



Introduction
The cities of the Roman and Byzantine world around the Mediterranean and Black Sea are well known for their water supply infrastructure, including aqueducts, fountains, cisterns and wells. However, we have little knowledge of how clean their drinking water was, or if it was contaminated by human faeces and other waste. In this article we explore this topic, focusing on the wells of Phanagoria, a port on the coast of the Black Sea.
Water supply systems in ancient settlements provide important insights into historical sanitation practices and disease transmission patterns. The Northern Black Sea coast territory has been extensively studied in relation to water supply infrastructure in antiquity, with long-term archaeological excavations revealing various constructional elements including wells, cisterns, and ceramic plumbing (Kovalevskaya and Sedikova, Reference Kovalevskaya and Sedikova2005; Ushakov et al. Reference Ushakov, Makaev and Strukova2006). Modern researchers possess significant information about water supply, storage, and drainage systems from ancient times, with constructional features of technical structures used for water transportation and storage being well-documented, including methods of fresh water storage in tanks, rainwater collection, irrigation, and wastewater regulation (Deming, Reference Deming2019; Londra et al. Reference Londra, Antoniou, Theocharis, Dalezios and Angelakis2021; Angelakis et al. Reference Angelakis, Krasilnikoff and Tzanakakis2022, Reference Angelakis, Passchier, Valipour, Krasilnikoff, Tzanakakis, Ahmed and Dercas2023; Klingborg, Reference Klingborg2024).
Vitruvius, an ancient Roman architect and mechanic of the 1st century BC, provided detailed descriptions of water supply methods, fresh water source exploration, and engineering practices for the construction of various water supply and drainage facilities in the eighth book of his treatise De architectura (Ten Books on Architecture) (Morgan and Warren, Reference Morgan and Warren1914).
For instance, the first inhabitants of the city of Tauric Chersonese obtained their water from wells carved into limestone rock that contained aquifers. Similar structures were also discovered in rural farmsteads across the region. In addition to wells, cisterns were used to collect and store rainwater in courtyards and farmsteads (Belov, Reference Belov1948; Kovalevskaya and Sedikova, Reference Kovalevskaya and Sedikova2005). The monumental complex of the city’s water reservoir, built in the 3rd century AD, deserves special attention due to its volume of 1500 cubic meters. Such reservoir ensured the stability of the city’s fresh water supply during times of crisis.
No communal wells were discovered in the streets of the ancient city of Gorgippia (modern Anapa). However, nearly every courtyard had its own fresh water well, typically cut into the ledge rock (Alekseeva, Reference Alekseeva1997). Ceramic plumbing was also discovered during archaeological excavations at Gorgippia, mostly dating back to the Roman period (Alekseeva, Reference Alekseeva1997; Tsvetaeva, Reference Tsvetaeva1979, 66).
In the medieval layer of Hermonassa (Tamatarhe) dating to the 7th–10th centuries, researchers identified a well that was likely used as a reservoir (cistern) for water collection (Chkhaidze, Reference Chkhaidze2008). Similarly, during the research of the settlement Primorsky 23, located in the vicinity of Phanagoria, a well from the Classical period was discovered in the coastal part of the Taman Gulf on the flooded ancient site. The upper part of the well was made of well-worked blocks (Kolesnikov, Reference Kolesnikov2010).
Regarding Phanagoria, it is evident that a city would require a reliable water supply system. However, the archaeological excavations conducted thus far have not provided a comprehensive understanding of the water supply systems that were in place during the town’s operation from the 6th century BC to the 11th century AD. Though a small number of wells and elements of the water supply infrastructure have been found during the excavation history of the city. One of the first wells to be investigated was discovered at the Northern Excavation in 1939 (Blavatsky, Reference Blavatsky1950). The well was composed of well-cut stones resting on a base of wooden planks and had a square shape (0.85 × 0.88 m) in the cross-section. It was dug to a depth of only 1.3 m.
Another well, likely from the Late Antique period (3rd-5th century AD), was located along the road and was examined in the ‘Necropolis A’ excavation (Blavatsky, Reference Blavatsky1950, 40). The object had an almost square cross-section measuring 2.05 × 1.85 meters and was excavated to a depth of 5.75 meters. The upper part consisted of tightly fitted quadras, with rows of poorly worked stones and oblong-shaped slabs below.
In 1954, a well dating to the 4th century AD, composed of unworked stone, was discovered near the eastern defensive wall. The object was excavated to a depth of 2.5 metres and had a square shape in cross-section with a diameter of approximately 1.2 meters (Kobylina, Reference Kobylina1956).
In 1959–1960, an excavation was conducted at the Central site, which was situated in close proximity to the modern Lower Town site. This excavation revealed the remains of a well that was in use from the 1st to the 4th centuries AD (Kobylina, Reference Kobylina1963). The well was 6 m in depth and had an inner diameter of approximately 80 cm. The walls were constructed from large stones of varying sizes. The well belonged to a well-preserved house with a cellar.
Excavations of 1964–1965 on the south-eastern edge of the settlement revealed a circular well with a diameter of about 0.8 m in the layer dating to the 4th–5th centuries AD near the fortress wall. Twenty rows of masonry were preserved. The stones, of trapezoidal shape, were chiselled on the inner side along the curve, untreated on the outer side. The side walls of the well were chiselled, and the stones were well fitted to each other. It had a dedicated platform adjacent to the well for the convenience of lifting water.
The 2019 Upper Town excavations investigated a small round cistern dating to the second half of the 6th to early 5th century BC. It is probable that this cistern was used for collecting rainwater (Zavoikin and Kuznetsov, Reference Zavoikin and Kuznetsov2020).
Although archaeological research has provided relatively extensive information on water supply and drainage systems in Antiquity, there is a lack of knowledge regarding their parasitic infestation and certainly minimal data is available on the Northern Black Sea coast. Despite the fact that knowledge of constructive and functional features of water supply and water conservation sources, supplemented with information about their faecal contamination and parasitic contamination, is a unique source of information for bioarchaeological reconstructions, research on this topic is still insufficient (Slepchenko et al. Reference Slepchenko, Ostapenko and Khrustalev2023; López-Gijón et al. Reference López-Gijón, Hernández-Robles, Duras, Celma, Curto, González-Ballesteros, Dufour, Le Bailly and Eiroa2025).
Research conducted during 2019–2023 at the Lower Town excavation site has enabled a new perspective to be drawn on the problems of fresh water supply for the inhabitants of medieval Phanagoria during the Khazar period (7th–9th centuries AD). Results concerning parasite eggs in sediments from the drains of medieval Phanagoria during the Khazar period were published (Slepchenko et al. Reference Slepchenko, Ostapenko and Khrustalev2023). The analysis of parasite species composition found in the waterways of medieval Phanagoria, examined in a broad historical and archaeological context, led to the reconstruction of certain aspects related to nutrition, possible migratory directions and communication of the city’s population, and revealed a significant parasitic infestation of the studied materials (Slepchenko et al. Reference Slepchenko, Ostapenko and Khrustalev2023). However, no data were available at that time about the presence of parasite eggs in the wells, including one connected to drains. This information could be useful for our understanding of when the wells were used as water cisterns and when they were used as a source of water.
