



Introduction
Balamuthia mandrillaris is an opportunistic free-living amoeba responsible for granulomatous amoebic encephalitis (GAE), a rare but often fatal disease of the central nervous system (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Bhosale and Parija, Reference Bhosale and Parija2021; Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022). Since its first recognition as a human pathogen, Balamuthia has been associated with high mortality rates, diagnostic delay and limited therapeutic success (Cope et al. Reference Cope, Landa, Nethercut, Collier, Glaser, Moser, Puttagunta, Yoder, Ali and Roy2019; Siddiqui et al. Reference Siddiqui, Rizvi, Maciver and Khan2025). Unlike Naegleria fowleri, which produces an acute and rapidly fatal meningoencephalitis, Balamuthia infection typically follows a subacute or chronic course, evolving over weeks to months (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; McCormick-Baw et al. Reference McCormick-Baw, Pence and Filkins2023). This prolonged disease trajectory reflects fundamental differences in routes of invasion, tissue tropism and host immune interaction (Siddiqui et al. Reference Siddiqui, Rizvi, Maciver and Khan2025). Comparative pathology across species provides an essential framework for understanding the conserved mechanisms by which B. mandrillaris invades neural tissue, induces inflammation and disrupts vascular integrity. However, a systematic comparison of the neuropathological features of B. mandrillaris infection across humans, naturally infected animals and experimental models remains limited. By examining pathological features shared between human cases, naturally infected animals and experimental models, it will be possible to distinguish core disease mechanisms from host-specific modifiers and to assess the translational relevance of available models. This narrative comparative review was based on literature identified through searches of PubMed, Scopus and Google Scholar using combinations of the terms ‘Balamuthia mandrillaris’, ‘granulomatous amoebic encephalitis’, ‘neuropathology’, ‘comparative pathology’ and the names of affected species including human, dog, horse, tiger, non-human primate and rodent. Representative human cases, naturally infected animals and experimental studies providing neuropathological detail were prioritized. We describe neuropathological findings across species to define the characteristic pathological signature of Balamuthia encephalitis.
