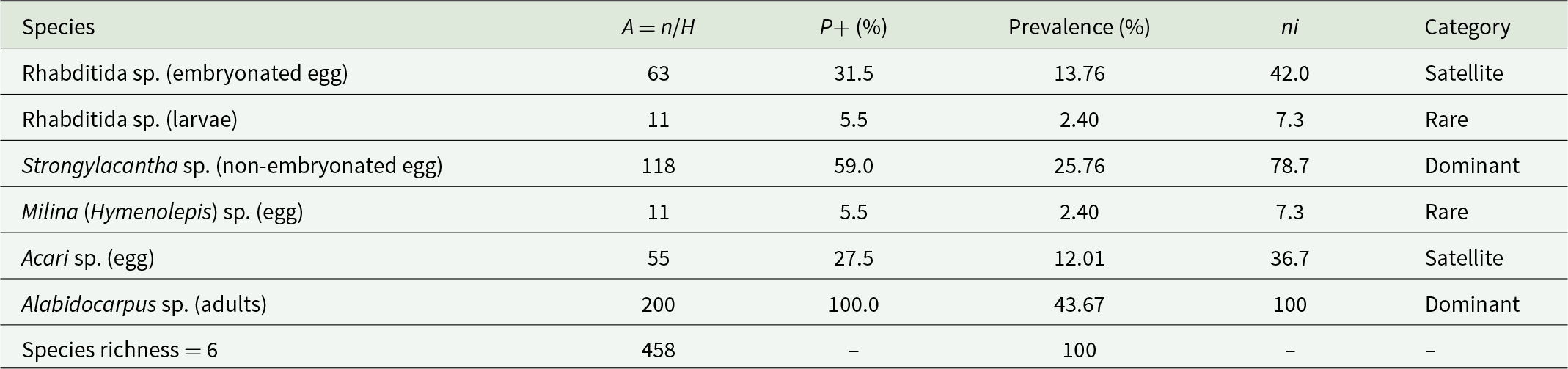
A study of the parasitic fauna of the bats revealed the presence of a species of cestode and 2 species of nematodes within 150 g of guano samples (Table 1). A substantial presence of arthropods was also recorded, represented by adult mites and eggs (n = 255) (Figure 7). We observed a high RA for the arthropod phylum (55.7%). The species Alabidocarpus sp. (adults) dominated with a rate of 43.7%. The nematode Strongylacantha sp. (non-embryonated eggs) ranked second with 118 individuals (RA = 25.8%), followed by Acari sp. (eggs) with 55 individuals (RA = 12.0%). Rhabditida sp. (embryonated eggs) was observed with an RA of 13.8% (63 individuals), while the cestode Milina (Hymenolepis) sp. (eggs) was represented by 11 individuals (RA = 2.4%) (Table 1).
Inventory and abundance of parasites found in bat guanos studied in Chrea National Park

Table 1 Long description
The table reports parasite taxa detected in bat guano samples, listing each taxon’s phylum, life stage, number of individuals counted per 150 g of guano, and relative abundance (RA in %). A total of 458 individuals were recorded across 6 species within 3 phyla. Arthropods were most abundant overall, driven by Alabidocarpus sp. adults with 200 individuals (43.7%) and Acari sp. eggs with 55 (12.0%). Nematodes accounted for three entries: Strongylacantha sp. non-embryonated eggs were the second most common item at 118 (25.8%), while Rhabditida sp. appeared as embryonated eggs (63; 13.8%) and larvae (11; 2.4%). The only cestode listed, Milina (Hymenolepis) sp. eggs, was rare at 11 individuals (2.4%). Percentages sum to 100% and reflect relative abundance within the total count, not prevalence across bats or sites.
S = Total richness, n = number of individuals (per 150 g guanos), RA = relative abundance in %.
