


Introduction
Freshwater ecosystems are home to a remarkable degree of biodiversity (approximately 10% of all so-far-known species, according to Strayer and Dudgeon, Reference Strayer and Dudgeon2010) and are undoubtedly one of the most threatened types of ecosystems in the world (Sala et al., Reference Sala, Chapin, Armesto, Berlow, Bloomfield, Dirzo, Huber-Sanwald, Huenneke, Jackson, Kinzig, Leemans, Lodge, Mooney, Oesterheld, Poff, Sykes, Walker, Walker and Wall2000). In Iraq, freshwater ecosystems provide a variety of services; however, traditional fisheries are quickly being replaced by the farming, cultivation and harvesting of non-native and invasive species. These are, namely, Carassius auratus (Linnaeus, 1758), Ctenopharynodon idella (Valenciennes, 1844), Cyprinus carpio Linnaeus, 1758 and Hypophthalmichthys molitrix (Valenciennes, 1844), which pose the greatest threat to the local endemic freshwater fauna and are replacing formerly endemic cyprinids (e.g. Khalaf, Reference Khalaf1961; Al-Hassan et al., Reference Al-Hassan, Hussain and Saoud1989; Jawad, Reference Jawad2006). Currently 6 cyprinoid species (sensu Tan and Armbruster, Reference Tan and Armbruster2018) are recognized as invasive in Iraq (Al-Faisal, Reference Al-Faisal2020), posing a potential threat to the native fauna. The native freshwater fish fauna is relatively well known, according to the most recent checklist by Al-Faisal (Reference Al-Faisal2020) and includes 31 species (cyprinoids being the most speciose suborder in the area), out of which more than half are considered as threatened (17 species according to the IUCN Red List, 2023). The relatively high species diversity is mainly due to the geographical position of Iraq, as it overlaps 3 major biodiversity hotspots (Myers et al., Reference Myers, Mittermeier, Mittermeier, de Fonseca and Kent2000). The most species-rich genera of cyprinoids are Luciobarbus, with 6 species, and Capoeta and Garra, with 4 each. The freshwater diversity in Iraq is mainly bound to the Euphrates and Tigris basins, as these represent the major river systems in the area. These river systems were important historical dispersion crossroads for cyprinoids and besides the entirely endemic genera (e.g. Capoeta), the rivers currently harbour species closely related to the congeners common in Africa (e.g. Garra, Luciobarbus), eastern Asia (e.g. Cyprinion) and Europe (e.g. Barbus, Luciobarbus) (Kottelat and Freyhof, Reference Kottelat and Freyhof2007; Coad, Reference Coad2010; Yang et al., Reference Yang, Sado, Hirt, Pasco-Viel, Arunachalam, Li, Wang, Freyhof, Saitoh, Simons, Miya, He and Mayden2015; Froese and Pauly, Reference Froese and Pauly2023).
Although the diversity and distribution of cyprinoids have been thoroughly investigated in Iraq (Coad, Reference Coad2010; Al-Faisal, Reference Al-Faisal2020; Abdullah et al., Reference Abdullah, Abdulah and Onda2022), little is known about their parasites. Such parasites represent a biological threat to already endangered native fish, especially considering the co-invasion of parasites with non-native fish species (Lymbery et al., Reference Lymbery, Morine, Kanani, Beatty and Morgan2014; Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018; Šimková et al., Reference Šimková, Řehulkova, Jean Robertin, Jorissen, Scholz, Faltynková, Mašová and Vanhove2018; Wilson et al., Reference Wilson, Saunders and Hutson2019). The highest metazoan parasite diversity in Iraq is reported for monogeneans (e.g. Mhaisen and Al-Rubaie, Reference Mhaisen and Al-Rubaie2016; Mhaisen and Abdullah, Reference Mhaisen and Abdullah2017; Mhaisen et al., Reference Mhaisen, Al-Mayali and Al-Abodi2019). These ectoparasitic flatworms mainly infest ectothermic vertebrates and several invertebrate taxa, as their life cycle is strictly limited to the aquatic environment. The taxonomy of monogeneans is quite complex and mainly based on the composition and morphology of the hard structures of the attachment organ (i.e. haptor) and the reproductive systems, especially the male copulatory organ (MCO) (Boeger and Kritsky, Reference Boeger and Kritsky1993; Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009; Řehulková et al., Reference Řehulková, Seifertová, Přikrylová, Francová, Scholz, Vanhove, Smit, Jayasundera and Gelnar2018). As the localization of a particular species on a host may differ, the haptor is considered as a highly morphologically specialized apparatus; therefore, each host microhabitat (e.g. specific position on the external or internal organ) is associated with a morphological adaptation, i.e. a haptoral morphotype (Rohde, Reference Rohde1979). Cyprinoid fish serve as hosts for several monogenean genera, of which oviparous and gill-infesting Dactylogyrus is the most species rich (more than 900 nominal species, according to the latest checklist compiled by Gibson et al., Reference Gibson, Timofeeva and Gerasev1996). Dactylogyrus parasites are almost exclusively associated with cyprinoids and their remarkable species diversity is presumably linked to the diversification and phylogeography of their fish hosts, as each cyprinoid species may potentially serve as a host to at least 1 specialist Dactylogyrus species (Ergens, Reference Ergens1970; Šimková and Morand, Reference Šimková and Morand2008; Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018). This is especially evidenced in species diversity hotspots or in regions with a high degree of local endemism (e.g. Dupont and Lambert, Reference Dupont and Lambert1986; El Gharbi et al., Reference El Gharbi, Renaud and Lambert1992, Reference El Gharbi, Birgi and Lambert1994; Rahmouni et al., Reference Rahmouni, Řehulková, Pariselle, Rkhami and Šimková2017; Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018, Reference Benovics, Desdevises, Šanda, Vukić, Scheifler, Doadrio, Sousa- Santos and Šimková2020b). Nonetheless, the highest Dactylogyrus diversity is harboured by host species with wide distribution ranges (Hoffman, Reference Hoffman1999; Seifertová et al., Reference Seifertová, Vyskočilová, Morand and Šimková2008; Musilová et al., Reference Musilová, Řehulková and Gelnar2009; Molnár, Reference Molnár2012). The other highly diverse genus harboured by cyprinoid fish is viviparous Gyrodactylus, whose species are present on fins, skin and gills of their hosts. Currently, more than 400 Gyrodactylus species are described around the world (Harris et al., Reference Harris, Shinn, Cable and Bakke2004), with new species being described almost yearly (e.g. Dos Santos et al., Reference Dos Santos, Maina and Avenant-Oldewage2019; Hansen et al., Reference Hansen, Alvestad, MacKenzie, Darrud, Earlsbakk, Hemmingsen and Arneberg2020; Truter et al., Reference Truter, Smit, Malherbe and Přikrylová2022; Shigoley et al., Reference Shigoley, Rahmouni, Louizi, Pariselle and Vanhove2023). The host specificity in species of Gyrodactylus is comparatively high, as in Dactylogyrus (more than 70% species infect a single host species; Bakke et al., Reference Bakke, Harris and Cable2002), even in spite of the fact that the life cycle lacks free-living larval stages. However, the taxonomy is usually ambiguous, as the most relevant hard parts are only the haptoral ones. This is because the MCOs are often not well recognized, or not developed (Bakke et al., Reference Bakke, Harris and Cable2002; Huyse and Volckaert, Reference Huyse and Volckaert2005).
The study of the diversity of parasites in Iraq can be traced back to the 1970s, to the work of Herzog (Reference Herzog1969), who examined various fishes from markets and identified 4 monogenean species besides endoparasitic helminths. Since then, the vast majority of local research has been conducted only on a small number of targeted fish host species (e.g. Al-Rubaie et al., Reference Al-Rubaie, Hussain and Abdul-Ameer2007; Hussain, Reference Hussain2008; Bashȇ and Abdullah, Reference Bashȇ and Abdullah2010; Mohammad, Reference Mohammad2016), on specific parasite taxa (e.g. Rahemo, Reference Rahemo1980, Reference Rahemo1982; Abdullah and Mhaisen, Reference Abdullah and Mhaisen2005; Al-Ayash et al., Reference Al-Ayash, Gustinelli, Al-Nasiri and Caffara2021) or in restricted geopolitical or hydrological regions (e.g. Al-Shaikh et al., Reference Al-Shaikh, Mhaisen, Al-Khateeb, Balasem and Mansoor1995; Mhaisen, Reference Mhaisen1995; Muhammad et al., Reference Muhammad, Dhahir, Bilal and Abdullah2013; Hashim et al., Reference Hashim, Abdullah and Hassan2015). According to the host–parasite checklists composed for the different Iraq regions (and some additional records; Rasheed and Al-Saadi, Reference Rasheed and Al-Saadi2018), local monogenean fauna comprises more than 107 species belonging to 12 genera (Mhaisen, Reference Mhaisen1995; Mhaisen and Al-Nasiri, Reference Mhaisen and Al-Nasiri2012; Mhaisen and Abdullah, Reference Mhaisen and Abdullah2017; Mhaisen et al., Reference Mhaisen, Al-Rubae and Al-Sa'adi2015, Reference Mhaisen, Al-Mayali and Al-Abodi2019). However, no previous study focusing on the diversity of monogeneans parasitizing fish species in Iraq applied genetic data for taxonomical purposes, nor performed molecular phylogenetic reconstruction. Besides the studies of Koyee and Abdullah (Reference Koyee and Abdullah2019) and Benovics et al. (Reference Benovics, Nejat, Abdoli and Šimková2021a), no genetic data are so far available for Iraq monogeneans. Recently, an integrative approach combining genetic characterization with commonly used, taxonomically important morphological characters has become the gold standard with respect to taxonomical research on monogeneans (e.g. Řehulková et al., Reference Řehulková, Benovics and Šimková2020; Acosta et al., Reference Acosta, Truter, Malherbe and Smit2022; Bahanak et al., Reference Bahanak, Mbondo, Bassock Bayiha, Pariselle, Nack, Bilong Bilong and Agnèse2022; Jin et al., Reference Jin, Li, Cheng, Li, Wu, Zou and Wang2022; Nitta, Reference Nitta2023), as, only by evaluating both morphological and molecular data, can the potential complexes of cryptic species (morphologically indistinguishable and closely related) be revealed, as previously documented for Gyrodactylus (Bueno-Silva et al., Reference Bueno-Silva, Boeger and Pie2011), Cichlidogyrus (Kmentová et al., Reference Kmentová, Gelnar, Mendlová, Van Steenberge, Koblmüller and Vanhove2016) and Dactylogyrus (Rahmouni et al., Reference Rahmouni, Řehulková, Pariselle, Rkhami and Šimková2017; Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018).
Therefore, the present study aimed to employ for the first time such an integrative approach to investigate the species diversity of monogeneans of cyprinoids in Iraq. The newly obtained molecular data for previously and newly described Dactylogyrus and Gyrodactylus species were used to investigate the phylogenetic relationships of endemic parasite taxa to congeners from other geographical regions in order to assess the possible phylogeographical scenarios for cyprinoid hosts and their associated monogeneans. Since for freshwater fauna the Middle East served as a dispersion crossroads between 3 continents (Africa, Asia and Europe), we expected that cyprinoid fish would harbour host-specific parasites phylogenetically associated with congeneric species from all these regions.
Materials and methods
Collection and identification of fish hosts
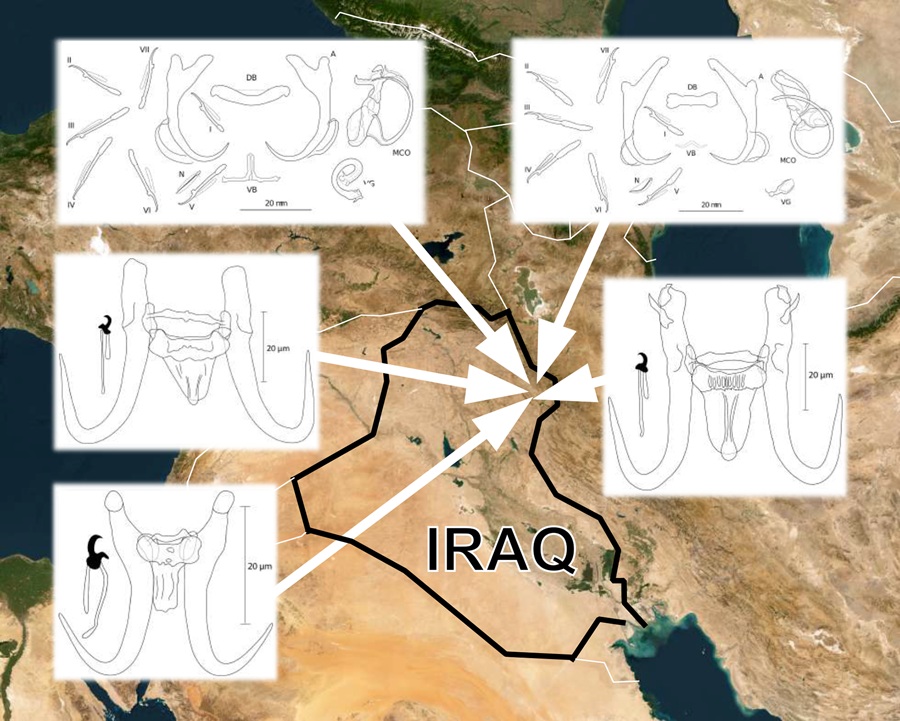
In September 2021, 13 endemic cyprinoid species were surveyed in Iraq for the presence of ectoparasitic monogeneans. A total of 149 fish specimens were collected at 6 localities in northern and north-western Iraq (see Fig. 1, and also Table 1 for coordinates). The number of collected and examined species represented 41% of indigenous cyprinoid taxa according to the recent checklist compiled by Al-Faisal (Reference Al-Faisal2020). The identification of fish was performed by experienced ichthyologists, and the complete cytochrome b (cyt-b) was genotyped to confirm species assignment following polymerase chain reaction (PCR) protocols published by Viñuela-Rodríguez et al. (Reference Viñuela-Rodríguez, Šanda, Zogaris and Vukić2021) (see supplementary Table 1 for GenBank accession numbers).
Map with points showing collection sites in Iraq. The codes at points correspond to locality IDs in Table 1.