In addition, while interpreting the archaeoparasitological data, researchers encountered a problem in assigning eggs detected in the samples to human and/or animal parasites. To address this issue, they relied on the morphological and morphometric characteristics of the detected parasite eggs, as well as the archaeological context of the sampling. However, the question of whether the eggs derived from human and/or animal parasites could rarely be unambiguously resolved.
In recent decades, the analytical technique of gas chromatography–mass spectrometry (GC-MS) has been applied to solve such problems. For example, samples of room filling and cultural layer from the Middle Eastern Neolithic site of Çatalhöyük were analysed by GC-MS. The evaluation of sterols detected in the sediments enabled differentiation between samples of a faecal and non-faecal origin. In addition, some sediments were shown to contain human faecal matter (Shillito et al. Reference Shillito, Bull, Matthews, Almond, Williams and Evershed2011). The 21 coprolite samples from Paisley Cave in Oregon were analysed by GC-MS and paleogenetic methods. The use of faecal lipid biomarkers demonstrated that 3 coprolites dated to pre-Clovis are human, supporting the theory of pre-Clovis human occupation (Shillito et al. Reference Shillito, Whelton, Blong, Jenkins, Connolly and Bull2020). Archaeological excavations at Durrington Walls, a large Neolithic settlement in the UK dating from approximately 2500 BC and the site of the Stonehenge builders, revealed 19 coprolites. Of these, 5 contained eggs of intestinal parasites. GC-MS analyses determined that 4 of the coprolites most likely derived from dogs, while 1 was very likely of human origin (Mitchell et al. Reference Mitchell, Anastasiou, Whelton, Bull, Pearson and Shillito2022).
The objective of this study was to ascertain the sanitary condition of the water supply sources of medieval Phanagoria by determining the parasite range present in the soils from the wells. A further objective was to ascertain, through the use of GC-MS analysis of faecal sterols, whether the identified parasite eggs belonged to a specific animal species and/or to humans, and to evaluate the suitability of this approach for such materials. Additionally, the study aimed to determine the operational stages of the wells as sources of water supply in medieval Phanagoria.
Archaeological context
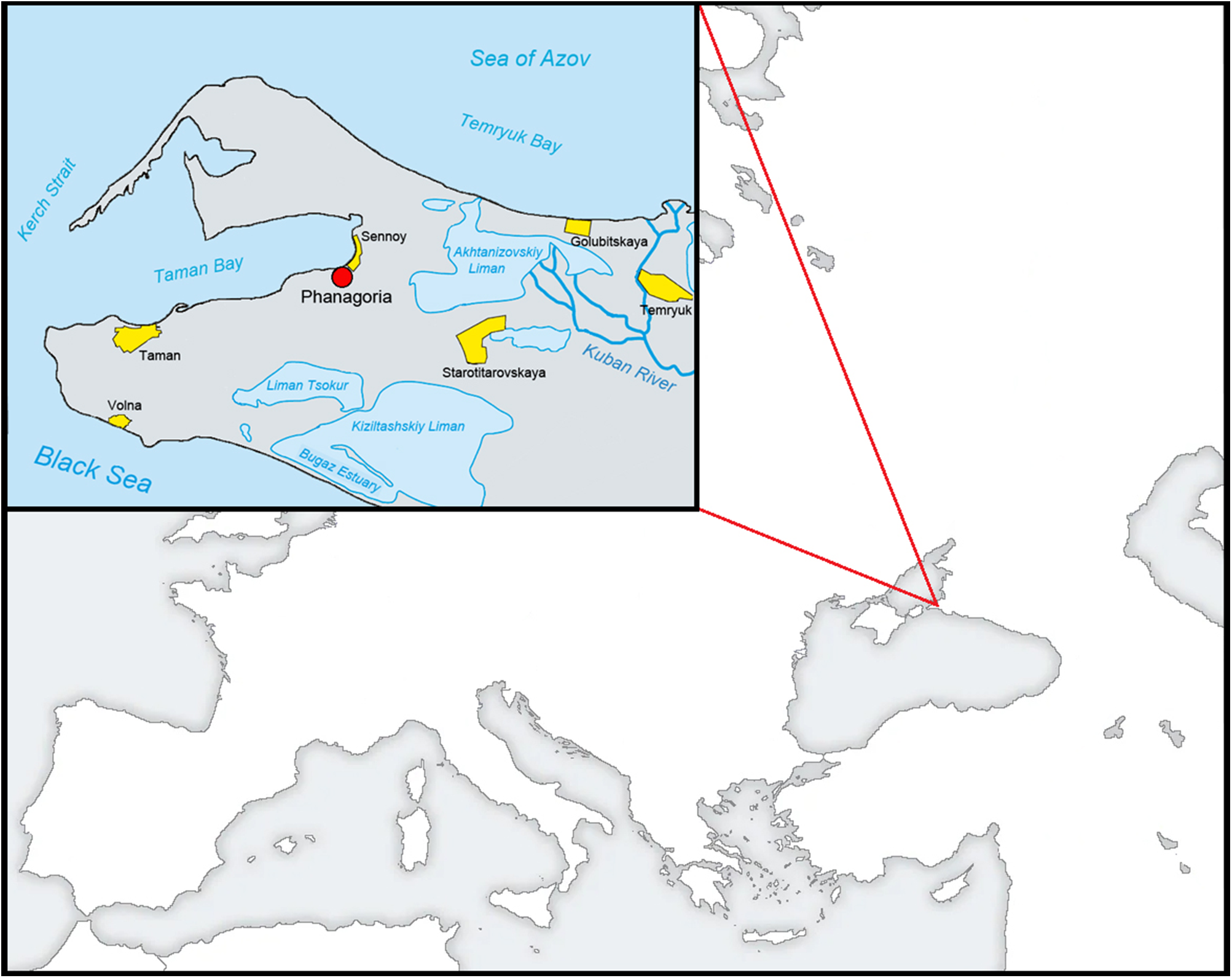
Phanagoria was the largest Greek city on the Taman Peninsula, founded around 540 BCE by colonists from the city of Teos in western Asia Minor (modern Turkey) (Figure 1). During its first decades, Phanagoria existed as an independent city-state (polis). At the end of the 5th century BCE, Phanagoria was forcibly incorporated into the Bosporan Kingdom, which united the Greek cities on the shores of the Cimmerian Bosporus (Strait of Kerch). Phanagoria served as the metropolis of the Asian part of the Bosporan Kingdom, as well as its important economic and cultural centre.
Location of the Phanagoria city, Taman Peninsula in the 8–9 centuries AD.