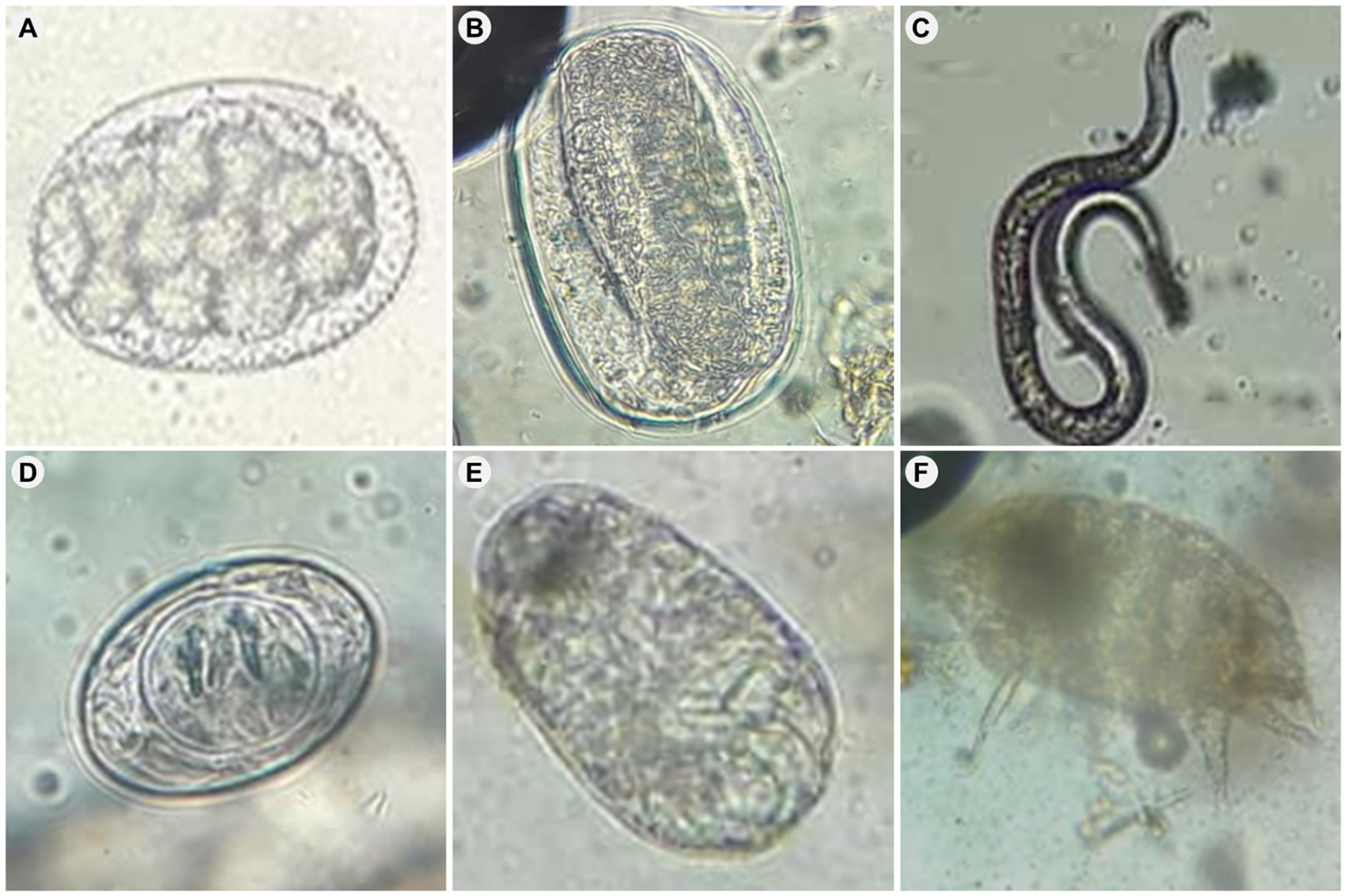
Parasites found in bat guanos observed under light microscope at 40× magnification. (A) Non-embryonated egg of Strongylacantha sp.; (B) embryonated egg of Rhabditida sp.; (C) larva Rhabditida sp.; (D) egg of Milina (Hymenolepis) sp.; (E) egg Acari and (F) adult Alabidocarpus sp.

Figure 7 Long description
The image A showing a non-embryonated egg Strongylacantha sp.. The image B showing an embryonated egg of Rhabditida sp. The image C showing a larva of Rhabditida sp. The image D showing an egg of Milina (Hymenolepis) sp. The image E showing an egg of Acari. The image F showing an adult of Alabidocarpus sp.
Prevalence of endoparasites
The prevalence of endoparasites found in the guanos of the Rhinolophidae studied is presented in Table 2. Out of a total of 150 g of Rhinolophidae guano, a prevalence of 100% was infested by Alabidocarpus sp. (adults), followed by Strongylacantha sp. (non-embryonated eggs) with a prevalence rate of 78.7%. Rhabditida sp. (embryonated eggs) has a prevalence rate of 42.0%, while Acari sp. (eggs) shows a prevalence rate of 36.7%. Both Rhabditida sp. (larvae) and Milina (Hymenolepis) sp. (eggs) represent a prevalence rate of 7.3% each. We also note the presence of 2 dominant species: Alabidocarpus sp. (adults) and Strongylacantha sp. (non-embryonated eggs). Additionally, there are satellite species, including Rhabditida sp. (embryonated eggs) and Acari sp. (eggs), as well as rare species such as Rhabditida sp. (larvae) and Milina (Hymenolepis) sp. (eggs) (Table 2).