List of examined cyprinoid species and collected monogenean parasites with the prevalence of individual parasite species in a population of host

The prevalence is in %. N = number of processed fish specimens per population. Locality IDs correspond to those in Fig. 1.
Collection, fixation, identification and quantification of monogenean parasites
The body surface (including head cavities), fins and gills of freshly killed fishes were examined under a dissection microscope for the presence of ectoparasitic monogeneans, which were collected using fine needles. Parasite collection and fixation followed Řehulková et al. (Reference Řehulková, Seifertová, Přikrylová, Francová, Scholz, Vanhove, Smit, Jayasundera and Gelnar2018). In short, specimens that were subjected to morphological analysis of the hard structures (i.e. haptoral components and copulatory organs – MCO, vagina) were completely flattened under coverslip pressure and fixed with a mixture of glycerine and ammonium picrate (Malmberg, Reference Malmberg1957). For each monogenean species, at least 5 specimens intended for DNA analysis were bisected using fine needles: one-half of the body was fixed in 96% ethanol for DNA extraction; the remaining half (either the posterior part containing the haptoral sclerites of Gyrodactylus spp., or the anterior part with the MCO of Dactylogyrus spp.) was mounted on a slide for further identification and kept as a hologenophore (sensu Pleijel et al., Reference Pleijel, Jondelius, Norlinder, Nygren, Oxelman, Schander, Sunberg and Thollesson2008). Species identification was performed according to the shape and size of the hard elements, following Pugachev et al. (Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009).
Prevalence in the host populations (Table 1), as the percentage of fish infected by a given parasite species, was calculated for each monogenean species, following Bush et al. (Reference Bush, Lafferty, Lotzs and Shostakll1997).
Morphometric data and species description
The mounted monogeneans (or their parts) were studied using an Olympus BX 61 microscope equipped with phase contrast optics. The terminology and measurement procedure for the hard structures adopted here essentially follow those of Malmberg (Reference Malmberg1970) and Pugachev et al. (Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). Measurements of morphometrical characters (in micrometres) were taken using digital image analysis software (StreamMotion, version 1.9.2; Olympus). Meristic data are presented in the tables and are given as means followed by the range in parentheses; the number of specimens measured (in subscript font) is given after the respective parentheses. The dimensions of the body and haptor were obtained from unflattened specimens as the longest body measurements, whereas measurements of the hard structures were taken from completely flattened specimens. Drawings were made with the aid of a drawing attachment and redrawn with a graphics tablet compatible with Adobe Illustrator software. Concerning Dactylogyrus species, the numbering of hook pairs (in Roman numerals I–VII) follows Mizelle (Reference Mizelle1936). Type specimens and hologenophores of the monogeneans studied were deposited in the Helminthological Collection of the Institute of Parasitology of the Czech Academy of Sciences (IPCAS), Czech Republic, under the accession numbers IPCAS M-782–793. To comply with the regulations set out in article 8.5 of the amended 2012 version of the International Code of Zoological Nomenclature (ICZN, 2012), details of the new monogenean species have been submitted to ZooBank.
DNA extraction and amplification
Prior to DNA extraction, the parasites halves were dried from the ethanol using a vacuum centrifuge. Extraction was performed using DNeasy Blood & Tissue kit (Qiagen, Hilden, Germany) following the standard protocol provided by the manufacturer. For Dactylogyrus and Dogielius, 2 DNA fragments were amplified. Specifically, a section of the partial 18S rRNA gene (18S) with the complete internal transcribe spacer 1 region (ITS1), and the partial 5.8 rRNA gene (5.8S) were amplified using either the combination of forward primer S1 and reverse primer IR8 (Šimková et al., Reference Šimková, Plaisance, Matějusová, Morand and Verneau2003), or the combination of S1 and reverse primer Lig5.8R if the former combination was not yielding successful amplification (Šimková et al., Reference Šimková, Plaisance, Matějusová, Morand and Verneau2003; Blasco-Costa et al., Reference Blasco-Costa, Míguez-Lozano, Sarabeev and Balbuena2012). The amplification reactions followed protocols optimized in Benovics et al. (Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018) and Benovics et al. (Reference Benovics, Desdevises, Šanda, Vukić and Šimková2020a). A DNA fragment of the partial 28S rRNA gene (28S) was amplified using forward primer C1 and reverse primer D2 (Hassouna et al., Reference Hassouna, Michot and Bachellerie1984). The amplification reaction for this region followed Benovics et al. (Reference Benovics, Desdevises, Šanda, Vukić and Šimková2020a). For Gyrodactylus, the region containing a fragment of ITS1, complete 5.8S rDNA and partial internal transcribe spacer 2 region (ITS2) was amplified using the combination of the primers ITS1A (forward) and ITS2 (reverse) (Matějusová et al., Reference Matějusová, Gelnar, McBeath, Collins and Cunningham2001a). The amplification reaction followed the protocol optimized by Kvach et al. (Reference Kvach, Ondračková, Seifertová and Hulak2019). For diplozoids, complete ITS2 was amplified using the forward primer D and reverse primer B1 (Bachellerie and Qu, Reference Bachellerie and Qu1993), and the amplification reaction, including PCR conditions, followed the protocol described in Matějusová et al. (Reference Matějusová, Koubková, D'Amelio and Cunningham2001b). The PCR products were checked on 1% agarose gel and subsequently purified using ExoSAP-ITTM (ThermoFisher Scientific, Waltham, MA, USA). Sequencing was performed by Macrogen Europe (Amsterdam, the Netherlands), and was carried out using amplification primers.
Phylogenetic analyses
Phylogenetic analyses were performed separately for each of 2 highly diversified monogenean taxa (Dactylogyrus and Gyrodactylus) to infer the relationships of the newly described species to the congeners. The orthologue sequences of congeneric species were aligned using the fast Fourier transform algorithm employing MAFFT (Katoh et al., Reference Katoh, Misawa, Kuma and Miyata2002) and applying the G-INS-i refinement method. In instances of concatenated sequence datasets, the alignments were treated as partitioned, and an optimal evolutionary model was selected for each partition individually. The phylogenetic analyses were conducted by means of the maximum likelihood (ML) method and Bayesian inference (BI) in RAxML 8.1.12 (Stamatakis, Reference Stamatakis2006, Reference Stamatakis2014) and MrBayes 3.2. (Ronquist et al., Reference Ronquist, Teslenko, van der Mark, Ayres, Darling, Höhna, Larget, Liu, Suchard and Huelsenbeck2012), respectively. For both analyses, all parameters were a priori set free to simulate a general time reversible evolutionary model and without reducing the robustness of heuristic search. This allowed respective algorithms to select the optimal model for DNA evolution over the initial search period. The nodal support in each ML analysis was assessed by simulating 1000 pseudoreplicates. Bayesian analyses were run for 5 000 000 generations, with a tree sampling frequency every 100. After checking that the standard deviation fell under 0.01, the first 30% of samples were discarded as representing an initial burn-in period. The convergence of 2 parallel runs was checked in Tracer 1.7.1. (Rambaut et al., Reference Rambaut, Drummon, Xie, Baele and Suchard2018). Posterior probabilities for each tree node were calculated as the frequency of samples recovering a given clade. The outgroups for each phylogenetic analysis were selected individually to represent phylogenetically sister taxa.
The sequence dataset for assessing phylogenetic relationships of Dactylogyrus spp. was built of concatenated sequences of 18S and 28S. Regions containing ITS1 were omitted from the analyses due to ITS1's hypervariability and problematic alignment when comparing phylogenetically divergent taxa (see Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018, Reference Benovics, Nejat, Abdoli and Šimková2021a, Reference Benovics, Vukić, Šanda, Nejat, Charmpila, Buj, Shumka, Porcelloti, Tarkan, Aksu, Emiroğlu and Šimková2023). Orthologue 18S and 28S sequences from a total of 97 Dactylogyrus species representing all so-far-known (Šimková et al., Reference Šimková, Řehulková, Choudhury and Seifertová2022) phylogenetic lineages were retrieved from the GenBank database (see supplementary Table 2 for metadata and GenBank accession numbers). The species Ancyrocephalus percae (Ergens 1966) was selected as the outgroup for phylogenetic reconstruction, following Mendoza-Palmero et al. (Reference Mendoza-Palmero, Blasco-Costa and Scholz2015).
The alignment for assessing phylogenetic relationships in Gyrodactylus was built of orthologue sequences of the region containing partial ITS1, complete 5.8S and partial ITS2. Sequences from a total of 38 congeneric species were retrieved from the GenBank database (see supplementary Table 3 for metadata and GenBank accession numbers), and Macrogyrodactylus karibae Douëllou and Chishawa, 1995 was selected as outgroup, following Přikrylová et al. (Reference Přikrylová, Vanhove, Janssens, Billeter and Huyse2013). The sequences were carefully selected to represent the individual Gyrodactylus lineages and to concur with the length of the newly generated sequences from the species collected in this study.
Results
Overall diversity of collected monogenean taxa
A total of 33 monogenean species belonging to 4 genera (Dactylogyrus, Dogielius, Gyrodactylus and Paradiplozoon) were collected from the fins and gills of the examined cyprinoid hosts. The prevalence of each parasite species is shown in Table 1. Monogenean communities with the greatest species richness were harboured by Garra rufa (Heckel, 1843) from the Little Zab River, Paracapoeta trutta (Heckel, 1843) from Kani Shok and Luciobarbus barbulus (Heckel, 1847) from the Aw-e Shiler River, where 5 monogenean species were reported. In contrast, populations of Acanthobrama marmid Heckel, 1843, Alburnus sellal Heckel, 1843 (at the Grdi Go collection site), Alburnus sp., Carasobarbus luteus Heckel, 1843 (from the Tarbin River) and Squalius berak harboured only 2 monogenean species each. The most species-diverse genus was Dactylogyrus (Dactylogyridae) with 16 species, followed by Gyrodactylus (Gyrodactylidae) with 12 species. Only 4 species of Dogielius (Dactylogyridae) were recorded on the examined cyprinoids, each from a single endemic host species. Dogielius molnari Jalali, 1992 and D. mokhayeri Jalali and Molnár, Reference Jalali and Molnár1990 were collected from Cyprinion macrostomum Heckel, 1843 and P. trutta, respectively. A potentially new species for science, Dogielius sp., was collected from Capoeta umbla (Heckel, 1843) at wadi Kalat Shirah. Dogielius cf. persicus Molnár and Jalali, Reference Molnár and Jalali1992 was collected from C. luteus – however, only at the Grdi Go site, where only 2 C. luteus specimens were collected and examined. The last reported monogenean genus was Paradiplozoon (Diplozoidae), with a single representative, Paradiplozoon homoion Bychowsky and Nagibina, 1959, which was recorded from 10 of the investigated cyprinoid species. The highest prevalence of P. homoion was recorded on A. sellal at Grdi Go (P = 70%). Dactylogyrus and Gyrodactylus species diversity and phylogeny, and descriptions of the new species are presented in the subsequent sections below.
Species diversity of Dactylogyrus parasites in Iraq
A total of 12 of the 13 investigated cyprinoid species were parasitized by Dactylogyrus species. Eight cyprinoid species were parasitized by a single Dactylogyrus species, whilst the remaining 4 cyprinoid species were parasitized by 2 or 3 Dactylogyrus species. A total of 16 Dactylogyrus species were identified. The majority of Dactylogyrus species were recorded only on a single host species. Only Dactylogyrus holciki Molnár and Jalali, Reference Molnár and Jalali1992 was collected from 2 congeneric hosts – A. sellal and Alburnus sp. The prevalence of D. holciki differed between 2 populations of A. sellal. Nonetheless, its prevalence also differed between 2 Alburnus species from the same locality (Table 1). The highest number of Dactylogyrus species (3) were recorded on L. barbulus at Du-Choman (the Aw-e Shiler River).
Four new Dactylogyrus species were found on the gills of endemic cyprinoids and are described below. Each of them was collected from only a single host species (i.e. A. marmid, Chondrostoma regium (Heckel, 1843), G. rufa and Squalius lepidus Heckel, 1843). Except for Dactylogyrus medicus n. sp. from G. rufa, all other new species were obtained from the same site on the Aw-e Shiler River.
Phylogenetic relationships of Dactylogyrus in Iraq
The final concatenated nucleotide alignment comprising partial 18S and 28S included 110 sequences of 105 Dactylogyrus species (4 previously published conspecific sequences were used to confirm the identity of newly collected species) and spanned 1148 unambiguously aligned nucleotide positions (429 bp for 18S; 719 bp for 28S). Both phylogenetic analyses (BI and ML) generated trees with identical topologies and differed only in their nodal support (see tree generated by BI in Fig. 2).
Phylogenetic tree of 105 Dactylogyrus spp. parasitizing various cyprinoid fish hosts. The tree is based on 111 combined sequences of partial genes coding 18S and 28S rRNA, and rooted using Ancyrocephalus percae. Values at the nodes indicate posterior probabilities from BI and bootstrap values from ML analyses. Dashes indicate values below 0.70 and 50, respectively. Letters (A–E) represent specific well-supported clades. The newly described and newly reported species from this study are in red.