From the end of the 2nd century BCE to 63 BCE, Phanagoria, like the rest of Bosporus, was part of the Black Sea empire of King Mithridates VI Eupator (120–63 BCE). In 63 BCE, a revolt of Bosporan cities against Mithridates erupted in Phanagoria, leading to the king’s death. Under Roman rule in the 1st century BCE, Phanagoria was renamed Agrippia and retained its role as one of the most important cities of Bosporus.
The decline of the Bosporan Kingdom in subsequent centuries and the strengthening of its barbarization were also reflected in Phanagoria, which gradually lost the features of an ancient city. In the 4th century, Phanagoria was destroyed by the Huns but continued its existence. Subsequently, the city’s history was connected with Byzantium, Great Bulgaria, and the Khazar Khaganate. It was abandoned by its inhabitants approximately at the turn of the 9th–10th centuries.
In 2014, on the lower plateau of the Phanagoria settlement, at the intersection of the main ancient thoroughfare connecting the coastal part of the city with its southern outskirts and the main street leading from the eastern gates westward, an excavation site called Lower City was established, having the form of a rectangle stretched from west to east (50 × 40 m). The surface level of the excavated area slopes downward from south to north, as does the entire lower plateau.
During the recent excavations of the Lower City of Phanagoria, medieval layers of the 8th–9th centuries were uncovered (Figure 2), which allow for a more substantive examination of issues related to the city’s urbanism during this period (2010; Kuznetsov, Reference Kuznetsov2016; Kuznetsov and Ostapenko, Reference Kuznetsov and Ostapenko2019).
General view of the Lower city excavation site (medieval Phanagoria). (A) A well or a cistern made of unprocessed stones a stone drain (Object 33). (B) A well or a cistern made of unprocessed stones a stone drain (Object 46). (C) A stone drain (Object 38). (D) A stone drain (Object 39).

The investigated houses have relatively good preservation and consisted of 2, 3, 5, and more rooms. The floors consisted of a layer of beaten clay. External walls were constructed from unworked stones in 2 shells with filling using the opus spicatum technique, which appeared in Bosporus with the arrival of Khazar population in the second half of the 6th–7th centuries and was characteristic of all Khazar monuments from the 7th–10th centuries. Internal partition walls were built from mud brick on stone foundations laid in a single row of small stones.
The rooms had multifunctional purposes. Practically all houses uncovered to date have courtyards. Along the long wall of the house flanking the courtyard, a canopy was made, supported by wooden posts mounted in stone bases. In all probability, public buildings and some houses of wealthy residents could be roofed with tiles. Large pithoi (vessels for storing grain or wine) were located inside rooms and courtyards.
The medieval quarters of Phanagoria were oriented practically according to cardinal directions (with a deviation of about 10°). When paving streets, worked stone was used in rare cases; most often streets were covered with broken ceramics mixed with marine mollusks (mussels and oysters), animal bones, and small rubble (apparently waste from stone-cutting work).
The Phanagoria settlement remains extremely understudied to this day. Archaeological excavations have uncovered only small sections of the settlement in total (about 2%), which is explained by the large size of the city itself (more than 65 hectares) and the thickness of the cultural layer (about 7 m). When studying medieval Phanagoria, the question of water supply organization always remained open, since previous research had not discovered water pipes, wells, and cisterns from the Khazar period.
Regarding the ancient period, wells were found during excavations on the city’s territory, which the Phanagoriotes stopped using in the late ancient period. This was apparently connected with changes in groundwater levels or environmental pollution from anthropogenic impact (for example, the arrangement of garbage and household pits in the courtyards of urban quarters with sufficiently dense construction).
The description of well 33, along with information on the water conduits discovered during the excavations of the lower town (Objects 38 and 39), was previously published by Slepchenko et al. (Reference Slepchenko, Ostapenko and Khrustalev2023). Further details on the layout and findings can be found in that publication. The relative positions of the objects studied are shown in Figure 2.
Description of Object 46
Object 46 was excavated in 2023 and was discovered during excavations in square A4, 2 m from the western side of the Lower Town excavation and 17 m from the southern side (Figure 3). It was a well (cistern) constructed from reused stones and covered with a lid, which was a reused building block (there are manufacturing holes for fixing the block with other architectural elements with the help of dowel). The well was almost filled with soil, and the wellhead section was free of filling to a depth of 85 cm. The upper stones of the crown were recorded at a depth of 4.34–4.4 m from the modern-day surface. The crown is composed of relatively large stones. The masonry is executed with great care and precision. In plan, the object has a rectangular shape that widens with depth. The inner section of the mouth measures 0.54 × 0.55 m, while the lower section (at the time of conservation) measures 1.15 × 1.19 m. The neck is lined with irregularly shaped recycled blocks. The sides of the well are constructed of medium and large stones that are well fitted to each other, without the use of a binding solution. The well mouth exhibits an important characteristic feature: the southern facade is laid in 2 rows in the opus spicatum (or herringbone) technique and supported by a massive shoulder stone, which indicates that it was repaired (at least 2 construction periods are evident) (Figure 3). The lower portion of the southern facade, as well as all facades, is constructed using a technique that is similar to opus incertum. The well was excavated to a depth of 3.81 m.
Object 46: Medieval well-cistern from Phanagoria (8th–9th centuries AD). (A) Stone covering slab of the well-cistern (Object 46) made from a reused building block that served as protective covering of the mouth. (B) Mouth of the medieval well-cistern of rectangular shape with internal dimensions of 0.54 × 0.55 m, constructed from irregularly shaped reused stone blocks. (C) Longitudinal section of the well-cistern, demonstrating the construction that widens downward, built from medium and large stones without binding mortar.

Dating of the object: the well is located within the boundaries of square A4, where the common yard of 2 medieval houses 10 and 16 of the same time of construction and functioning (the 8th–9th centuries) was investigated earlier (Figure 2). This is evidenced by the level of wall bases and their unified orientation. Between the 2 houses was an alley 1.35–1.4 m wide, which led to their common courtyard with galleries. Stones were set at an equal distance from the walls of the houses as a support for wooden columns under the sheds. Based on planigraphic analysis of houses 10 and 16, it can be concluded that the well ceased to exist at the time of construction of these houses.
However, the second construction period of the well (repair works of the southern facade in the opus spicatum technique) allows us to conclude with confidence that it existed during the 7th and a short period of the 8th centuries. The opus spicatum masonry is also an important date marker, reflecting a new construction technique following the arrival of a new population (Khazars) to the city in the late 7th and early 8th centuries.
Materials and methods
As the result of partial excavation of the contents of Objects 33 and 46 during the previous field seasons (2021–2022 years), only the bottom parts of these structures were available for investigation. During previous years’ excavations, removal of the filling was halted at a depth of 4.2 m for Object 33 and 2.90 m for Object 46 due to the reduction or complete absence of artifacts in the fill and the associated danger of proceeding. Given that the filling of Object 33 was excavated to a greater depth, only 3 layers (45 cm) were available for investigation before the emergence of groundwater.