Abundance (A), positivity index (P+) and prevalence of parasites found in bat guanos studied in Chrea National Park (Algeria)

Table 2 Long description
The table summarizes six parasite taxa detected in bat guano, reporting sample size (ni), parasite abundance (A), positivity index (P+), parasite prevalence, and a frequency category. Alabidocarpus sp. (adults) is the most prominent taxon, with ni 200, abundance 100.0, P+ 43.67%, and 100% prevalence, categorized as dominant. Strongylacantha sp. (non-embryonated eggs) is also dominant (ni = 118; A = 59.0; P+ = 25.76%; prevalence = 78.7%). Rhabditida sp. embryonated eggs and Acari sp. eggs are intermediate (“satellite”) with prevalence 42.0% and 36.7% and P+ 13.76% and 12.01%, respectively. The least frequent (“rare”) taxa are Rhabditida larvae and Milina (Hymenolepis) eggs, each with ni = 11, A = 5.5, P+ = 2.40%, and prevalence = 7.3%. Overall species richness is 6, total ni is 458, and P+ sums to 100%, so comparisons should focus on relative differences across taxa rather than interpreting P+ as an independent prevalence measure.
n = number of individuals, A = parasite abundance, H = number of parasitized hosts.
Discussion
The present study provides the first survey of bat ectoparasites, with supplementary coprological data, in CNP, Algeria, thereby filling a significant gap in the regional knowledge of chiropteran ecology. Three bat species were recorded, including R. blasii and R. ferrumequinum, with N. leisleri reported for the first time in the park, marking an extension of its known distribution in North Africa. Parasitological analyses revealed a remarkable diversity of metazoan parasites, encompassing both ectoparasites (Diptera and Acari) and endoparasites (Nematoda and Cestoda). Among the most noteworthy findings, N. schmidlii parasitizing R. blasii was reported for the first time in Algeria, as well as a distinct composition of helminth communities, which varied across host species. These results underscore not only the ecological complexity of bat-parasite interactions in this understudied region but also the originality of the faunistic data presented here, which contribute novel records at both national and local levels.
Bat species
Leisler’s Noctule (N. leisleri) is a migratory species found in Europe and has been documented in the northern Himalayan region of Pakistan (Walker et al., Reference Walker, Warnick, Lange, Uible, Hamlet, Davis and Wright1964). It is recognized as a Mediterranean species and appears to be the most prevalent Nyctalus species in Africa. This species has been recorded only in Yakouren, located in northern Algeria (Tizi Ouzou), as noted by Hanak and Gaisler (Reference Hanak and Gaisler1983), Gaisler (Reference Gaisler1983) and Gaisler and Kowalski (Reference Gaisler and Kowalski1986). Hanak and Gaisler conducted a biometry study of the 4 individuals discovered in 1983 in Northern Cyrenaica, Libya and Morocco (El Ibrahimi and Idrissi, Reference El Ibrahimi and Idrissi2015), as well as in Bekrite in the Middle Atlas (Benda et al., Reference Benda, Ruedi and Aulagnier2004, Reference Benda, Červený, Konečný, Reiter, Ševčík, Uhrin and Vallo2010). N. leisleri was first recorded in CNP. While its current distribution and demographic data are not available, its presence has been confirmed. This locality represents the southernmost point for this species in the Maghreb and is also the most continental site in Africa.
Ectoparasites
Bats host a diverse array of ectoparasites and serve as crucial food sources for numerous haematophagous arthropods. These ectoparasites can negatively impact the fitness of their bat hosts, affecting both survival and reproductive success. Such fitness costs drive the evolution of host behavioural defences to mitigate parasitism (Ter Hofstede and Fenton, Reference Ter Hofstede and Fenton2005). In Algeria, studies on bat ectoparasites remain relatively limited. Our findings concerning ectoparasites from the Nycteribiidae family align with the work of Bendjeddou et al. (Reference Bendjeddou, Loumassine, Scheffler, Bouslama and Amr2017), who compiled a systematic list of bat ectoparasites in Algeria, documenting 25 species, including 3 bat fly species from the Nycteribiidae family.