The phylogenetic reconstruction divided all Dactylogyrus species into 5 well-supported lineages. Lineage A included 10 Dactylogyrus species collected in this study, which were in paraphyly. Our results suggest that 4 North American Dactylogyrus species (i.e. D. parvicirrus Seamster, 1948, D. flagristylus Chien, 1974, D. cheloideus Rogers, 1967 and D. pectenatus Mayes, 1977) form a nested group within lineage A; however, the position of this group to other congeners within lineage A was not well resolved. Dactylogyrus anoigeus n. sp. from A. marmid, and D. rivalis n. sp. from S. lepidus were revealed to be phylogenetically close to D. folkmanovae Ergens, 1956 and D. vranoviensis Ergens, 1956, both common species of Squalius spp. in Europe and the Middle East. Dactylogyrus regius n. sp., described from C. regium, grouped with common species of Chondrostoma and Parachondrostoma (the Chondrostoma sensu lato group) in Europe and the Middle East, namely D. ergensi Molnár, 1964, D. dirigerus Gussev, 1966, D. conchatus Benovics, Francová, Volta, Dlapka and Šimková, 2021 and D. globulatus Benovics, Francová, Volta, Dlapka and Šimková, 2021. All these species share remarkable similarities in their hard taxonomically important characters with D. sagittarius Benovics, Francová, Volta, Dlapka and Šimková, 2021, D. caucasicus Mikailov and Shaova, 1973, D. rutili Glaser, 1965 and D. tissensis Zachvatkin, 1951. Lineage B included all other analysed North American Dactylogyrus species. Lineage C included Dactylogyrus species originating in eastern and southeastern Asia, associated with C. carpio and Carassius spp. fishes, Indonesia (i.e. D. tapienensis Chinabut and Lim, 1993 and D. viticulus Chinabut and Lim, 1993), North Africa (D. marocanus El Gharbi, Birgi and Lambert, Reference El Gharbi, Birgi and Lambert1994) and the Middle East (i.e. D. acinacus Gussev, Jalali and Molnár, Reference Gussev, Jalali and Molnár1993, D. pulcher Bychowsky, 1957 and the newly described D. medicus n. sp. from G. rufa). Lineage D included only 2 species collected in Iraq – specifically, D. microcirrus Gussev, Jalali and Molnár, Reference Gussev, Jalali and Molnár1993, originally described from C. trutta (syn Paracapoeta trutta [Heckel, 1843]) in Iran, and D. macrostomi Gussev, Ali, Abdul-Ameer, Amin and Molnár, 1993, described from C. macrostomum, also in Iran. Finally, lineage E encompassed species parasitizing cyprinoids in the western peri-Mediterranean region and 2 Middle Eastern species, which were present also in Iraq (i.e. D. carassobarbi Gussev, Jalali and Molnár, Reference Gussev, Jalali and Molnár1993 and D. lenkorani Mikailov, 1974).
Morphological and molecular characterization of the new Dactylogyrus species
Dactylogyrus anoigeus Řehulková n. sp. (Fig. 3)
Type-host: Acanthobrama marmid Heckel, 1843 (Cyprinoidei: Leuciscidae).
Hard structures of Dactylogyrus anoigeus n. sp. ex Acanthobrama marmid. A, anchor; DB, dorsal bar; VB, ventral bar; N, needle; I–VII, hooks; VG, vagina; MCO, male copulatory organ.

Type-locality: Du Choman, the Aw-e Shiler River, Sulaymaniyah Province, Iraq.
Site on host: Gill filaments.
Type-material: Holotype, 2 paratypes, 4 hologenophores (IPCAS M-790).
Representative DNA sequence: A nucleotide sequence of the partial gene for 28S rRNA (733 bp long; OR817682), and nucleotide sequences representing a fragment (955 bp long; OR817699) including the partial gene for 18S rRNA (488 bp), and the ITS1 region (467 bp). No intraspecific variability was found.
Infection indices: Prevalence 70%, 1–4 monogeneans per infected host.
Etymology: The specific name is from Greek (anoigeus = opener) and refers to the shape of the distal part of the accessory piece of the MCO.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:EB687773-3B01-4741-B6D0-02CDFE4DF023.
Description: (Dimensions of the hard structures are given in Table 2.) Two pairs of anchors with roots of similar lengths (inner root slightly longer, with flattened termination), elongate shaft bent near its proximal third, recurved point not well demarcated from the shaft and extending well past level of tip of inner root. Dorsal bar broadly V-shaped, weakly narrowed medially, with anteromedial inconspicuous membrane. Ventral bar vestigial, inverted T-shaped, 3-armed. One pair of needles located near hooks of pair V. Seven pairs of hooks; each with delicate point, truncate thumb and shank inflated along proximal 1/3; filamentous hook (FH) loop extending to near level of shank inflation. MCO composed of basally articulated copulatory tube and accessory piece. Copulatory tube with base angularly demarcated from C-shaped shaft. Accessory piece appearing as a plaited rod encircling partially the base and distally formed as a tin opener guiding the end of the tube. Vagina a curled short tube.
Morphometric data for newly described Dactylogyrus species

The first number represents the mean value and is followed by the range of obtained measurements in brackets. The lower index number represents the number of measured specimens.
Differential diagnosis: Dactylogyrus anoigeus n. sp. belongs to the group of congeners having an inverted T-shaped ventral bar and an MCO between the ʻnanus' and ʻchondrostomi' types (see Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). It most closely resembles D. folkmanovae, a parasite of Squalius cephalus (Linnaeus, 1758) (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009), in the comparative morphology of the ventral bar and MCO. In both species, all 3 processes of the ventral bar are similar in length, and each has the same diameter throughout its length, but in D. anoigeus n. sp., the termination of the anterior process is rounded (vs flattened in D. folkmanovae). The MCO of the 2 species is characterized by a sickle-shaped copulatory tube with a recurved base and an accessory piece with a distal widening formed as 2 parts, of which 1 serves as a guide for the distal termination of the tube (pincer-shaped in D. anoigeus n. sp. vs finger-shaped, with a subterminal filament in D. folkmanovae), while the other is directed backwards along the distal curvature of the tube (filamentous in D. anoigeus n. sp. vs more robust claw-shaped in D. folkmanovae). In addition to the above differences, D. anoigeus n. sp. is easily differentiated from D. folkmanovae by having anchors with roots of similar size (the inner root is markedly longer than the outer root in D. folkmanovae) and a recurved point (vs open point in D. folkmanovae). The sister relationship between D. anoigeus n. sp. and D. vranoviensis Ergens, 1956 was supported (albeit weakly by ML) by molecular phylogeny. The 2 species share a similar MCO morphology (i.e. a sickle-shaped copulatory tube and an accessory piece encircling partially the base of the tube in the form of a finger-like process), but they clearly differ in that the new species has anchors with a well-developed point (vs markedly reduced point in D. vranoviensis) and an inverted T-shaped ventral bar (vs vestigial rod-shaped ventral bar in D. vranoviensis).
Until now, only 2 species of Acanthobrama have been reported as hosts for species of Dactylogyrus: A. terraesanctae (now Mirogrex terraesanctae [Steinitz, 1952]) for D. acanthobramae Paperna, Reference Paperna1961, D. carmeli Paperna, Reference Paperna1961 and D. sphyrna Linstow, 1878 (Paperna, Reference Paperna1961, Reference Paperna1964), and A. simoni (now Pseudobrama simoni [Bleeker, 1864]) for D. acanthobramis Zhang and Ji, Reference Zhang and Ji1980 and D. jiayuensis Zhang and Ji, Reference Zhang and Ji1980 (Zhang and Ji, Reference Zhang and Ji1980). Of the 5 Dactylogyrus species, D. anoigeus n. sp. is most similar to D. acanthobramae, as the MCO of the 2 species appears to have some common features. Although Paperna's (Reference Paperna1961) drawing of the MCO is confusing and not strongly diagnostic, the copulatory tube is depicted and described as ‘winding’, and the medial part of the accessory piece appears to be markedly thinner than in D. anoigeus n. sp. Considering the different host (M. terraesanctae) and locality (Israel, Lake Galilee) recorded for D. acanthobramae, we do not consider these 2 species conspecific; however, D. acanthobramae requires redescription that should be based on new specimens collected from its type host and the type locality.
Dactylogyrus medicus Řehulková n. sp. (Fig. 4)
Type- host: Garra rufa (Heckel 1843) (Cyprinoidei: Cyprinidae).
Hard structures of Dactylogyrus medicus n. sp. ex Garra rufa. A, anchor; DB, dorsal bar; VB, ventral bar; N, needle; I–VII, hooks; VG, vagina; MCO, male copulatory organ.

Type-locality: by the road Suleymania-Dukan, Little Zab, Sulaymaniyah Province, Iraq.
Site on host: Gill filaments.
Type-material: Holotype, 2 paratypes, 2 hologenophores (IPCAS M-791).
Representative DNA sequence: A nucleotide sequence of the partial gene for 28S rRNA (729 bp long; OR817691), and nucleotide sequences representing a fragment (961 bp long; OR817710) including the partial gene for 18S rRNA (467 bp), and the ITS1 region (494 bp). No intraspecific variability was found.
Infection indices: Prevalence 60%, 1–6 monogeneans per infected host.
Etymology: The specific name refers to the fish host G. rufa, also known as the doctor fish.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:6FAD82DA-C64D-4C48-A894-E6DC57FB340E.
Description: (Dimensions of the hard structures are given in Table 2.) Two pairs of anchors with elongate terminally tapering inner root, moderately developed outer root, proximally slightly swollen shaft and recurved point reaching level of tip of inner root. Dorsal bar straight, bone-shaped, with enlarged slightly indented ends. Ventral bar vestigial, poorly defined or absent, resembling an inverted flying bird symbol. One pair of needles located near hooks of pair V. Seven pairs of hooks, each with delicate point, protruded thumb and shank inflated along proximal half; FH loop extending to near level of shank inflation. MCO composed of basally articulated copulatory tube and accessory piece. Copulatory tube comprising bulbous base with flange and usually number 6-shaped shaft. Accessory piece articulated just posteriorly to base at level of the basal flange, closed leaf-shaped, serving as a guide for distal part of the tube. Vagina inconspicuous, lightly sclerotized, variable in shape.
Differential diagnosis: Dactylogyrus medicus n. sp. represents the fifth species of Dactylogyrus besides D. tylognathi Paperna, Reference Paperna1961, D. garrae Paperna, Reference Paperna1964 (Israel; Paperna, Reference Paperna1961, Reference Paperna1964), D. acinacus Gussev et al., Reference Gussev, Jalali and Molnár1993 and D. rectotrabus (Iran, Turkey; Gussev et al., Reference Gussev, Jalali and Molnár1993; Koyun, Reference Koyun2011) so far recorded on the doctor fish, G. rufa. A further 3 species of Dactylogyrus parasitizing species of Garra, i.e. D. lingualis Lang, Reference Lang1981, D. onychocirrus Lang, Reference Lang1981 and D. spirotubivagina Ann and Zang, 1988, have been described from Garra orientalis in China (Lang, Reference Lang1981; Ann and Zhang, Reference Ann and Zhang1988). The haptoral configurations of all the Dactylogyrus species parasitizing Garra spp. show common features such as a rod-shaped dorsal bar, a missing or vestigial ventral bar, and anchors of the pseudanchoratus type (see Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009), which is characterized by a long inner root and short outer root, a swelling on the shaft and a point that is not well demarcated from the shaft (vs angularly demarcated from the shaft in the anchoratus type). Dactylogyrus medicus n. sp. is clearly differentiated from D. garrae and D. tylognathi parasitizing G. rufa and from all known Dactylogyrus spp. reported from G. orientalis (i.e. D. lingualis, D. onychocirrus and D. spirotubivagina) by having a shorter copulatory tube of the MCO (the copulatory tube is markedly longer and thinner, meandering or coiled in the 5 respective species). In this respect, D. medicus n. sp. is similar to D. acinacus and D. rectotrabus, which possess a relatively short J-shaped copulatory tube with a bulbous base. However, it clearly differs from the above 2 congeners by having an MCO with an accessory piece resembling a closed leaf (an accessory piece in the form of 1 or more rod-shaped plates placed in parallel to the copulatory tube). In addition, unlike in D. acinacus and D. rectotrabus, the ventral bar is present in D. medicus n. sp, although is barely visible, even under phase contrast optics.
Dactylogyrus spp. parasitizing G. rufa (i.e. D. acinacus and D. medicus n. sp.) are morphologically similar to Dactylogyrus marocanus (Fig. 2), a phylogenetically closely related parasite with a broad host range including torins and barbins in Morocco, as was previously suggested also by Řehulková et al. (Reference Řehulková, Rahmouni, Pariselle and Šimková2021).
Dactylogyrus regius Řehulková n. sp. (Fig. 5)
Type-host: Chondrostoma regium (Heckel 1843) (Leuciscidae: Leuciscinae).
Hard structures of Dactylogyrus regius n. sp. ex Chondrostoma regium. A, anchor; DB, dorsal bar; VB, ventral bar; N, needle; I–VII, hooks; VG, vagina; MCO, male copulatory organ.