Sampling was conducted in layers from the bottom of the wells. Two samples of approximately 300 cm2 in volume were randomly taken from different sections of each excavated horizon, which was about 15 cm in thickness. Six samples were taken from 3 layers at Object 33 before reaching the aquifer at a depth of 4.64 m. At Object 46, 12 samples were collected from 6 layers excavated to a depth of 3.81 m. Sampling was discontinued due to possible collapse of the object’s upper sections. Between the fifth and sixth (lowest) layers, a clay layer up to 5 cm in thickness was identified at a depth of 3.65–3.60 m, from which one sample was taken.
Two control soil samples were collected directly during the excavation. One sample was obtained from the modern surface located 50 m away from the excavation, while the second was collected in close proximity to the Lower Town excavation at a depth of 5 cm.
The samples were packed in zip-lock bags, labelled and transported to the Tyumen Scientific Center laboratory.
For analysis, 6 samples were taken from 3 layers of Object 33, and 12 samples were taken from 6 layers and 1 intermediate layer between the fifth and sixth layers of Object 46. In addition, samples were collected from 2 drains (Objects 38 and 39), as part of the investigation of drains (Objects 38 and 39) described and published earlier (Slepchenko et al. Reference Slepchenko, Ostapenko and Khrustalev2023).
Methods of archaeoparasitological analysis
From each sample, between 85 and 110 g of soil was collected for archaeoparasitological study. The collected sediment was then processed according to a standard scheme with the use of 0.5% trisodium phosphate (Na3PO4) solution (Callen and Cameron, Reference Callen and Cameron1960; Slepchenko et al. Reference Slepchenko, Slavinsky, Ivanov, Rakultseva, Siben, Tsybankov and Galukhin2020). After isolation of the organic component, 20 microslides were prepared from each sample. Microscopy was performed using AxioSkop 40, Zeiss Primo Star, MicMed 2 var.2. microscopes at 100 × and 400 × magnification. AxioVision 4.6, Axiocam 105 colok and Scope Photo 3.0 applications were used for measurement.
We used the absolute size range (minimum, min; maximum, max), arithmetic mean (M) with standard deviation (SD) calculation as descriptive statistics for the measured egg characteristics. MS Excel software from MS Office 2019 was used to calculate numerical parameters. Statistical analysis of helminth egg concentrations was performed using a test for comparing 2 independent Poisson rates. All analyses were performed using R statistical software (version 4.3.1; R Core Team, 2023).
To identify the species of human parasite eggs, we consulted the manuals of Ash and Orihel (Ash and Orihel, Reference Ash and Orihel2007) and Gaevskaya (Reference Gaevskaya2016), as well as materials from the CDC DPDx website (Laboratory Identification of Parasites of Public Health Concern).
Methods of faecal biomarker extraction
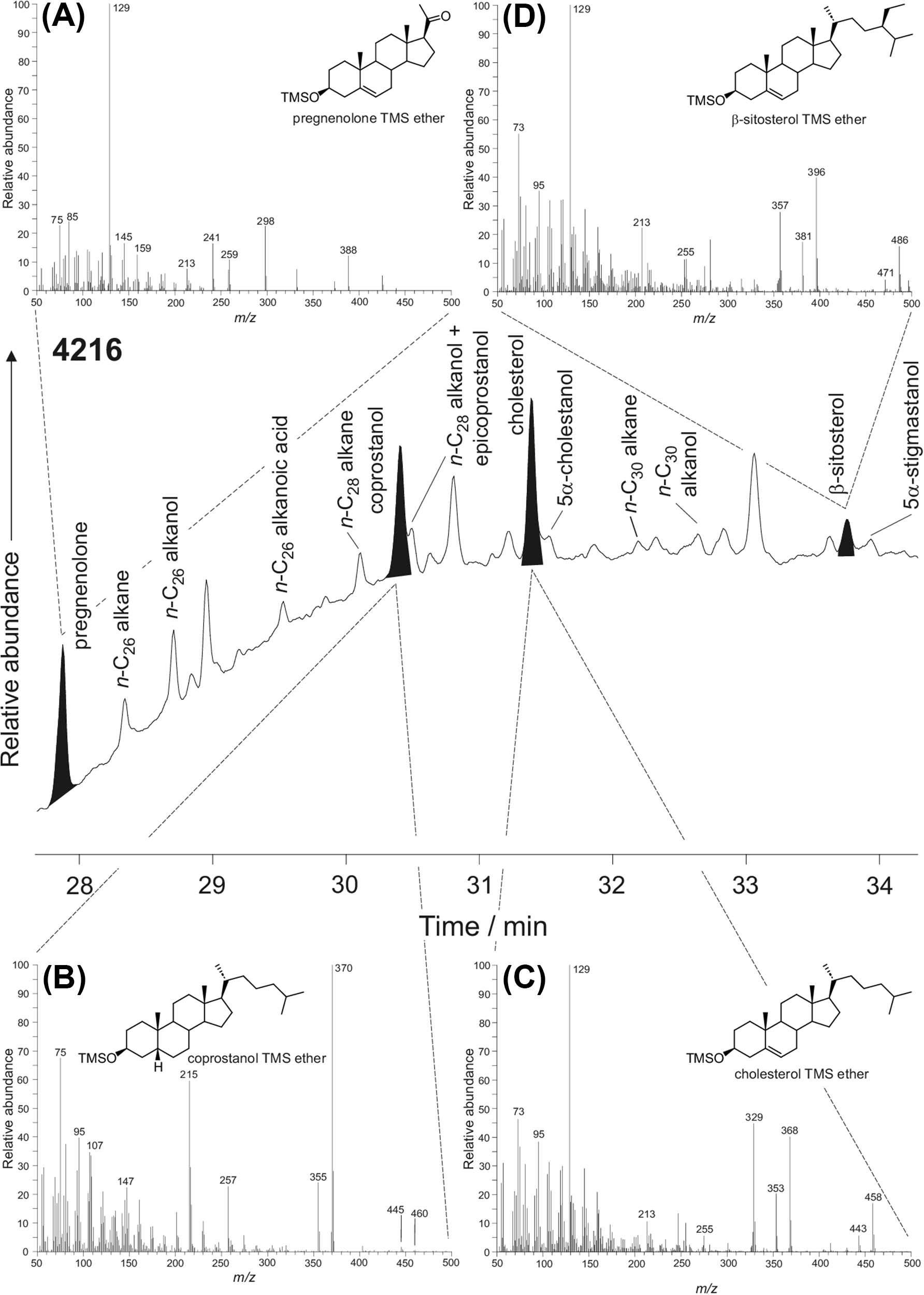
Faecal biomarker analysis was conducted using protocols established by Wu et al. (Reference Wu, Hu, Yue, Yang and Zhang2010). Briefly, sterols were extracted from soil samples (1–1.5 g) using methylene chloride/methanol (2:1 v/v) with pregnenolone as an internal standard. Derivatization was performed with BSTFA + 1% TMCS at 60°C for 1 hour, followed by GC-MS analysis on an Agilent 5977 instrument equipped with an HP-5 MS column. Separation and detection parameters replicated those of Wu et al. (Reference Wu, Hu, Yue, Yang and Zhang2010), with sterol identification based on selective ion monitoring (SIM) of trimethylsilyl derivatives.