The bat fly N. schmidlii exhibits a broad distribution, spanning southern Europe, North Africa, southwestern Asia and parts of Central Asia, including Afghanistan (Hůrka, Reference Hůrka1964). In Algeria, this species has been reported on various bat hosts, including Miniopterus schreibersii and Rhinolophus mehelyi (Falkoz, 1923), R. ferrumequinum (Theodor and Moscona, Reference Theodor and Moscona1954), Myotis blythi (Rotrou, Reference Rotrou1939; Aellen, Reference Aellen1955), M. schreibersii, M. blythi and R. euryale (Amr and Qumsiyeh, Reference Amr and Qumsiyeh1993). Elsewhere, N. schmidlii has been documented on Asellia tridens in Palestine (Theodor and Moscona, Reference Theodor and Moscona1954) and on M. schreibersii in Turkey (Albayrak, Reference Albayrak2003). Hůrka (Reference Hůrka1964) identified M. schreibersii as the principal host of N. schmidlii, while also listing secondary hosts such as M. blythi, M. capaccinii, M. emarginatus, Myotis myotis, Rhinolophus euryale, R. ferrumequinum and R. mehelyi. These bats are typically social, and residing in caves (Scheffler et al., Reference Scheffler, Dolch, Ariunbold, Stubbe, Abraham and Thiele2012). Notably, this study reports N. schmidlii schmidlii parasitizing R. blasii in Algeria for the first time.
Phthiridium biarticulatum is a circum-Mediterranean species associated primarily with cave-dwelling bats. It demonstrates a strong preference for Rhinolophus species (Ševčík et al., Reference Ševčík, Benda and Lucan2013), particularly R. ferrumequinum and R. euryale (Beaucournu and Noblet, Reference Beaucournu and Noblet1996). In this study, this bat fly was collected from R. blasii and R. ferrumequinum. In Algeria, P. biarticulatum has been reported on R. ferrumequinum (Theodor and Moscona, Reference Theodor and Moscona1954) and R. blasii (Amr and Qumsiyeh, Reference Amr and Qumsiyeh1993; Bendjeddou et al., Reference Bendjeddou, Bitam, Abiadh, Bouslama and Amr2013, Reference Bendjeddou, Loumassine, Scheffler, Bouslama and Amr2017). Beyond Algeria, it has been recorded on R. mehelyi in Libya (Hůrka, Reference Hůrka1982; Amr and Qumsiyeh, Reference Amr and Qumsiyeh1993), R. blasii, R. euryale and R. ferrumequinum in Iran (Nader and Kock, Reference Nader and Kock1983; Benda et al., Reference Benda, Spitzenberger, Hanák, Andreas, Reiter, Ševčík, Šmíd and Uhrin2014), and R. blasii, R. clivosus, R. euryale and R. ferrumequinum in Jordan (Benda et al., Reference Benda, Červený, Konečný, Reiter, Ševčík, Uhrin and Vallo2010).
Nycteribosca africana is the sole representative of the Streblidae family in Western Europe. This small-winged, thermophilic fly is predominantly associated with troglophilic and gregarious bats of the genera Myotis and Rhinolophus (Beaucournu and Noblet, Reference Beaucournu and Noblet1996). Previously referred to as Nycteribosca kollari, this species thrives along winter isotherms of +8 to +10 °C (Hůrka, Reference Hůrka1962). Although primarily parasitizing Mediterranean Rhinolophus species, N. africana is also common on Myotis species (Hůrka, Reference Hůrka1962; Scheffler et al., Reference Scheffler, Dolch, Ariunbold, Stubbe, Abraham and Thiele2012). It has been documented on R. blasii, R. ferrumequinum, R. hipposideros and R. mehelyi in regions such as Cyprus, Jordan and Syria (Amr and Qumsiyeh, Reference Amr and Qumsiyeh1993; Walter and Ebenau, Reference Walter and Ebenau1997; Ševčík et al., Reference Ševčík, Benda and Lucan2013).