Type-locality: Du Choman, the Aw-e Shiler River, Sulaymaniyah Province, Iraq.
Site on host: Gill filaments.
Type-material: Holotype, 2 paratypes, 2 hologenophores (IPCAS M-792).
Representative DNA sequence: A nucleotide sequence of the partial gene for 28S rRNA (703 bp long; OR817693), and nucleotide sequences representing a fragment (845 bp long; OR817707) including the partial gene for 18S rRNA (467 bp), and the ITS1 region (378 bp). No intraspecific variability was found.
Infection indices: Prevalence 83%, 1–8 monogeneans per infected host.
Etymology: The specific name refers to the fish host.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:306AB571-A70D-4069-A964-DE265945723C.
Description: (Dimensions of hard structures are given in Table 2.) Two pairs of anchors with moderately long and terminally flattened inner root, rounded outer root and evenly curved shaft and point, point with slightly recurved tip and extending well past level of tip of inner root. Dorsal bar nearly yoke-shaped, with anteromedial depression. Ventral bar vestigial, inverted T-shaped, 3-armed. One pair of needles located near hooks of pair V. Seven pairs of hooks; each with delicate point, flattened thumb and shank inflated along proximal half; FH loop extending to near level of shank inflation. MCO composed of basally articulated copulatory tube and accessory piece. Copulatory tube with saclike base recurved posteriorly; shaft elongated, thin, nearly C-shaped. Accessory piece attached to base of tube as 2 filaments (1 markedly thinner) and formed distally as a plate-like sheath giving rise to tongue-shaped lobe directed backwards along the circle of the curved tube. Vagina a relatively long tube of variable course, with lobed ends.
Differential diagnosis: Dactylogyrus regius n. sp. belongs to the group of congeners having the MCO of the ʻchondrostomi' type, which is characterized by an accessory piece with a tongue-shaped lobe directed backwards along the circle of the curved copulatory tube. This morphological group includes parasites mostly of Chondrostoma hosts (e.g. D. dirigerus, D. ergensi and D. globulatus), Telestes hosts (e.g. D. conchatus and D. sagittarius) and Alburnoides hosts (e.g. D. caucasicus and D. tissensis) (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009; Benovics et al., Reference Benovics, Francová, Volta, Dlapka and Šimková2021b), which clustered together in the phylogenetic tree (Fig. 2). Dactylogyrus regius n. sp. differs from other congeners in the cluster by having the following combination of characters: sabre-shaped anchors, an inverted T-shaped ventral bar and an accessory piece of the MCO with a robust distal part appearing as plate-like sheath.
Dactylogyrus regius n. sp. most closely resembles D. elegantis (not included in our phylogenetic analyses), a parasite of C. knerii, C. nasus and C. regium (Stojanovski et al., Reference Stojanovski, Kulišić, Baker, Hristovski, Cakić and Hristovski2004; Koyun, Reference Koyun2011; Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018) in the comparative morphology of their haptoral structures. In both species, the anchors possess an evenly curved shaft and point (sabre-shaped type), straight to a broadly V-shaped dorsal bar with an anteromedial depression, and an inverted T-shaped ventral bar. Dactylogyrus regius n. sp. clearly differs from D. elegantis by having an accessory piece with a robust distal part appearing as a plate-like sheath through which the distal end of the copulatory tube passes (distal part simple, with groove in D. elegantis) and a proximal part formed as 2 filaments (proximal part appearing as a more compact bifurcated rod in D. elegantis).
Dactylogyrus rivalis Řehulková n. sp. (Fig. 6)
Type-host: Squalius lepidus Heckel 1843 (Cyprinoidei: Leuciscidae).
Hard structures of Dactylogyrus rivalis n. sp. ex Squalius lepidus. A, anchor; DB, dorsal bar; VB, ventral bar; N, needle; I–VII, hooks; VG, vagina; MCO, male copulatory organ.

Type-locality: Du Choman, Aw-e Shiler River, Sulaymaniyah Province, Iraq.
Site on host: Gill filaments.
Type-material: Holotype, 3 paratypes, 3 hologenophores (IPCAS M-793).
Representative DNA sequence: A nucleotide sequence of the partial gene for 28S rRNA (730 bp long; OR817698), and nucleotide sequences representing a fragment (995 bp long; OR817715) including the partial gene for 18S rRNA (487 bp), the ITS1 region (488 bp) and 5.8S region (20 bp). No intraspecific variability was found.
Infection indices: Prevalence 44%, 9–22 monogeneans per infected host.
Etymology: The specific name (an adjective) is from Latin (rivalis = a rival, competing) and refers to the co-occurrence of the new species with D. vistulae on the gills of S. lepidus.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:206B6BE8-3767-419F-9662-48E1189F8D6B.
Description: (Dimensions of the hard structures are given in Table 2.) Two pairs of anchors with elongate and terminally flattened inner root, rounded outer root, markedly elongate shaft bent at its proximal third and short point not well demarcated from shaft and extending past level of tip of inner root. Dorsal bar nearly yoke-shaped, with anteromedial depression. Ventral bar vestigial, inverted T-shaped, 3-armed; anteromedial arm with lightly sclerotized termination; lateral arms resembling wings. One pair of needles located near hooks of pair V. Seven pairs of hooks; each with delicate point, flattened thumb and shank inconspicuously inflated along proximal half; FH loop extending to near level of shank inflation. MCO composed of basally articulated copulatory tube and accessory piece. Copulatory tube comprising elongate base and distally tapering sinusoidal shaft. Accessory piece bifurcated near midpoint into rod-shaped arm articulating to base of tube and membranous flap following convex curve of proximal half of the tube. Vagina a curled short tube.
Differential diagnosis: Until now only 3 species of Dactylogyrus have been recorded on S. lepidus, namely D. dyki Ergens and Lucký, 1959, D. elegantis and D. vistulae Prost, 1957 (Iraq; Abdullah and Abdullah, Reference Abdullah and Abdullah2013). However, the microphotograph and drawings of the haptoral structures of D. dyki presented by the above authors show the anchors as having an elongate shaft markedly bent at its proximal third and poorly demarcated from the short point, which does not correspond to those originally described for D. dyki (anchors with moderately long shaft angularly demarcated from long point, i.e. the ʻwunderi' type in Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). The configuration and morphology of the haptoral structures in D. dyki of Abdullah and Abdullah (Reference Abdullah and Abdullah2013), however, corresponds well to that in our specimens of D. rivalis n. sp. In addition, the measurements and overall morphology of the MCO and vagina reported by these authors match those of D. rivalis n. sp. Thus, considering that both parasites were found on the same host species and in close geographical proximity in Iraq, it is probable that the specimens identified by Abdullah and Abdullah (Reference Abdullah and Abdullah2013) as D. dyki were actually the new species described here as D. rivalis n. sp.
Dactylogyrus rivalis n. sp. shares a similar morphology of the dorsal and ventral bar with Dactylogyrus spp. occupying the same clade in the phylogenetic tree (see Fig. 2). Of these species, it most closely resembles D. folkmanovae in having anchors with a markedly elongate shaft bent at its proximal third and a short point not well demarcated from the shaft. It clearly differs from the latter species in possessing a sinusoidal copulatory tube (vs sickle-shaped copulatory tube in D. folkmanovae) supported by membranous accessory piece (vs more compact and distally bifurcated accessory piece in D. folkmanovae).
Species diversity of Gyrodactylus parasites in Iraq
The diversity of the genus Gyrodactylus was found to be poorer when compared to Dactylogyrus diversity. Gyrodactylus spp. were collected from the gills, fins and skin of fish. Ten out of 13 cyprinoid species were parasitized by Gyrodactylus spp. A total of 12 Gyrodactylus species were identified and all of them were recognized as new to science, according to the autapomorphies in taxonomically important morphological characteristics and molecular phylogeny. The highest Gyrodactylus species diversity was recorded from A. sellal, collected from 3 localities; this host species was parasitized by 3 Gyrodactylus species at 3 localities (a maximum of 2 species were found from a single collection site, see below and Table 1). Two new Gyrodactylus species were collected from G. rufa, i.e. G. satanicus n. sp., and G. vukicae n. sp., with the former one exhibiting the highest prevalence across all collected Gyrodactylus species (P = 70%). The G. iraqemembranatus n. sp., which exhibited the widest host range among congeners, was collected from A. sellal (at 2 collection sites), and also from Barbus lacerta Heckel, 1843 and P. trutta (Heckel, 1843).
Due to there being insufficient material for morphological analyses of 4 species (i.e. a low number of mounted Gyrodactylus specimens or the presence of malformed taxonomically important characters), we properly describe here only 8 species below out of a total of 12. For 3 species, insufficient material was available for studying the morphology (i.e. Gyrodactylus sp. 2 from L. barbulus, Gyrodactylus sp. 3 from A. sellal and Gyrodactylus sp. 4 from B. lacerta); therefore, only orthologue sequences were deposited in GenBank, and were also included in the phylogenetic analyses. For the last species, Gyrodactylus sp. 1 collected from P. trutta, no DNA sequence data are available; therefore, it is only mentioned as ‘recorded’.
Phylogenetic relationships of Gyrodactylus in Iraq
The final concatenated nucleotide alignment comprising ITS1, 5.8S and ITS2 regions included 51 sequences of 49 Gyrodactylus species (for G. mhaiseni n. sp. and G. vukicae n. sp., 2 genetic variants were recorded and included in analyses) and spanned 858 unambiguously aligned nucleotide positions (285 bp for ITS1; 162 bp for 5.8S; 411 bp for ITS2). Both phylogenetic analyses (BI and ML) generated trees with identical topologies and differed only partially in their nodal support (see the tree generated by BI in Fig. 7).
Phylogenetic tree of 49 Gyrodactylus spp. parasitizing various fish hosts. The tree is based on 52 combined sequences of partial ITS1 and ITS2 regions with 5.8S rRNA, and rooted using Macrogyrodactylus karibae. Values at the nodes indicate posterior probabilities from BI and bootstrap values from ML analyses. Dashes indicate values below 0.70 and 50, respectively. Letters (A–F) represent specific well-supported clades or lineages. The newly described species from this study are in red.

The phylogenetic analyses revealed that the investigated Gyrodactylus species belonged to 6 well-supported lineages. Lineage A encompassed 9 new Gyrodactylus species, congeners from Europe and also G. kobayashii Hukuda, 1940 parasitizing C. auratus from China and G. hildae García-Vásquez, Hansen, Christison, Bron and Shinn, 2011 parasitizing Oreochromis niloticus (Linnaeus, 1758) from Ethiopia. While the phylogenetic positions of Gyrodactylus sp. 1 from L. barbulus, G. azeezsaeedi n. sp., G. vukicae n. sp., G. satanicus n. sp. and G. jurajdai n. sp. within lineage A were not fully resolved, the other 4 newly described species formed a well-supported monophyletic group. Minor intraspecific variability was observed at the geographical level (for G. mhaiseni n. sp.) and the host species level (for G. vukicae n. sp.).
Lineage B included Gyrodactylus species parasitizing African freshwater fish of Cichlidae. The species belonging to lineage C were monogeneans of Palearctic (G. arcuatus Bychowsky, 1933, G. branchicus Malmberg, 1964, G. nipponensis Ogawa and Egusa, 1978, G. rhodei Žitňan, 1964 and G. pterygialis Bychowsky and Polyansky, 1953), Nearctic (G. mediotorus King, Marcogliese, Forest, McLaughlin and Bentzen, 2013) and Neotropic (G. poeciliae Harris and Cable, 2000) fish hosts. Gyrodactylus blazeki n. sp. grouped together with common Holarctic species within the lineage D. Lineage E encompassed 4 Gyrodactylus species parasitizing African silurids. Gyrodactylus iraqemembranatus n. sp., differing from other congeners by the morphology of taxonomically important characters (see above), has an unresolved relationship to the monophyletic group including lineages A–C and the monophyletic group including lineages D and E.
Morphological and molecular characterization of the new Gyrodactylus species
Gyrodactylus azeezsaeedi Rahmouni n. sp. (Fig. 8)
Type-host: Squalius berak Heckel, 1843 (Cyprinoidei: Leuciscidae)
Hard structures of haptor of Gyrodactylus azeezsaeedi n. sp. ex Squalius berak.