Table 5 provides the layer numbers, laboratory sample numbers, and sample weights.
Results of archaeoparasitological research
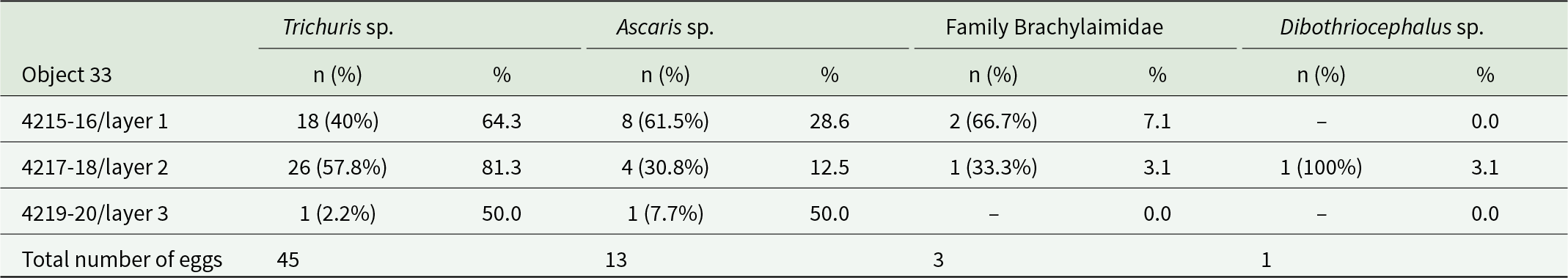
Eggs of the following parasites were found in the soil samples from the studied layers of the Lower Excavation wells (cisterns). One egg was found in layer 2 of Object 33 (3.1% of the total number of eggs found in this layer, with elongated asymmetrical shape, light brown colour, single-layer thick shell with a small knob at the opposite end of the egg. The cap was absent. Based on morphological and morphometric characteristics, the egg was assigned to the genus Dibothriocephalus (Figure 4A) (Table 1).
Eggs suggestive of Dibothriocephaliidae family (A), Trichuris trichiura/suis (B), Trichuris muris (C), Calodium hepaticum (D), Ascaris lumbricoides/suis (E) and family Brachylaimidae (F).

Total number of eggs in the sample/proportion of eggs from the total number of parasite eggs in the layers in Object 33

n (%) – number of parasite eggs found in the layer and percentage of the total number of eggs detected.
% – proportion of parasite eggs from the total number of detected eggs of all parasites found in the layer.
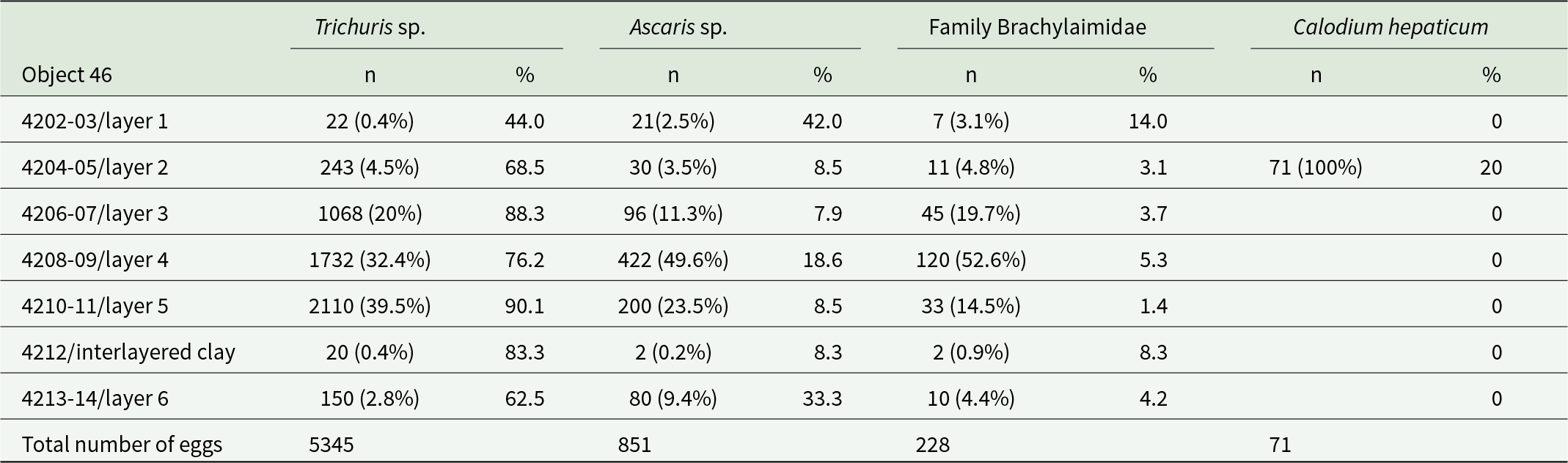
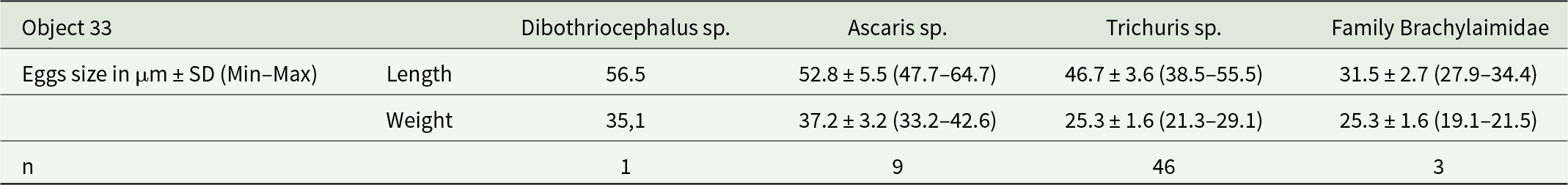
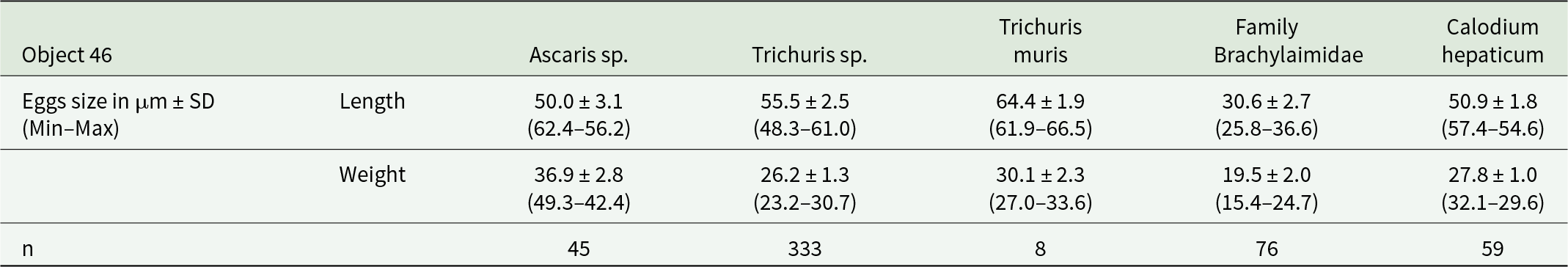
Nematode eggs of the genus Trichuris were found in all layers of Objects 33 and 46 without exception. Depending on location, the proportion of these eggs ranged from 44% to 88.3% of the total number of helminth eggs found (Table 1, 2). The nematode eggs of this type exhibited a distinctive elongated barrel-shape, narrowing towards the poles with mouths and a thick shell with a double contour. All eggs lacked plugs at the poles, and a large number of eggs exhibited degraded outer shells. A correction factor of 1.1 was applied when estimating length measurements of nematode eggs of the genus Trichuris, to account for the absence of polar plugs and for eggs with degraded outer shells.