On the island of Crete, Brachytarsina flavipennis Macquart, 1851 was reported on R. blasii for the first time by Scaravelli et al. (Reference Scaravelli, Priori and Georgiakakis2014). Additionally, N. africana has been recovered from R. blasii in Iran and Jordan (Amr and Qumsiyeh, Reference Amr and Qumsiyeh1993; Benda et al., Reference Benda, Spitzenberger, Hanák, Andreas, Reiter, Ševčík, Šmíd and Uhrin2014), from R. ferrumequinum and R. blasii in Cyprus (Ševčík et al., Reference Ševčík, Benda and Lucan2013) and from R. mehelyi in Libya and Syria (Hůrka, Reference Hůrka1982; Walter and Ebenau, Reference Walter and Ebenau1997). One female specimen was captured on R. ferrumequinum at El-Haouaria, Tunisia, in April 1955 (Vermeil, Reference Vermeil1960).
Although the chi-square test showed no significant association between host species and ectoparasite composition, some differences in relative prevalence were observed. Nycteribosca africana was more common on R. blasii, while Phthiridium biarticulatum occurred more frequently on R. ferrumequinum. These patterns may reflect subtle differences in roosting behaviour, microhabitat use or host physiology rather than strict host specificity. Similar non-exclusive associations have been reported in Mediterranean and North African bat–ectoparasite systems, where parasite prevalence varied among host species without clear statistical segregation (Bain, Reference Bain1966; Burazerović et al., Reference Burazerović, Ćakić, Mihaljica, Sukara, Ćirović and Tomanović2015; Frank et al., Reference Frank, Kuhn, Werblow, Liston, Kochmann and Klimpel2015; Khelfaoui et al., Reference Khelfaoui, Kebaci and Benyacoub2018). Such findings suggest that the distribution of bat ectoparasites is often influenced by ecological factors such as roost type, host colony size and microclimatic conditions, rather than strict host specialization (Amr and Qumsiyeh, Reference Amr and Qumsiyeh1993; Patterson et al., Reference Patterson, Dick and Dittmar2007; Bendjeddou et al., Reference Bendjeddou, Bouslama, Amr and Bani Hani2016, Reference Bendjeddou, Khelfaoui, Abiadh, Mechouk, Mihalca and Sándor2024; Webber and Willis, Reference Webber, Willis and Ortega2016). This highlights the ecological plasticity of bat flies and streblid parasites, which may exploit multiple sympatric hosts within the same habitat.
Endoparasites
The present study provides preliminary coprological data on helminths and arthropods found in bat guano in the El Hamdania sector of CNP. It should be noted that this coprological approach has important limitations: it yields only qualitative presence–absence data, does not permit estimation of parasite burden and cannot detect trematodes, which are among the most common bat endoparasites. A full helminthological necropsy involving dissection of all organ systems would be required to obtain comprehensive data on the endoparasite fauna. All species reported represent new records for the country.
Previous studies have demonstrated that flukes (digeneans) and roundworms (nematodes) are prevalent in horseshoe bat populations (Esteban et al., Reference Esteban, Oltra-Ferrero, Botella and Mas-Coma1991, Reference Esteban, Botella and Toledo1999; Ricci, Reference Ricci1995). Esteban et al. (Reference Esteban, Oltra-Ferrero, Botella and Mas-Coma1991) attributed this high prevalence to factors such as bat lifestyle, habitat preferences and diet. For instance, Strongylacantha spp. have simple life cycles, whereas Rhabditida spp. exhibit more complex parasitic cycles involving blood-sucking insect vectors (Bain, Reference Bain1966; Mészáros, Reference Mészáros1971). These insects consume larvae produced by mature female nematodes within bat hosts.
In related studies, Horvat et al. (Reference Horvat, Čabrilo, Paunović, Karapandža, Jovanović, Budinski and Čabrilo2015) analysed helminth infections in 17 individuals of the greater horseshoe bat (R. ferrumequinum) collected across 8 localities in Serbia. Helminth prevalence was high, with 88.2% of bats infected. Among the parasites identified, Strongylacantha glycirrhiza showed the highest prevalence (76.5%), comprising 65 of the 108 helminth individuals recovered. This study highlights the importance of both helminths and protozoa as common endoparasites in bats (Gardner and Jiménez-Ruiz, Reference Gardner and Jiménez-Ruiz2009; Klimpel and Mehlhorn, Reference Klimpel and Mehlhorn2014). For instance, Mühldorfer et al. (Reference Mühldorfer, Speck, Kurth, Lesnik, Freuling, Müller, Kramer-Schadt and Wibbelt2011) identified protozoan infections, such as Eimeriidae and Sarcocystidae, in Pipistrellus pipistrellus bats in Germany.