Type-locality: Kani Shok, a tributary of the Tabin River, Sulaymaniyah Province, Iraq
Site on host: Fins
Type material: Holotype and 2 paratypes (IPCAS M-782).
Representative DNA sequence: A nucleotide sequence representing a fragment (1274 bp long; OR773093) including the partial ITS1 region (684 bp long), 5.8S rDNA (157 bp) and the partial ITS2 region (433 bp). No intraspecific variability was found.
Infection indices: Prevalence 10%, 1–5 monogeneans per infected host.
Etymology: The specific name ‘azeezsaeedi’ honours Dr Mohammed Azeez Saeed, the coordinator of international cooperation at Salahaddin University (Erbil, Iraq), for his precious help with the organization of the field trip and hospitality.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:5139D2BB-0B0A-45F6-8D80-76F88C41D9E4.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner roots with hat-like cover; base with posterior folds; root relatively long and straight; shaft slightly bowed; point curved and elongated. Ventral bar with blunt and short bilateral processes extending out of bar; median part may show a hollow; membrane slightly trapezoid, almost 2/3 length of hamuli shaft, with striations ending posteriorly in a median ridge. Dorsal bar simple, with projections at halfway point and attenuated ends inserted into terminal plates. Marginal hooks with a flat base, globose heel slightly curved outward, conspicuous finger-like toe inward, conspicuous shelf, curved point, and sickle proper approximately perpendicular to terminal edge of toe, gently curved downward to a point slightly exceeding the toe; filament loop extending about 1/2 handle length. MCO not observed.
Morphometric data for newly described Gyrodactylus species

The first number represents the mean value and is followed by the range of obtained measurements in brackets. The lower index number represents the number of measured specimens.
Differential diagnosis: Herein, Gyrodactylus from S. berak was identified for the first time. The haptoral morphology exhibited by G. azeezsaeedi n. sp. resembles that of G. gobii (Schulman, 1953) parasitizing widespread Gobio gobio (Linnaeus, 1758); G. leucisci Žitňan, 1964 and G. osoblahensis Ergens, 1963, mostly parasitizing Leuciscus leuciscus (Linnaeus, 1758) and S. cephalus; and, finally, G. scardiniensis Glaser, 1974 from Scardinius erythrophthalmus and Scardinius cephalus (Ergens, Reference Ergens1991, Reference Ergens1992; Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). The new species differs from G. gobii by the shape and size of its dorsal bar, which shows projections at the halfway point, a feature missing in G. gobii, and which is comparatively longer in G. azeezsaeedi n. sp. Gyrodactylus azeezsaeedi n. sp. is distinguishable from G. leucisci in having (i) shorter hamuli (54.8–57.3 μm in G. azeezsaeedi n. sp. vs 63.0–73.0 μm in G. leucisci), (ii) a longer dorsal bar (25.7–27.3 μm in G. azeezsaeedi n. sp. vs 19.0–21.0 μm in G. leucisci) and (iii) shorter marginal hooks (23.5–25.2 μm in G. azeezsaeedi n. sp. vs 30.0–31.0 μm in G. leucisci). Gyrodactylus azeezsaeedi differs from G. osoblahensis in having (i) shorter hamuli (54.8–57.3 μm in G. azeezsaeedi n. sp. vs 60.0–70.0 μm in G. osoblahensis), (ii) a shorter ventral bar (24.0–26.1 μm in G. azeezsaeedi n. sp. vs 29.0–33.0 μm in G. osoblahensis) associated to (iii) a longer membrane (14.0–15.9 μm in G. azeezsaeedi n. sp. vs 22–26 μm in G. osoblahensis) and finally (iv) shorter marginal hooks (23.5–25.2 μm in G. azeezsaeedi n. sp. vs 31.0–39.0 μm in G. osoblahensis). It is further discriminated from G. scardiniensis by its (i) shorter hamuli (54.8–57.3 μm in G. azeezsaeedi n. sp. vs 60.0–70.0 μm in G. scardiniensis), and (ii) shorter marginal hooks (23.5–25.2 μm in G. azeezsaeedi n. sp. vs 32.0–38.0 μm in G. scardiniensis).
Gyrodactylus blazeki Rahmouni n. sp. (Fig. 9)
Type-host: Alburnus sp. (Cyprinoidei: Cyprinidae)
Hard structures of haptor of Gyrodactylus blazeki n. sp. ex Alburnus sp.

Type-locality: Grdi Go, Zalm stream, Sulaymaniyah Province, Iraq
Site on host: Gill filaments
Type material: Holotype and 1 paratype (IPCAS M-783).
Representative DNA sequence: A nucleotide sequence representing a fragment (895 bp long; OR773085) including the partial ITS1 region (347 bp long), 5.8S rDNA (157 bp) and the partial ITS2 region (391 bp). No intraspecific variability was found.
Infection indices: Prevalence 5%, 3 monogeneans per infected host.
Etymology: The specific name ‘blazeki’ honours the ichthyologist Dr Radim Blažek from the Institute of Vertebrate Biology, Czech Academy of Sciences (Brno, Czech Republic) in recognition of his past research on Gyrodactylus.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:2F14F7A0-277D-4DA3-9224-9AC9EAE84E42.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner roots with hat-like cover; base with no posterior folds; root relatively long; shaft slightly bowed; point curved and elongated. Ventral bar lacking bilateral processes; median part of a common width may show holes; membrane slightly rectangular, almost 1/3 length of hamuli shaft, with striations. Dorsal bar simple, with posteriorly directed projections and attenuated ends inserted into terminal plates. Marginal hooks with prominent globose heel, curved finger-like toe downward, conspicuous shelf, curved point, sickle proper curved downward to a point slightly exceeding the toe; filament loop extending about handle length. MCO not observed.
Differential diagnosis: This study presents the first data on monogeneans parasitizing Alburnus spp. from the Middle East. So far, no Gyrodactylus species with similar haptoral morphology has been reported in the Middle East. The overall morphology exhibited by G. blazeki n. sp. resembles that of G. laevis Malmberg, Reference Malmberg1957 and G. prostae Ergens, 1963, both known from a range of Palearctic cyprinids (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). This resemblance is seen in the shape of the hamuli with well-developed roots with folds, the ventral bar lacking bilateral processes and marginal hooks with a well-developed heel. Compared to G. laevis, G. blazeki n. sp. possesses a shorter ventral bar membrane (8.3–8.6 μm in G. blazeki n. sp. vs 9–16 μm in G. laevis). Gyrodactylus blazeki n. sp. differs from G. prostae in having (i) shorter hamuli (31.6–32.1 μm in G. blazeki n. sp. vs 44.0–60.0 μm in G. prostae), (ii) shorter ventral bar membrane (8.3–8.6 μm in G. blazeki n. sp. vs 12.0–16.0 μm in G. prostae) and (iii) shorter marginal hooks (19.9–20.5 μm in G. blazeki n. sp. vs 24.0–30.0 μm in G. prostae).
Gyrodactylus iraqemembranatus Rahmouni n. sp. (Fig. 10)
Type-host: Paracapoeta trutta (Heckel, 1843) (Cyprinoidei: Cyprinidae)
Hard structures of haptor of Gyrodactylus iraqemembranatus n. sp. ex Paracapoeta trutta (A), ex Alburnus sellal (B), ex Barbus lacerta (C).

Additional hosts: Alburnus sellal Heckel, 1843 (Cyprinoidei: Leuciscidae), Barbus lacerta Heckel, 1843 (Cyprinoidei: Cyprinidae)
Type-locality: Kani Shok, tributary of Tabin River, Sulaymaniyah Province, Iraq
Additional locality: wadi Kalat Shirah, a tributary of the Tabin River, and the Tabin River in Zahrzi for A. sellal; Kani Shok, a tributary of the Tabin River also for B. lacerta, all localities in Sulaymaniyah, Iraq
Site on host: Gill filaments for P. trutta and B. lacerta, fins for A. sellal.
Type material: Holotype and 6 paratypes (IPCAS M-784/1-3).
Representative DNA sequence: A nucleotide sequence representing a fragment (905 bp long; OR773087) including the partial ITS1 region (370 bp long), 5.8S rDNA (157 bp) and the partial ITS2 region (378 bp). No intraspecific variability was found.
Infection indices: prevalence at type host 30%, 8–41 monogeneans per infected host.
Etymology: The specific name ‘iraqemembranatus’ refers to the morphological similarity between the new species found on cyprinid hosts from Iraq and its previously described congener G. emembranatus Malmberg, Reference Malmberg1970, whose specific name refers to the absence of the membrane in the ventral bar.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:B4738C07-9748-4217-80C0-D5510AC31E4F.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner root uncovered; base with no posterior folds; root short; shaft slightly bowed; point curved and elongated. Ventral bar lacking bilateral processes and membrane; median part of a common width. Dorsal bar constricted at halfway point, with attenuated ends inserted into terminal plates. Marginal hooks with a flat base, circular heel downward, blunt toe slightly outward, no shelf, curved point, sickle proper approximately perpendicular to the base, gently curved downward to a point approximately the same level of toe; filament loop extending almost handle length. MCO with single prominent apical spine and row of at least 10 spinelets.
Differential diagnosis: Previous parasitological investigations performed by Al-Sa'adi (Reference Al-Sa'adi2007) on A. sellal, a native leuciscid inhabiting watersheds in Iraq, revealed the presence of specimens that were assigned to G. sprostonae Ling, 1962 (Mhaisen and Abdul-Ameer, Reference Mhaisen and Abdul-Ameer2013). The original work of Al-Sa'adi (Reference Al-Sa'adi2007) was unavailable for us to check the validity of this assignment. Considering the overall morphology of G. sprostonae, known so far from a range of widespread cyprinids (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009), the newly described G. iraqemembranatus n. sp. from A. sellal differs considerably from the former species in having (i) shorter hamuli (23.1–25.0 μm in G. iraqemembranatus n. sp. vs 41.0–62.0 μm in G. sprostonae) and (ii) a shorter ventral bar (2.7–8.2 μm in G. iraqemembranatus n. sp. vs 13.0–26.0 μm in G. sprostonae) with no membrane (vs well-developed membrane in G. sprostonae). The distinction between G. iraqemembranatus n. sp. and G. sprostonae was further supported by genetic data (Fig. 7). Likewise, specimens identified as G. sprostonae were also reported on B. lacerta occurring in the Tajan River, together with G. ctenopharyngodonis Ling, 1962 (both localities in Iran), but no drawings of the haptoral structures were included (Barzegar et al., Reference Barzegar, Ebrahimzadeh Mousavi, Rahmati-Holasoo, Taheri Mirghaed and Bozorgnia2018). With regard to meristic data available in Barzegar et al. (Reference Barzegar, Ebrahimzadeh Mousavi, Rahmati-Holasoo, Taheri Mirghaed and Bozorgnia2018), G. iraqemembranatus n. sp. is highly distinguishable from G. ctenopharyngodonis in having (i) shorter hamuli (23.1–25.0 μm in G. iraqemembranatus n. sp. vs 53.8–54.5 μm in G. ctenopharyngodonis), (ii) a shorter ventral bar (2.7–8.2 μm in G. iraqemembranatus n. sp. vs 20.7–21.2 μm in G. ctenopharyngodonis) with no membrane, (iii) a shorter dorsal bar (6.8–8.5 μm in G. iraqemembranatus n. sp. vs 15.4–16.1 μm in G. ctenopharyngodonis) and (iv) shorter marginal hooks (11.0–12.9 μm in G. iraqemembranatus n. sp. vs 24.6–25.2 μm in G. ctenopharyngodonis). Hitherto, G. elegans (Nordmann, 1832) was reported by Nasraddin (Reference Nasraddin2013) on P. trutta inhabiting the Middle East (Iraq) (Mhaisen and Abdul-Ameer, Reference Mhaisen and Abdul-Ameer2013), and eastern Anatolia (Turkey) (Koyun et al., Reference Koyun, Korkut and Gül2019). Since no morphology of the haptoral apparatus of G. elegans from P. trutta has so far been detailed, it remains hard to know whether the previously collected specimens truly corresponded to G. elegans. This latter species has been repeatedly misidentified and many researchers have randomly assigned dozens of species to G. elegans (see remarks in Malmberg, Reference Malmberg1970; Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). Gyrodactylus iraqemembranatus n. sp. resembles G. elegans regarding the ventral bar, which lacks bilateral processes. Besides the size of the haptoral sclerotized structures, the main differences between G. iraqemembranatus n. sp. and G. elegans are in (i) the shape of the hamuli, which have poorly developed roots in G. iraqemembranatus n. sp. (vs well-developed roots in G. elegans) and in (ii) the ventral bar membrane, which is absent in G. iraqemembranatus n. sp. (vs the presence of a spine-like shaped membrane in G. elegans) (Malmberg, Reference Malmberg1970). The distinction between G. iraqemembranatus n. sp. and G. elegans was further supported by genetic data (Fig. 7). Gyrodactylus markevitschi Kulakovskaya, 1951 was first described from European Barbus barbus (Linnaeus, 1758), then later reported from distinct west Asian locations (Iraq) (Mhaisen and Abdul-Ameer, Reference Mhaisen and Abdul-Ameer2013; Koyun et al., Reference Koyun, Korkut and Gül2019). It was also recorded on a range of Palearctic cyprinids (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). On the basis of merisitic data available in Pugachev et al. (Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009), G. iraqemembranatus n. sp. differs from G. markevitschi in having (i) shorter hamuli (23.1–25.0 μm in G. iraqemembranatus n. sp. vs 56–58 μm in G. markevitschi), (ii) a shorter ventral bar (2.7–8.2 μm in G. iraqemembranatus n. sp. vs 22.0–25.0 μm in G. markevitschi) with no bilateral processes, (iii) a shorter dorsal bar (6.8–8.5 μm in G. iraqemembranatus n. sp. vs 18.0–20.0 μm in G. markevitschi) and finally (iv) shorter marginal hooks (11.0–12.9 μm in G. iraqemembranatus n. sp. vs 26.0–29.0 μm in G. markevitschi).
In this study, G. iraqemembranatus n. sp. showed intraspecific variability in the size of the ventral bar, where the longest and narrowest ventral bar was observed in specimens parasitizing A. sellal (see Table 3).
Gyrodactylus jurajdai Rahmouni n. sp. (Fig. 11)
Type-host: Chondrostoma regium (Heckel, 1843) (Cyprinoidei: Leuciscidae)
Hard structures of haptor of Gyrodactylus jurajdai n. sp. ex Chondrostoma regium.