Total number of eggs in the sample/proportion of eggs from the total number of parasite eggs in the layers in Object 46

n (%) – number of parasite eggs found in the layer and percentage of the total number of eggs detected.
% – proportion of parasite eggs from the total number of detected eggs of all parasites found in the layer.
In the examined layers from Object 33, all eggs matched the T. trichiura/suis species group in size: 49–65 × 20–29 (cf. 54 × 22) µm (by CDC) (Figure 4B) (Table 3).
Parasite species and egg size in the layers in Object 33

Most eggs found in layers from Object 46. Match the T. trichiura/suis species group in size: 49–65 × 20–29 (mean 54 × 22) µm (by CDC) (Figure 4B). However, some eggs fall outside of this size range and are more consistent with the T. muris species: 63.7 ± 2.2 (51.9–70.0) × 31.3 ± 1.3 (27.7–38.2) (as per Koyama, Reference Koyama2013) (Figure 4C) (Table 4).
Parasite species and egg size in the layers in Object 46

Additionally, brown-coloured, round or oval-shaped eggs with thick, knobby shells and amorphous contents were observed in all layers of Objects 33 and 46 (Figure 4D). Based on their morphological and morphometric characteristics, these helminth eggs can be assigned to the genus Ascaris (A. lumbricoides/suis) (Tables 3,4). The highest proportion of this helminth’s eggs (50% of the total number of eggs in the sample) was found in the third, lowest layer of Object 33, and the lowest proportion (7.9%) was found in the third layer of Object 46 (Tables 1,2).
Virtually all layers, except layer 3 of Object 33, contained eggs with distinctive features of ellipsoidal asymmetric shape and thick, dark brown shells. However, the tubercle on the opposite pole from the cap, typical for numerous trematodes, was absent, but some eggs had thickening of the wall at this place. Egg showed little variation (see Tables 3,4). The length of trematode eggs was measured with correction for missing caps (coefficient 1.1). Based on morphological and morphometric characters, we identified the eggs as trematodes of the family Brachylaimidae (Figure 4F).
One of the samples from layer 2 of Object 46 contained eggs with a distinctive outer shell, in most cases partially or completely degraded, and about a half of eggs exhibited degraded polar structures of the mouths.
To obtain comparable data, we measured the length of eggs between the bases of the mouths, the width was measured excluding the outer shell. For the final correction, we added doubled values of the average shell thickness to the measured width and the length of the mouths to the measured length. According to preliminary measurements, these values were 2.2 and 3.7 μm, respectively. Based on the distinctive morphology and size, the eggs of this type belong to nematodes of the species Calodium hepaticum (Figure 4E) (Table 4).
The most comprehensive reconstruction and tracing of some key aspects of the wells functioning was possible on the example of Object 46. During the layer-by-layer excavation and sampling of the filling, the presence of a 5 cm thick clay layer, which divided the filling into 2 horizons, was revealed at a depth of 3.65 m. Archaeoparasitological examination of soil samples revealed the presence of geohelminths above and below this layer. However, the concentration of all parasite eggs was significantly higher in the layers above the clay layer (1171.5 eggs per sample) than in the layers below (120 eggs per sample), representing a 9.8-fold difference. Statistical analysis using a test for comparing Poisson rates confirmed this difference as highly significant (z = 41.379, P < 0.001). It is noteworthy that the minimal number of helminth eggs was detected in the layers below the clay layer of Object 46 and in soil samples collected from horizons immediately adjacent to the aquifer from Object 33. Moreover, the concentration of helminth eggs was minimal in the clay parting itself, which may be attributed to the diffusion of eggs into the parting from higher layers or to contamination with soil from higher/lower layers during sampling. This statistically robust difference (based on 2343 total eggs above vs 240 eggs below the clay layer in 2 samples each) demonstrates a dramatic shift in parasitological conditions and provides robust evidence for a functional transition in the well’s usage pattern.
A single egg with morphometric characteristics similar to trematodes of the family Brachylaimidae was found in one of the control samples collected from the modern surface 50 m away from the excavation. In the second control sample, which was collected directly from the Lower Town excavation at a depth of 5 cm, no parasite eggs were identified.