The observed helminth diversity in our study differs from those reported in the literature, potentially due to host species differences, dietary diversity or unique immune characteristics. For example, R. ferrumequinum exhibited consistently high parasitism levels, contrasting with the lower parasitism observed in R. blasii. This disparity mirrors findings by Mituch (Reference Mituch1964) in Czechoslovakia, where R. euryale displayed impoverished helminthofauna (14% parasitism) compared to R. ferrumequinum (43%). These differences may reflect variations in host immunity or ecological and behavioural factors, such as prey selection or activity patterns. It must be emphasized, however, that meaningful statistical comparison of endoparasite levels between R. ferrumequinum and R. blasii is not possible in the present study, given that only a single individual of R. ferrumequinum was sampled.
Cestodes in bats, particularly in Algeria, remain understudied. Comparative literature from France, Poland, Hungary, Germany and Austria reveals diverse Cestoda communities, with species of the Hymenolepididae family representing 4% of bat-associated taxa (Frank et al., Reference Frank, Kuhn, Werblow, Liston, Kochmann and Klimpel2015; Léger, Reference Léger2019). Differences in dietary composition, such as consumption of aquatic larvae – essential vectors for trematodes – may influence infection prevalence (Holmes, Reference Holmes1966). Such ecological nuances warrant further investigation to better understand host-parasite dynamics.
Additionally, mites are well-documented ectoparasites in bats, including ticks (Kolonin, Reference Kolonin2007; Hornok et al., Reference Hornok, Kontschán, Kovács, Angyal, Görföl, Polacsek, Kalmár and Mihalca2014) and other mite species (Tsai et al., Reference Tsai, Chang, Chuang and Chomel2011). At our study site, adult mites of the genus Alabidocarpus were identified in bat guano, underscoring the complex ecological interactions between bats and their parasites. Host-parasite relationships are deeply influenced by the behaviour and ecology of both parties (Patterson et al., Reference Patterson, Dick and Dittmar2007).
The helminth species reported here are consistent with those expected in greater horseshoe bats, and El Hamdania represents a newly confirmed locality for their geographical distribution. This study provides a foundational contribution to the parasitic fauna of bats in Algeria and lays the groundwork for future research. Such work could explore faunistic, epizootiological and biocenotic aspects, which offers valuable insights into regional and local biodiversity, as well as the intricate relationships within these ecosystems (Altringham, Reference Altringham2011; Fenton and Simmons, Reference Fenton and Simmons2014).
Conclusion
This study provides new data on the ectoparasite communities associated with cave-dwelling bats in CNP, together with supplementary coprological observations on endoparasite traces in guano. The findings should be interpreted in light of the methodological limitations inherent to the coprological approach, which yields only qualitative, fragmentary data and does not permit detection of trematodes or estimation of parasite burdens. Among the bat species captured in the Chiffa Tunnel (El Hamdania region), R. blasii (Blasius’ horseshoe bat) was the most abundant, followed by R. ferrumequinum (greater horseshoe bat) and N. leisleri (Leisler’s noctule), the latter newly reported in CNP.
Data availability statement
The datasets used and analysed during the current study are available from the corresponding author on reasonable request.
Acknowledgements
This work could not have been carried out without the help of Mr Ramdane Dehal (Director of the Chrea National Park ‘CNP’, Algeria), Mrs Salima Rahmouni (Veterinary Inspector at CNP), Mr Reda El-Ferroudji (Head of the El Hamdania Sector, CNP) and his team. We thank Prof Amel Milla (Higher National Veterinary School, Algiers) for her availability and the laboratory technician Mr Khaled Dalil for his help at the Zoology Laboratory of the Higher National Veterinary School.
Author contributions
Djamel Bendjoudi (conceptualization, methodology, investigation, data curation, resources, writing – original draft, writing – review & editing). Faiza Marniche (conceptualization, methodology, data curation, resources, writing – original draft). Sarah Banat (investigation). Hichem Zaoui (investigation). Haroun Chenchouni (visualization, writing – review & editing).
Financial support
This study was not funded by any source.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical standards
The study received approval from the Scientific Committee of the Faculty of Nature and Life Sciences at the University of Blida 1 in Algeria. All procedures were conducted in accordance with the standards and regulations established by the Ethics Committee of the SNV Faculty. Additionally, the study adhered to the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines, and the U.K. Animals Act 1986 guidelines.
Statement of animal rights
No animal suffered during the course of the studied. All the animal studies were conducted with the utmost regard for animal welfare, and all animal rights issues were appropriately observed.





 Open access
Open access