Type-locality: Du Choman, the Aw-e Shiller River, Sulaymaniyah Province, Iraq
Site on host: Fins
Type material: Holotype and 1 paratype (IPCAS M-785).
Representative DNA sequence: A nucleotide sequence representing a fragment (1117 bp long; OR773088) including the partial ITS1 region (546 bp long), 5.8S rDNA (157 bp) and the partial ITS2 region (414 bp). No intraspecific variability was found.
Infection indices: Prevalence 50%, 1–2 monogeneans per infected host.
Etymology: The specific name ‘jurajdai’ honours the ichthyologist Dr Pavel Jurajda from the Institute of Vertebrate Biology, Czech Academy of Sciences (Brno, Czech Republic) in recognition of his crucial contribution to parasitological work.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:CA4D2FD9-E78F-4BCF-BE22-5CAC37EE457D.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner root with narrow wart-like projections anteriorly; base with posterior folds; relatively long; shaft slightly bowed; point curved and elongated. Ventral bar with blunt, short, triangular bilateral processes extending out of bar; median part may show a hollow; membrane oval, almost 2/3 length of hamuli shaft, with striations ending posteriorly in a median ridge. Dorsal bar curved, slightly swollen at halfway point, with attenuated ends inserted into terminal plates. Marginal hooks with flat globose heel, elongate toe and curved downward, conspicuous shelf, curved point, sickle proper gently curved downward to a point approximately perpendicular to toe shelf; filament loop extending almost 2/3 handle length. MCO not observed.
Differential diagnosis: We present herein the first morphological characterization of a gyrodactylid species from C. regium. Gyrodactylus jurajdai n. sp. resembles its Eurasian relatives known from Chondrostoma spp. regarding the haptoral sclerotized structures – specifically, G. chondrostomi Ergens, 1967 and G. macrocornis Ergens, 1963, both parasitizing C. nasus (Linnaeus, 1758), and G. derjavini Mikailov, 1975 from Chondrostoma oxyrhynchum Kessler, 1877 (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). Gyrodactylus jurajdai n. sp. differs from G. chondrostomi in having (i) longer hamuli (57.2–74.5 μm in G. jurajdai n. sp. vs 38.0–40.0 μm in G. chondrostomi), (ii) shorter ventral bar (22.0–33.4 μm in G. jurajdai n. sp. vs 18.0–19.0 μm in G. chondrostomi) and (iii) longer dorsal bar (24.1–31.3 μm in G. jurajdai n. sp. vs 15.0–17.0 μm in G. chondrostomi), and (iv) differently shaped sickle of marginal hooks with a conspicuous shelf in G. jurajdai n. sp. in comparison to that in G. chondrostomi. The new species is discriminated from G. macrocornis by having (i) a ventral bar possessing a ridge (ridge missing in G. macrocornis) and (ii) relatively shorter hamuli (57.2–74.5 μm in G. jurajdau n. sp. vs 74.0–58.0 μm in G. macrocornis).
Gyrodactylus mhaiseni Rahmouni n. sp. (Fig. 12)
Type-host: Alburnus sellal Heckel, 1843 (Cyprnoidei: Leuciscidae)
Hard structures of haptor of Gyrodactylus mhaiseni n. sp. ex Alburnus sellal.

Type-locality: wadi Kalat Shirah, a tributary of the Tabin River, Sulaymaniyah Province, Iraq
Additional locality: Zahrzi in Tabin River, Sulaymaniyah Province, Iraq
Type material: Holotype and 2 paratypes (IPCAS M-786).
Site on host: Fins
Representative DNA sequence: A nucleotide sequence representing a fragment (1148 bp long; OR773082) including the partial ITS1 region (551 bp long), 5.8S rDNA (157 bp) and the partial ITS2 region (440 bp). Minor intraspecific variability was found between 2 host populations (ITS1, p-distance = 0.4%; ITS2, p-distance = 0.3%).
Infection indices: Prevalence at the type locality 25%, 1–4 monogeneans per infected host.
Etymology: The specific name ‘mhaiseni’ honours the parasitologist Professor Furhan T. Mhaisen in recognition of his crucial contribution to parasitological work on marine and freshwater fishes in the Middle East.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:31283AC6-1CD4-450B-998F-A1D5042C97E5.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner root with narrow wart-like projections anteriorly; base with posterior folds; root long; shaft slightly bowed; point curved and elongated. Ventral bar with blunt, short, almost triangular bilateral processes extending out of bar; median part with a hollow; membrane slightly trapezoid, almost 1/2 length of hamuli shaft, with striations ending posteriorly in a median ridge. Dorsal bar straight, with projections near extremities and attenuated ends inserted into terminal plates. Marginal hooks with globose downward heel, elongate toe and curved downward, conspicuous shelf, curved point, sickle proper approximately perpendicular to base, gently curved downward to a point slightly perpendicular to toe shelf; filament loop extending almost 1/2 handle length. MCO not observed.
Differential diagnosis: In addition to G. iraqemembranatus n. sp., A. sellal hosted another species recognized as new to science, namely G. mhaiseni n. sp. These 2 species are easily distinguishable regarding the morphotype of the hamuli, comprising long roots in G. mhaiseni n. sp. unlike the poorly developed ones in G. iraqemembranatus n. sp., as well as that of the ventral bar, showing bilateral processes and a long membrane with a ridge in the former species, but the absence of these features in G. iraqemembranatus n. sp. According to genetic data, slight intraspecific variability in haptoral sclerites was observed at the geographical scale. With respect to G. sprostonae, a species already reported from A. sellal (Iraq) (Mhaisen and Abdul-Ameer, Reference Mhaisen and Abdul-Ameer2013), measurements of the haptoral sclerites overlap those of G. mhaiseni n. sp. The main differences between these 2 species are (i) the projections on the dorsal bar, (ii) the median ridge in the ventral bar membrane and (iii) the thick shaft of the hook sickle in G. mhaiseni n. sp., all features missing in G. sprostonae (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009).
Gyrodactylus sandai Rahmouni n. sp. (Fig. 13)
Type-host: Capoeta umbla (Heckel, 1843) (Cyprinoidei: Cyprinidae)
Hard structures of haptor of Gyrodactylus sandai n. sp. ex Capoeta umbla (A), ex Cyprinion macrostomum (B).

Type-locality: wadi Kalat Shirah, tributary of the Tabin River, Sulaymaniyah Province, Iraq
Additional hosts: Cyprinion macrostomum Heckel, 1843 (Cyprinoidei: Cyprinidae)
Site on host: Fins
Type material: Holotype and 2 paratypes (IPCAS M-787/1-2).
Representative DNA sequence: A nucleotide sequence representing a fragment (1120 bp long; OR773089) including the partial ITS1 region (491 bp long), 5.8S rDNA (157 bp) and the partial ITS2 region (472 bp). No intraspecific variability was found.
Infection indices: Prevalence at type host 50%, 1–2 monogeneans per infected host.
Etymology: The specific name ‘sandai’ honours the ichthyologist Dr Radek Šanda from the Czech National Museum (Prague, Czech Republic) in recognition of his crucial contribution to parasitological work and his precious help in identifying cyprinoid fish hosts during the field trips.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:F7B361CD-A677-4135-86CF-0F9A948F9486.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner roots with a hat-like cover; base may show groove-like folds; root long; shaft slightly bowed; point curved and elongated. Ventral bar with blunt, well-developed bilateral processes extending out of bar; median part may show a hollow; membrane elongated, oval, almost 2/3 length of hamuli shaft, with striations ending posteriorly in a median ridge. Dorsal bar straight, may show bifurcated projections near extremities, with attenuated ends inserted into terminal plates. Marginal hooks with globose heel, finger-like toe curved downward, conspicuous shelf, curved point, sickle proper approximately perpendicular to base, gently curved downward to a point slightly exceeding the toe; filament loop extending almost 1/3 handle length. MCO not observed.
Differential diagnosis: While no parasitological data were available for C. umbla, G. baicalensis Bogolepova, 1950 and G. elegans were previously reported to parasitize C. macrostomum from the Middle East (Mhaisen and Abdul-Ameer, Reference Mhaisen and Abdul-Ameer2013; Mhaisen et al., Reference Mhaisen, Abdul-Ameer and Hamdan2018). Gyrodactylus baicalensis was originally described from euryhaline hosts, namely Limnocottus godlewskii (Dybowski, 1874) and Batrachocottus multiradiatus Berg, 1907 (both Perciformes, Cottidae) and then reported on Planiliza abu (Heckel, 1843) (Iran) (Mugiliformes, Mugilidae) (Kritsky et al., Reference Kritsky, Ali and Khamees2013; Al-Jawda and Ali, Reference Al-Jawda and Ali2020). Gyrodactylus sandai n. sp. is easily distinguishable from both G. elegans and G. baicalensis by the well-developed bilateral processes on its ventral bar, these features either small or missing in G. elegans and G. baicalensis. Intraspecific variability was observed, where specimens parasitizing C. macrostomum exhibited a slightly longer dorsal bar compared to those parasitizing C. umbla. Gyrodactylus sandai n. sp., isolated herein from Middle Eastern C. umbla, is reminiscent of its Palearctic congener G. katharineri Malmberg, 1964 known from a wide range of Palearctic cyprinoids (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). Morphological similarities are mainly in (i) the shape of the hamuli base with covered tips and folds, (ii) the bifurcated projections on the dorsal bar and (ii) the posterior median ridge present in the ventral bar membrane of both species. Gyrodactylus katharineri is widely distributed in the Palearctic region, which has resulted in significant morphological intraspecific variability (highly variable sizes of the haptoral sclerotized structures; see meristic data in Pugachev et al. [Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009]). The newly described G. sandai n. sp. and G. katharineri had been taxonomically separated regarding the endemism of C. umbla to Tigris and Euphrates freshwaters (Froese and Pauly, Reference Froese and Pauly2023), this supported significant genetic divergence revealed by mean of ITS sequences (ITS1 [495 bp long], p-distance = 15.4%; 5.8S-ITS2 (614 bp long), p-distance = 2.9% [Matějusová et al., Reference Matějusová, Gelnar, McBeath, Collins and Cunningham2001a and references herein]). Gyrodactylus sandai n. sp. and the newly described G. mhaiseni n. sp. exhibit a similarly shaped ventral bar with a posterior median ridge. The main difference between these 2 species is the larger size of each haptoral sclerotized structure exhibited by G. sandai n. sp. compared to G. mhaiseni n. sp. Moreover, G. sandai n. sp. and G. jurajdai n. sp. both possess hamuli with conspicuous folds and a ventral bar membrane garnished with a median ridge. Compared to G. mhaiseni n. sp., G. sandai n. sp. showed (i) longer hamuli (100.0–105.1 μm in G. sandai n. sp. vs 51.7–59.9 μm in G. mhaiseni n. sp.), (ii) longer ventral bar (58.5–63.9 μm in G. sandai n. sp. vs 19.8–25.6 μm in G. mhaiseni n. sp.), with (iii) longer bilateral processes (16.6–22.6 μm in G. sandai n. sp. vs 1.6–2.5 μm in G. mhaiseni n. sp.), (iv) longer dorsal bar (47.0–55.0 μm in G. sandai n. sp. vs 19.9–25 μm in G. mhaiseni n. sp.) and finally (v) longer marginal hooks (45.0–46.5 μm in G. sandai n. sp. vs 23.2–27.4 μm in G. mhaiseni n. sp.), with (vi) longer handle (37.4–39–1 μm in G. sandai n. sp. vs 17.7–21.6 μm in G. mhaiseni n. sp.).
Gyrodactylus satanicus Rahmouni n. sp. (Fig. 14)
Type-host: Garra rufa (Heckel, 1843) (Cyprinoidei: Cyprinidae)
Hard structures of haptor of Gyrodactylus satanicus n. sp. ex Garra rufa.

Type-locality: By the road Sulaymaniyah–Dukan, Little Zab, Iraq
Site on host: Fins
Type material: Holotype and 2 paratypes (IPCAS M-788).
Representative DNA sequence: A nucleotide sequence representing a fragment (1275 bp long; OR773091) including the partial ITS1 region (683 bp long), 5.8S rDNA (157 bp) and the partial ITS2 region (435 bp). No intraspecific variability was found.
Infection indices: Prevalence 70%, 1–5 monogeneans per infected host.
Etymology: The specific name ‘satanicus’ (as an adjective in the nominative singular) refers to the morphology of the hamuli, with horn-like projections reminiscent of a silhouette of the devil's face.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:AB3D872E-3A8F-4443-8C33-DBED0892DFE0.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner roots uncovered; base with 2 pairs of horn-like anterior projections and may show groove-like posterior folds; root relatively long, slightly curved inwards; shaft slightly bowed; point curved and elongated. Ventral bar with blunt, short, triangular bilateral processes extending out of bar; median part with hollows; membrane elongated, oval, almost 2/3 length of hamuli shaft ending posteriorly in a median ridge. Dorsal bar straight, slightly constricted at halfway point, with attenuated ends inserted into terminal plates. Marginal hooks with globose heel, blunt toe, conspicuous shelf, curved point, sickle proper gently curved downward to a point perpendicular to toe shelf; filament loop extending almost 1/2 handle length. MCO not observed.
Differential diagnosis: Gyrodactylus elegans is the sole species hitherto reported from G. rufa in Iraq (Mhaisen and Abdul-Ameer, Reference Mhaisen and Abdul-Ameer2013). The typical morphology of the hamuli of G. satanicus n. sp. with 2 pairs of horn-like projections makes it easily distinguishable from G. elegans and all congeners known so far. The median ridge in the ventral bar membrane exhibited by G. satanicus n. sp. is a common feature in Palearctic Gyrodactylus, as already discussed above (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). This makes the present study the very first one to report a Gyrodactylus species with such features in the Middle East region.
Gyrodactylus vukicae Rahmouni n. sp. (Fig. 15)
Type-host: Garra rufa (Heckel, 1843) (Cyprinoidei: Cyprinidae)
Hard structures of haptor of Gyrodactylus vukicae n. sp. ex Garra rufa.