Gas chromatography–mass spectrometry results
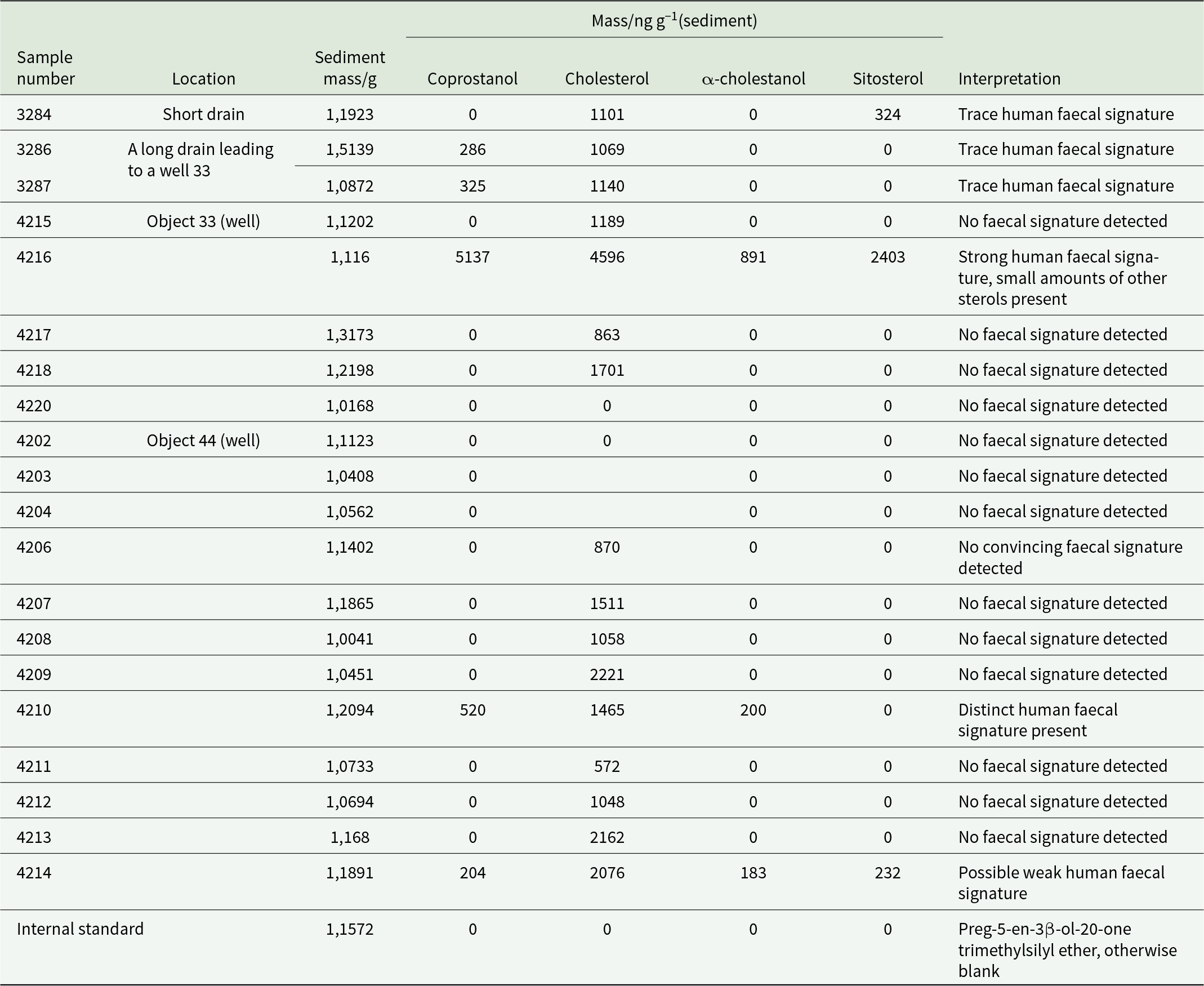
Table 5 summarizes the sterols detected in each of the archaeological samples along with their mass fraction in each sample. As expected, the internal standard, preg-5-en-3β-ol-20-one, was detected in all analyses as its TMS ether. In addition, cholesterol was detected in nearly all samples. It was absent only in 2 samples from Object 33 (first layer, laboratory sample #4215 and the third layer, laboratory sample #4220) and Object 46 (code: 4203 and 4204). Coprostanol and β-sitosterol were detected in the first layer sample of Object 33 (laboratory sample #4216; Figure 5) and to a lesser extent in the short drain (laboratory sample #3284) and another sample from Object 33 (laboratory sample #3284). Coprostanol was also detected in sample from Object 46 (laboratory sample #4210).
A partial gas chromatogram obtained for the soil from layer 6 of Object 33/laboratory number 4216 and mass spectra for (A) pregnenolone (internal standard), (B) coprostanol, (C) cholesterol and (D) β-sitosterol.

A summary of the sterol compounds detected in each archaeological sample

Discussion of archaeoparasitological data
The previously conducted research of the water conduits of the Lower City (Slepchenko et al. Reference Slepchenko, Ostapenko and Khrustalev2023) allowed to assume that the studied objects in different periods of their existence could have different functional purposes. The study of soil samples in the present work allowed to confirm this opinion.
It is hypothesized that the high concentration of parasite eggs in samples from the studied objects 33 and 46 may have resulted from their prolonged arrival with water contaminated with helminth eggs through water conduits, probably collecting rainwater and sewage from the areas adjacent to the wells, or by other means. It is possible that parasite eggs were transported to the wells via surface conduits that collected water from roofs of surrounding buildings. Over time, the eggs may have accumulated and concentrated at the bottom of the wells due to sedimentation.
Nevertheless, this mechanism can only be established with a certain degree of certainty for well 33, as it is connected a stone drain №38 No water conduits leading to Object 46 were found, and the mechanism of helminth eggs entry is not quite clear. Additionally, it is challenging to reconstruct the stages of functioning of this object. It is known from the archaeological context that the necks of both objects were blocked by large stones. Upon the removal of these stones, it became apparent that the sediment deposited directly beneath in the case of Object 33, or that there was a cavity measuring approximately 1 m in depth in the case of Object 46. It is hypothesized that the observed difference in helminth egg concentrations above and below the clay layer may indicate different periods of Object 46 functioning. At the earliest stage, below the 3.81 m level, the object is believed to have existed as a well with minimal contamination of the water with helminth eggs and with little or no external inflow. The low concentration of helminth eggs in the layers of Object 33 adjacent to the aquifer, in comparison to samples from Object 46, likely indicates that this object also originally functioned as a well. At this stage of the wells’ functioning, they were probably not connected to conduits.
In the subsequent operational phase, the base of the well was filled with clay for various reasons, including a decline in groundwater levels, siltation, and so forth. This was done to create a waterproof layer to prevent rainwater and/or wastewater collected by drains from seeping deeper into the ground. Subsequently, Object 46 was utilized as a cistern, with water being conveyed to it via water conduits. During this period of operation as a cistern, the concentration of helminth eggs in the studied layers increased significantly.
At the final stage, both objects were utilized as garbage pits, as indicated by the presence of a significant number of amphorae and building stone fragments in the filling of the objects. The concentration of these materials decreased with depth. Furthermore, the presence of parasite eggs of the species Calodium hepaticum in the samples lends support to this hypothesis, as they could only have ended up in the object with the tissues of dead animals, such as rats.
Discussion of soil sample examination for faecal biomarkers
It was postulated that if the samples selected for study from objects 33 and 46 were derived from household garbage, manure, human and/or animal faeces, which may have entered the wells (cisterns), it would be possible to detect faecal biomarkers that are associated with particles of soil filling. In addition, soil samples from the conduits that brought water to the wells were analysed, which were expected to exhibit faecal biomarkers, but at lower concentrations. It was not anticipated that analysis of control samples would yield the detection of faecal biomarkers. At the same time, since manure can be detected in any soil, we planned to detect faecal biomarkers by comparing study and control samples.
The presence of a limited occurrence of human faecal signatures in a minimal number of samples, in conjunction with the presence of helminth eggs in virtually all samples studied, is perplexing. Most soils yielded a weak total lipid signature so in all likelihood extraction of a greater mass of soil in each case would have resulted in a greater number of positive hits. Nevertheless, we posit that even the minimal number of faecal biomarkers detected indicates contamination of wells with faeces. The identical sterols found in samples from wells (cisterns) and drains may indicate their interconnection in terms of sterols entering wells via drains. This is supported by the archaeoparasitological data.