Type-locality: By the road Sulaymaniyah–Dukan, Little Zab, Iraq
Site on host: Fins
Type material: Holotype and 1 paratype (IPCAS M-789).
Representative DNA sequence: A nucleotide sequence representing a fragment (1218 bp long; OR773090) including the partial ITS1 region (623 bp), 5.8S rDNA (157 bp) and the ITS2 region (438 bp). Minor intraspecific genetic variability was found in the ITS1 (P-distance = 0.4%).
Infection indices: Prevalence 20%, 1 monogenean per infected host.
Etymology: The specific name ‘vukicae’ honours the ichthyologist Dr Jasna Vukić from the Faculty of Sciences, Charles University (Prague, Czech Republic) in recognition of her crucial contribution to parasitological work and her precious help in identifying the cyprinoid hosts during the field trips.
ZooBank registration (LSID): urn:lsid:zoobank.org:act:3D2F7DB6-34A1-4534-B7A7-62AD66D14A0C.
Description: (Dimensions of the hard structures are given in Table 3.) Haptor subcircular; tips of inner roots with a hat-like cover; base with posterior folds; root long; shaft slightly bowed; point curved and elongated. Ventral bar with blunt, short bilateral processes extending out of bar; median part with no visible hollows; membrane slightly trapezoid, almost 1/3 length of hamuli shaft, may show a posterior ridge. Dorsal bar gently curved, with attenuated ends inserted into terminal plates. Marginal hooks with globose downward heel, triangular, curved finger-like toe curved downward, conspicuous shelf, curved point, sickle proper gently curved downward to a point slightly exceeding toe shelf; filament loop (lamella) extending over 1/2 handle length, MCO not observed.
Differential diagnosis: In association with G. satanicus n. sp. described above, G. rufa was shown to host an additional species described herein as G. vukicae n. sp. (Fig. 13). These 2 species are largely discriminated from each other by the atypical pairs of horn-like projections on the hamuli exhibited by G. satanicus n. sp. (see above). Gyrodactylus vukicae n. sp. is further easily differentiated from G. elegans, the only species previously reported from G. rufa (Mhaisen and Abdul-Ameer, Reference Mhaisen and Abdul-Ameer2013), by the shape of the ventral bar parts, mainly the blunt short bilateral processes and trapezoid ventral bar membrane in G. vukicae n. sp., which are in contrast to the form of the ventral bar in G. elegans, which lacks processes and exhibits a long and narrow membrane (Malmberg, Reference Malmberg1970; Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009).
Discussion
Although the monogenean fauna of freshwater fishes has been extensively studied in Iraq (e.g. Asmar et al., Reference Asmar, Balasem, Mhaisen, Al-Khateeb and Al-Jawda1999; Al-Awadi, Reference Al-Awadi2003; Balasem et al., Reference Balasem, Mhaisen, Adday, Al-Jawda and Asmar2003; Mhaisen et al., Reference Mhaisen, Balasem, Al-Khateeb and Asmar2003, Reference Mhaisen, Al-Rubae and Al-Sa'adi2015; Abdullah and Mhaisen, Reference Abdullah and Mhaisen2005; and numerous local student theses), the present study indicates that only a fraction of its real regional species diversity has been previously uncovered. Nowadays, an integrative approach combining morphological and molecular data is commonly applied in studies on monogeneans. It appears that the diversity of freshwater fish monogeneans revealed by this approach is much higher (Kmentová et al., Reference Kmentová, Gelnar, Mendlová, Van Steenberge, Koblmüller and Vanhove2016; Dos Santos and Avenant-Oldewage, Reference Dos Santos and Avenant-Oldewage2020; Řehulková et al., Reference Řehulková, Benovics and Šimková2020, Reference Řehulková, Rahmouni, Pariselle and Šimková2021; Šimková et al., Reference Šimková, Řehulková, Choudhury and Seifertová2022) than previously expected on the basis of morphology alone. In the current study, we recorded a total of 33 monogenean species on endemic cyprinoids in Iraq. Comparing the ectoparasitic monogeneans to endoparasitic helminth taxa parasitizing cyprinoids in Iraq, the former appear to be much more diversified (Mhaisen and Abdullah, Reference Mhaisen and Abdullah2017; Öktener, Reference Öktener and Jawad2021), similarly as was recorded for cyprinoids in Europe (e.g. Loot et al., Reference Loot, Reyjol, Poulet, Šimková, Blanchet and Lek2007; Seifertová et al., Reference Seifertová, Vyskočilová, Morand and Šimková2008; Rohlenová et al., Reference Rohlenová, Morand, Hyršl, Tolarová, Flajšhans and Šimková2011; Krasnovyd et al., Reference Krasnovyd, Vetešník, Gettová, Civáňová and Šimková2017; Pakosta et al., Reference Pakosta, Vetešník and Šimková2018; Benovics et al., Reference Benovics, Francová, Volta, Dlapka and Šimková2021b; see also review of Kuchta et al., Reference Kuchta, Řehulková, Francová, Scholz, Morand and Šimková2020). This is most likely connected to the overall level of host specificity, which is significantly higher in monogeneans than in, for example, cestodes or digeneans (Rohde, Reference Rohde1979; Whittington et al., Reference Whittington, Bronwen, Hamwood and Halliday2000; Cribb et al., Reference Cribb, Chisholm and Bray2002; Tkach et al., Reference Tkach, Littlewood, Olson, Kinsella and Swiderski2003; Kuchta et al., Reference Kuchta, Řehulková, Francová, Scholz, Morand and Šimková2020). In the present study, we recorded 27 monogenean species only from a single host species, although not all of them can be considered as strictly host-specific, as they were previously recorded also on other cyprinoid hosts in the region (e.g. D. goktschaicus Gussev, 1966 is a common parasite of Barbus spp. and Luciobarbus spp. in the Middle East [Pazooki et al., Reference Pazooki, Jalali Jafari and Ghobadian2006; Koyun et al., Reference Koyun, Ulupinar and Gül2015; Koyuncu et al., Reference Koyuncu, Yilmaz, Secer, Kaya, Genc and Jawad2021; Benovics et al., Reference Benovics, Nejat, Abdoli and Šimková2021a], and D. vistulae is the Dactylogyrus species with the widest distribution range in the western Palearctic [Seifertová et al., Reference Seifertová, Vyskočilová, Morand and Šimková2008; Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018, Reference Benovics, Vukić, Šanda, Nejat, Charmpila, Buj, Shumka, Porcelloti, Tarkan, Aksu, Emiroğlu and Šimková2023]). One out of these recorded monogenean species are the first record for Iraq, and additional 17 are newly discovered species (4 Dactylogyrus, 12 Gyrodactylus and 1 Dogielius). From the previously described species, only D. holciki, found in our study on A. sellal and non-described Alburnus sp. from the Tabin River and Zalm Stream (near Grdi Go), respectively, was a new host record for Iraq. Although D. holciki is a common parasite of A. mossulensis Heckel, 1843 and A. chalcoides Güldenstädt, 1772 in the Middle East (e.g. Al-Samman et al., Reference Al-Samman, Molnár and Székely2006; Aydoğdu et al., Reference Aydoğdu, Selver and Çirak2008; Tunç and Koyun, Reference Tunç and Koyun2018), described in Iran by Molnár and Jalali (Reference Molnár and Jalali1992), it was not previously reported in Iraq. Considering its host range, this species might be recognized as genus specific for Alburnus, and we can assume that its distribution will be restricted to that of the fish of this genus in the Middle East.
Following the host specificity levels designed in Šimková et al. (Reference Šimková, Verneau, Gelnar and Morand2006) and modified in Benovics et al. (Reference Benovics, Nejat, Abdoli and Šimková2021a), and including also previous host records for the respective species, all collected monogenean species fell into 4 categories – true generalists (D. vistulae, D. mokhayeri, G. iraqemembranatus n. sp. and P. homoion) parasitizing host species from different cyprinoid families; transitional generalists (D. goktschaicus, D. deziensis Gussev, Jalali and Molnár, Reference Gussev, Jalali and Molnár1993, D. deziensioides Gussev, Jalali and Molnár, Reference Gussev, Jalali and Molnár1993, D. molnari, D. cf. persicus and G. sandai n. sp.) parasitizing hosts belonging to a single subfamily; intermediate specialists (D. holciki and D. lenkorani); and strict specialists parasitizing only a single host species (all the remaining collected monogenean species). It appears that the cyprinoids in Iraq are predominantly parasitized by highly host-specific monogenean taxa. These also include taxa which are endemic to the Middle Eastern region (only D. vistulae was also previously documented outside the Middle East, e.g. Šimková et al., Reference Šimková, Morand, Jobet, Gelnar and Verneau2004; Seifertová et al., Reference Seifertová, Vyskočilová, Morand and Šimková2008; Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018), thus we can assume that they cospeciated with their respective cyprinoid host species over long evolutionary time. While Dactylogyrus, Dogielius and Paradiplozoon are almost exclusively limited to cyprinoid fishes (Gibson et al., Reference Gibson, Timofeeva and Gerasev1996; Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009), Gyrodactylus is the only reported genus with a host range encompassing also other fish taxa (e.g. gasterosteoids, salmonids, silurids, see Bakke et al., Reference Bakke, Harris, Jansen and Hansen1992, Reference Bakke, Cable and Harris2007; Harris et al., Reference Harris, Shinn, Cable and Bakke2004). The differences in host specificity levels are most likely connected to life cycle, as gyrodactylids are strictly viviparous; thus, their dispersal into new hosts is limited to direct contact between fish (Bakke et al., Reference Bakke, Cable and Harris2007) rendering the parasites opportunistic host-switchers. Surprisingly, only G. iraqemembranatus n. sp. and G. sandai n. sp. were collected from more than 1 species within our study; nevertheless, 2 host species for each of these Gyrodactylus spp. were collected from the same site (Kani Shok and wadi Kalat Shirah, respectively). These observations and recorded host–parasite associations only support the low host preferences of Gyrodactylus parasites. On the other hand, Dactylogyrus, Dogielius and Paradiplozoon are oviparous monogenean taxa which are known to have developed various morphological (Sasal et al., Reference Sasal, Trouvé, Müller-Graf and Morand1999; Huyse and Volckaert, Reference Huyse and Volckaert2002; Šimková et al., Reference Šimková, Verneau, Gelnar and Morand2006) and ecological (Whittington et al., Reference Whittington, Chisholm and Rohde1999; Buchmann and Lindenstrøm, Reference Buchmann and Lindenstrøm2002; Whittington and Kearn, Reference Whittington and Kearn2011) adaptations to seek and infest associated hosts and the ones most suitable for retaining parasite populations. These adaptations putatively limit the host-switching capacities of the given monogenean species and therefore their dispersal capacity is restricted to the dispersal of their respective hosts. The excepted high host specificity among dactylogrid monogeneans was in accordance with our observations, as the majority of species within the genera Dactylogyrus and Dogielius are restricted either to a single host species, or several local congeners. In Europe and North America, 30 and 61% of Dactylogyrus species, respectively, appear to be strictly host specific (Kuchta et al., Reference Kuchta, Řehulková, Francová, Scholz, Morand and Šimková2020). In our local study, focusing only on monogenean diversity in Iraq, 15 out of 16 Dactylogyrus species were recorded only from a single host species (although several species exhibit wider host range, as is discussed above). A similar high level of host specificity could initially be assumed also for Dogielius species, as D. mokhayeri was initially reported only from Leuciscus vorax (Heckel, 1843), an endemic species in the Middle East (Jalali and Molnár, Reference Jalali and Molnár1990; Abdullah and Mhaisen, Reference Abdullah and Mhaisen2005); however, our records suggest that the host range for this monogenean species might also encompass other leuciscid species in the Middle East, and thus this species is endemic to the Middle East without any host preferences. The other 2 previously described Dogielius species (i.e. D. cf. persicus and D. molnari Jalali, 1992) were reported in Iraq for the first time by Abdullah and Mhaisen (Reference Abdullah and Mhaisen2005), who recorded all 3 Dogielius species from 3 cyprinoid species in the Grater Zab River. In addition, it appears that in the Tabin River basin a potentially new Dogielius species can also be found, as the previously undescribed species was collected from C. umbla in Wadi Kalat Shirah valley. This putatively new Dogielius species will be properly described after the collection of additional material for morphometric data, and the description will be included in a study also investigating congeners from other geographical regions.
The phylogenetic analyses performed on the species of 2 selected monogenean genera (Dactylogyrus and Gyrodactylus) revealed that the endemic congeneric species from Iraq did not form a monophyletic group. The majority of Dactylogyrus species studied herein were positioned within the phylogenetic lineage A, encompassing Middle Eastern, European, North African and North American species. Several well-supported groups were formed within lineage A; however, the molecular data used in this study were not sufficiently phylogenetically informative to fully resolve the relationships between them. Within the groups, clustered species shared the morphotypes of taxonomically important characters, mainly the hard parts of the attachment organ. Dactylogyrus species associated with cyprinid hosts (i.e. Barbus, Capoeta, Carasobarbus, Cyprinion, Garra, Luciobarbus and Paracapoeta) were in paraphyly and were included in the 5 phylogenetic groups. While D. persis and D. barbuli clustered together with north African, Middle Eastern and European (Iberian in this case) congeners possessing the ‘carpathicus’ morphotype of haptoral connective ventral bar (see Pugachev et al. [Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009] for classification of morphotypes), D. goktschaicus, D. deziensis and D. deziensioides formed a group with European and Middle Eastern congeners. The latter group possesses the ventral bar of various shapes (especially the ‘tissensis’ and ‘rutili’ type); however, it is considered as ‘less complex’, i.e. with a lower number of extremities, when compared to the most complex ventral bar (Benovics et al., Reference Benovics, Nejat, Abdoli and Šimková2021a). The grouping of Dactylogyrus spp. of cyprinids within lineage A is in accordance with the proposed dual historical divergence and dispersion patterns for cyprinids (Doadrio, Reference Doadrio1990; Casal-Lopéz and Doadrio, Reference Casal-Lopéz and Doadrio2018) and their associated parasites (Benovics et al., Reference Benovics, Nejat, Abdoli and Šimková2021a). By the distribution of the respective Dactylogyrus species from Iraqi cyprinoids within 2 morphogroups, we can hypothesize that they historically split within the Middle Eastern region and while the group encompassing D. barbuli and phylogenetically close congeners sharing the ‘carpathicus’ morphotype of ventral bar dispersed via North Africa, the other species dispersed directly into Europe via the Balkan-Anatolian connection (Gomphoterium Land Bridge, Steininger and Rögl [Reference Steininger, Rögl, Dixon and Robertson1984]; Perea et al. [Reference Perea, Böhme, Zupančič, Freyhof, Šanda, Özuluğ, Abdoli and Doadrio2010]). Unprecedented (see Šimková et al. [Reference Šimková, Morand, Jobet, Gelnar and Verneau2004]; Benovics et al. [Reference Benovics, Nejat, Abdoli and Šimková2021a] or Acosta et al. [Reference Acosta, Truter, Malherbe and Smit2022] for comparison) is the phylogenetic position of the 2 presumably host-specific Dactylogyrus species, D. microcirrus and D. macrosotomi (from P. trutta and C. macrostomum, respectively), which formed an individual phylogenetic lineage. The deep nodal split of this lineage from other Dactylogyrus lineages and the inclusion of the Iraqi cyprinoid-specific Dactylogyrus species within suggest early diversification in the region, and we can assume that this lineage is endemic to the Middle Eastern area. We can hypothesize that this lineage may include also other endemic congeners sharing similar morphological features (e.g. D. barbioides Gussev et al., Reference Gussev, Jalali and Molnár1993, described from Arabibarbus grypus [Heckel, 1843] or D. cyprinioni parasitizing C. macrostomum, see Pugachev et al. [Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009]); however, the molecular data for these species are still missing. The ancestral origin of this lineage in the Middle East can be linked to the phylogenetic relationships of their hosts, as Paracapoeta is a sister to endemic Capoeta, with the basal position (Turan et al., Reference Turan, Kaya, Aksu and Bektaş2022), and Cyprinion represents a divergent phylogenetic entity in the region (Durand et al., Reference Durand, Tsigenopoulos, Ünlü and Berrebi2002; Yang et al., Reference Yang, Sado, Hirt, Pasco-Viel, Arunachalam, Li, Wang, Freyhof, Saitoh, Simons, Miya, He and Mayden2015) (although, the phylogenetic relationships among the Cyprinion species are not yet well investigated). Dactylogyrus cyprinioni is presumably specific to Cyprinion spp. which further supports our assumption of the position of this Dactylogyrus species within the endemic lineage; however, further investigation is required.
Dactylogyrus regius n. sp. formed a well-supported group with congeners possessing the inverted T-shape morphotype of ventral bar, and the ‘chondrostomi’ morphotype of MCO, which are common for Dactylogyrus species parasitizing Chondrostoma s. l. hosts, such as D. ergensi, D. elegantis and D. globulatus (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009; Řehulková et al., Reference Řehulková, Benovics and Šimková2020; Benovics et al., Reference Benovics, Desdevises, Šanda, Vukić and Šimková2020a, Reference Benovics, Francová, Volta, Dlapka and Šimková2021b). Therefore, we can assume that the species of this group are historically associated with Chondrostoma hosts, and only secondarily host-switched and speciated on other leuciscids (e.g. D. tissensis is a common species of Alburnoides spp. [Benovics et al., Reference Benovics, Desdevises, Vukić, Šanda and Šimková2018], and D. sagittarius parasitizes Telestes [Benovics et al., Reference Benovics, Francová, Volta, Dlapka and Šimková2021b]; however, all these species share the same morphological features). Contrastingly, Dactylogyrus anoigeus n. sp. described from A. marmid was not phylogenetically close to congeners associated with Abramis (e.g. D. auriculatus [von Nordmann, 1832] and D. zandti Bychowsky, 1933 [Dzika, Reference Dzika2002; Krasnovyd et al., Reference Krasnovyd, Vetešník and Šimková2020; Dedić et al., Reference Dedić, Vetešník and Šimková2023]) or Blicca (e.g. D. cornu Linstow, 1878 [Soylu, Reference Soylu2012; Krasnovyd et al., Reference Krasnovyd, Vetešník and Šimková2020], despite the phylogenetic proximity of these host genera [Teimori et al., Reference Teimori, Esmaeili, Sayyadzadeh, Zarei and Gholamhosseini2015; Schönhuth et al., Reference Schönhuth, Vukić, Šanda, Yang and Mayden2018]). Dactylogyrus anoigeus n. sp. grouped with congeners parasitizing Squalius spp., such as D. folkmanovae, D. vranoviensis (Seifertová et al., Reference Seifertová, Vyskočilová, Morand and Šimková2008; Benovics et al., Reference Benovics, Vukić, Šanda, Nejat, Charmpila, Buj, Shumka, Porcelloti, Tarkan, Aksu, Emiroğlu and Šimková2023) and the herein described D. rivalis n.sp., which appears to be a species endemic to the Middle East.
Novel molecular data for 2 Dactylogyrus species parasitizing G. rufa (i.e. D. acinacus and D. medicus n. sp.) revealed the phylogenetic proximity of these species to congeners with a putative origin in Eastern Asia, and associated with the cyprinids Carassius and Cyprinus (e.g. D. anchoratus [Dujardin, 1845], D. formosus Kulwiec, 1927 and D. vastator [Nybelin, 1924]), Barbonymus (D. tapienensis and D. viticulus), Middle Eastern Capoeta (i.e. D. pulcher) and North African Carasobarbus (i.e. D. marocanus). Benovics et al. (Reference Benovics, Nejat, Abdoli and Šimková2021a) previously discussed the phylogenetic relationships of the species within lineage C; however, these new findings might finally resolve the origin and dispersal of Dactylogyrus species into Africa. Cyprinids of the genus Garra can be found throughout Southwest Asia with a distribution range extending from the Indus River up to the Nile basin and Ethiopia (Stiassny and Getahun, Reference Stiassny and Getahun2007). The distribution and phylogeography of the genus might suggest that Garra species served as the historical mediator for the dispersion of Dactylogyrus between Eastern Asia and Africa. By similarities in their morphology, we can expect that other specialists of Garra (e.g. D. rectotrabus) will also belong to this phylogenetic lineage; however, additional sequences of Dactylogyrus parasitizing African and Asian Garra are required to test this hypothesis.
Similarly to Dactylogyrus, Gyrodactylus species in Iraq did not form a monophyletic group. Overall, the gyrodactylid species studied herein clustered in 3 distinct lineages of different origins, i.e. Palearctic and African. Gyrodactylus vukicae n. sp. and G. satanicus n. sp. found to parasitize G. rufa were grouped within lineage A, whilst G. satanicus n. sp. was genetically closer to G. hildae from African O. niloticus than to its congener from the same host. Well-supported monophyly was found for G. mhaiseni n. sp., Gyrodactylus sandai n. sp. and the undescribed Gyrodactylus sp. 3 and Gyrodactylus sp. 4 collected from A. sellal and B. lacerta, respectively. Specifically, the sister position of G. mhaiseni n. sp. to Gyrodactylus sp. 3 might suggest that these species are strongly associated with their Alburnus hosts in the Middle East and might represent a case of intrahost duplication on A. sellal. Previously, Huyse and Volckaert (Reference Huyse and Volckaert2005) revealed that Gyrodactylus species parasitizing the gills originated from host-switch and that only less host-specific fin Gyrodactylus co-speciated with their goby hosts, indicating that in their host–parasite system cospeciation is not associated with high host specificity in viviparous parasites. Moreover, they showed that the host switching of Gyrodactylus from Gasterosteus aculeatus Linnaeus, 1758 to non-congeneric fish hosts most likely facilitated the adaptive radiation of numerous highly host-specific Gyrodactylus species. According to Hahn et al. (Reference Hahn, Weiss, Stojanovski and Bachmann2015), cophylogenetic patterns are trackable also at the population level of Gyrodactylus parasites of G. aculeatus. Their study also supported host-switch as a common event in the evolutionary history of Gyrodactylus. In our case, we can expect that intrahost duplication will play an important role in the speciation of Gyrodactylus in geographically isolated regions, just as in Thaparocleidus (Šimková et al., Reference Šimková, Serbielle, Pariselle, Vanhove and Morand2013) and Cichlidogyrus (Mendlová et al., Reference Mendlová, Desdevides, Civáňová, Pariselle and Šimková2012). The distribution of A. sellal is rather widespread in the Middle East, as this species can be found in the rivers of the Mediterranean and Red Sea basins (e.g. Bogutskaya, Reference Bogutskaya1997; Kuru, Reference Kuru2004; Dağlı and Erdemli, Reference Dağli and Erdemli2009; Erk'akan and Özdemir, Reference Erk'akan and Özdemir2011; Bi̇reci̇kli̇gi̇l et al., Reference Bi̇reci̇kli̇gi̇l, Yücel and Çiçek2016), where we can expect that different monogenean species evolved in the individual parapatric populations independently. Alburnus sellal was also parasitized by G. iraqemembranatus n. sp., a highly genetically and morphologically divergent Gyrodactylus species. This species was recorded on 3 phylogenetically non-congeneric host species, all collected in the Tabin River basin (however, at different collection sites) and represents a phylogenetically divergent, basally positioned, lineage. We can only assume that other endemic Middle Eastern Gyrodactylus spp. will share the basal position with G. iraqemembranatus n. sp.; however, molecular data on local species are still scarce. The 3 distinct lineages of the analysed Iraqi Gyrodactylus revealed by genetic data are, in fact, in accordance with the haptoral morphology of the gyrodactylid species, we studied rather than with host phylogeny, similar pattern as was observed in African Cichlidogyrus (Rahmouni et al., Reference Rahmouni, Vanhove, Koblmüller and Šimková2022), or in Nearctic Dactylogyrus (Šimková et al., Reference Šimková, Verneau, Gelnar and Morand2006). Except for the morphologically unidentified species included in the phylogenetic analyses, all Gyrodactylus belonging to lineage A exhibited a similar morphotype of ventral bar, mainly characterized by the presence of lateral processes, a feature lacking in G. blazeki n. sp. and their congeners of lineage D, as well as in G. iraqemembranatus n. sp., which was positioned as the most basal lineage F. Except for G. vukicae n. sp., the species of lineage A identified in this study on endemic cyprinoids from Iraq showed ventral bars with a ridge in the median part of the membrane, a well-known characteristic of Eurasian gyrodactylid lineages, identified as the G. katharineri group in Malmberg (Reference Malmberg1970) – based on records of G. katharineri from a wide range of cyprinoids in this region (Pugachev et al., Reference Pugachev, Gerasev, Gussev, Ergens and Khotenowsky2009). In this study, G. sandai n. sp. was discriminated from G. katharineri using DNA sequences following Huyse et al. (Reference Huyse, Audenaert and Volckaert2003) and Ziȩtara and Lumme (Reference Ziȩtara and Lumme2003), while no haptoral features were found to morphologically differentiate between these 2 species. This may indicate a cryptic speciation as previously found in the Nearctic system (Rahmouni et al., Reference Rahmouni, Seifertová and Simková2023). Finally, G. iraqemembranatus n. sp. possesses a ‘simple’ form of haptoral sclerites, with a ventral bar lacking both lateral processes and a membrane, although it possesses the typical shape of marginal hooks, especially with prolonged basal part. Therefore, we can hypothesize that Gyrodactylus species with such features (see the diagnosis section for G. iraqemembranatus n. sp.) form a basal clade together with G. iraqemembranatus n. sp.; however, their molecular data are required for future phylogenetic studies.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182023001348.
Data availability statement
The data supporting the conclusions of this study are included in this article. The type-material of the new species described in this study was deposited in the Helminthological Collection of the Institute of Parasitology, Czech Academy of Sciences, České Budĕjovice, Czech Republic under the accession number IPCAS M-782 – 793. The newly generated sequences were submitted to the GenBank database under accession numbers OR773082 – OR773094, OR817699 – OR817715, and OR817682 – OR817698.
Acknowledgements
We are grateful to Robert Míč for his help with the fish examination and parasite collection. We are also indebted to Radek Šanda and Jasna Vukić (Natural Museum and Charles University, respectively, Czech Republic) for the fish collection in the field, and for their identification. Further thanks go to Mohammed Azeez Saeed and Salahaddin University in Erbil for hospitality and providing opportunity for sampling in Iraq. We kindly thank Matthew Nicholls for English revision of the final draft.
Author contributions
M. B. and A. Š. conceived and designed the study. M. B., C. R. and F. N. collected parasitological material. E. Ř. performed morphological analyses and prepared the descriptions for Dactylogyrus parasites, C. R. performed morphological analyses and prepared the descriptions for Gyrodactylus parasites, M. B. and F. N. performed molecular laboratory procedures and M. B. performed phylogenetic and statistical analyses. M. B. put together the results and wrote the draft. All the other authors revised the draft and approved the final version.
Financial support
This study was funded by the Czech Science Foundation, project n. GA20-13539S.
Competing interests
None.
Ethical standards
All applicable institutional, national and international guidelines for the care and use of animals were followed. This study was approved by the Animal Care and Use Committee of the Faculty of Science, Masaryk University in Brno (Czech Republic).





















 Open access
Open access