It is of interest that all faecal biomarkers in the samples from Object 46 were found above the clay layer. It is probable that during the period when Object 46 was used as a well, contamination of the water was minimal or non-existent. Only at the cistern stage of existence of Object 46 did contamination increase, which aligns with the archaeoparasitological data, showing that the highest number of parasite eggs was found in the samples above the clay layer.
Regarding the source of contamination, it can be tentatively concluded that the preponderance of coprostanol over all other stanols in terms of frequency of occurrence and mass fraction, when considered alongside the archaeoparasitological data, suggests that the contamination is most likely to have originated from humans and/or, where coprostanol is absent or a minor component, dogs. Coprostanol is the primary 5β-stanol in human faeces and accounts for approximately 60% of the total sterol content (Leeming et al. Reference Leeming, Ball, Ashbolt, Jones and Nichols1984). With regard to the presence of faecal biomarkers in the control samples, as predicted, they were not detected. The results of soil samples from the wells (cisterns) and drains revealed a minimal presence of faecal biomarkers, while significant amounts of intestinal parasite eggs were present in the same samples. It is possible that helminth eggs, being relatively heavy objects, entered the wells (cisterns) with contaminated water from the drains and were deposited in the bottom layers. Due to their higher resistance to environmental impact, they were less susceptible to degradation. Meanwhile, sterol molecules were probably constantly eliminated from wells (cisterns) as a result of degradation by aerobic bacteria or for some other unestablished reason(s).
The soils are very organic lean (the excessively high column bleed in the chromatograms indicates that the extracts are at a low concentration) which is why a faecal signature may not have been observable. Extracting more soil and/or using a more rigorous extraction (Soxhlet, microwave assisted) may have helped improve this.
Refined data on the sanitary conditions and dietary features of the population of the medieval city of Phanagoria
The acquisition, albeit minimal, of human signatures of faecal biomarkers and the absence of animal signatures allows us to consider humans specifically as the primary source of faecal contamination in the city, namely the investigated objects and drains. Furthermore, new archaeological data indicate that the entire investigated complex of structures, to which the drains and wells belonged, was under the management of a synagogue that existed at this location for at least 500 years (Kuznetsov, Reference Kuznetsov2024). The latter allows us to confidently rule out pigs, discussed in the previous article, as one of the possible sources of contamination. Thus, based on the refined data, the discovered helminth eggs, namely of the genera Ascaris, Trichuris, and Dibothriocephalus, can with greater confidence be attributed to the human species Ascaris lumbricoides, Trichuris trichiura, and Diphyllobothrium latum.
Following the clarification of the species affiliation of the found parasite eggs and the connection between the drains and wells, archaeoparasitological data allow us to confirm a number of important conclusions about the sanitary condition of the medieval settlement and the dietary features of its inhabitants, previously obtained from the study of soil samples from drains discovered in the same part of the city (Slepchenko et al. Reference Slepchenko, Ostapenko and Khrustalev2023). Thus, the analysis of samples from the wells showed a high concentration of geohelminth eggs (Ascaris lumbricoides, Trichuris trichiura), which is direct evidence of unsatisfactory sanitary conditions in the settlement and the general hygiene of its inhabitants during the medieval period. The detection of parasite eggs of the species Calodium hepaticum, the source of which could only be dead animals, such as rats, also points to unfavourable sanitary and hygienic conditions; the latter, being synanthropic rodents, could also be a source and vector of some dangerous infections.
The high level of geohelminth contamination in soil samples from the upper layers of the wells (layers 4 and 5 of Object 46, located above the clay layer at 3.65–3.60 m depth), contrasted with lower contamination levels in samples from deeper layers (layer 6 of Object 46, located below the clay layer), as well as from the drains (a long drain leading to Object 33), indicates a likely high risk of gastrointestinal disease spread among the population of medieval Phanagoria, given the identical transmission routes of geohelminths and various intestinal infections. Moreover, this situation suggests that water collected in these structures was unlikely used for human drinking purposes without prior purification and was more likely used for household needs: watering plants and watering livestock, which, in turn, contributed to maintaining the cycle of geohelminth reinvasion.
The significantly lower contamination level of soil samples from layers below the clay layer indicates that the well could only have been used as a source of quality water during the initial period of its operation. This also testifies to minimal geohelminth contamination of the well during this period.
The analysis of soil from the wells once again confirmed the presence of freshwater fish in the diet of the medieval inhabitants of Phanagoria and the importance of fishing in freshwater bodies (the Kuban River, the limans of the Taman Peninsula). Furthermore, the clarification of the probable source of the parasite eggs (human) allows us to attribute the Taenia sp. eggs found in the drains to the species Taenia saginata, which confidently points to the consumption of infested beef and possible disease prevalence in the herd concerning some anthropozoonotic infestations.
Conclusions
The Northern Black Sea coast has been extensively studied from an archaeological perspective. However, there has been a paucity of attention paid to the issues of water supply to the ancient population. There is even less information about the contamination regarding water supply sources with parasitic and infectious agents.
According to the results of archaeoparasitological research, it was possible to determine 3 stages of functioning of wells at the Lower Excavation of medieval Phanagoria. At the first stage of its existence, the well was used as a source of water. At the next stage, probably due to the lowering of the groundwater level, the well was covered with clay, which created a waterproof layer, and the well was used as a cistern in which the water supplied to it by the water pipes was accumulated. In the last stage, the cistern wells were used as rubbish pits.
The study once again demonstrated the promise of combining archaeoparasitological and GC-MS data to determine the source of intestinal parasite eggs. The GC-MS examination of soil samples provided clear signals of the human origin of faecal contamination in soil samples from drains and wells.
The somewhat confounding results of the study, indicate that it is necessary to continue further work to establish the mechanisms leading to the co-occurrence (or not) of parasite eggs and faecal biomarkers in sewage effluent and thereby better support the use of GC-MS in archaeoparasitology for the study of sediments from wells, drains and cultural layers in general.
Acknowledgements
We are very grateful to the editors and reviewers for taking the time and energy to help us improve the article.
Author contributions
Conceptualization, design of study, and correspondence: Sergey Slepchenko; archaeological data collection and investigation: Sergey Slepchenko, Sergey Ostapenko; archaeoparasitological investigation: Sergey Slepchenko, Alexander Khrustalev; the section of the article is dedicated to gas chromatography and mass spectrometry: Anastasiya Kashevskaya, Nikolaj Tretyakov and Ian Bull; literature review and synthesis of findings: Sergey Slepchenko, Sergey Ostapenko, Ian Bull; manuscript drafting: Sergey Slepchenko, Sergey Ostapenko, Anastasiya Kashevskaya, Nikolaj Tretyakov, Alexander Khrustalev, Ian Bull; critical review and editing: Sergey Slepchenko.
Financial support
This work also was supported by the Tyumen Scientific Centre SB RAS within the framework of the state assignment of the Ministry of Science and Higher Education of the Russian Federation (Project No. FWRZ-2021-0006)
Ethical approval
Not applicable.




 Open access
Open access