Pathogenesis and routes of central nervous system invasion
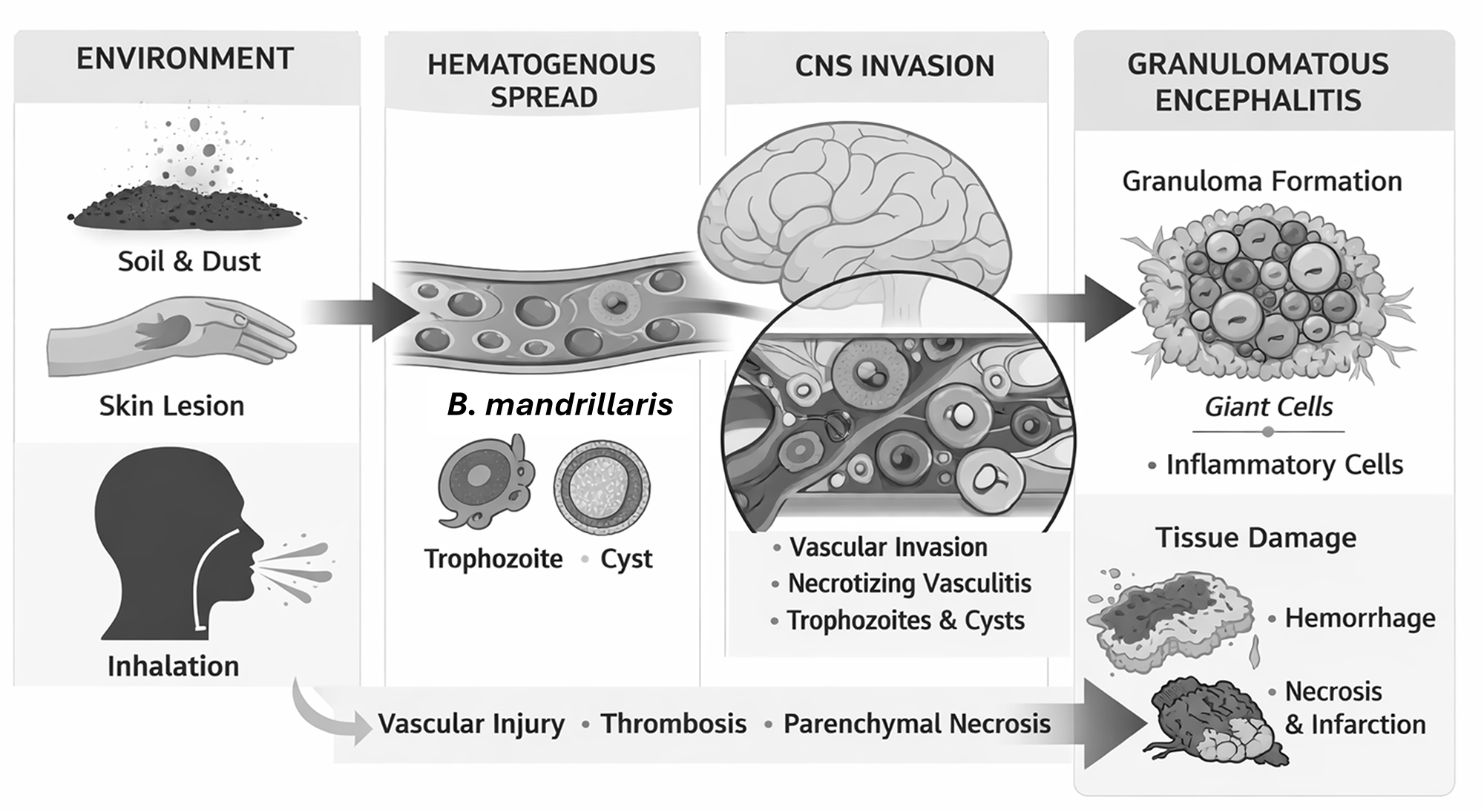
Given the free-living nature of B. mandrillaris, infection is contracted most commonly through the skin or respiratory tract (Cope et al. Reference Cope, Landa, Nethercut, Collier, Glaser, Moser, Puttagunta, Yoder, Ali and Roy2019; Zhang et al. Reference Zhang, Liang, Wei, Feng, Wang, Wang, Zhao, Yu, Gu and Yao2022). Cutaneous lesions frequently precede neurological disease in humans, and inhalation of environmental cysts or trophozoites has also been proposed as an entry route (Alvarez et al. Reference Alvarez, Torres-Cabala, Gotuzzo and Bravo2022). Histopathological examination revealed that these granulomatous cutaneous lesions showed histiocytes, lymphocytes and plasma cells infiltration (Lehmer et al. Reference Lehmer, Ulibarri, Ragsdale and Kunkle2017; Wang et al. Reference Wang, Cheng, Li, Jian, Qi, Sun and Gao2020). Following peripheral entry, the organism disseminates haematogenously and interacts with vascular endothelium to access the brain (Siddiqui and Khan, Reference Siddiqui and Khan2008). Unlike N. fowleri, which exploits the olfactory nerve to bypass the blood–brain barrier, Balamuthia appears to cross the barrier through endothelial invasion, paracellular disruption or vascular damage (Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022). Unlike N. fowleri, both B. mandrillaris and Acanthamoeba spp. produce a more subacute or chronic GAE with haematogenous dissemination and the presence of both trophozoites and cysts in tissue. However, B. mandrillaris demonstrates a more marked perivascular or angiocentric distribution, with trophozoites commonly observed around blood vessels and within vessel walls, together with prominent necrotizing vasculitis and granulomatous inflammation (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Siddiqui et al. Reference Siddiqui, Ortega-Rivas and Khan2008). In contrast to the predominantly neutrophilic and rapidly destructive inflammatory response associated with N. fowleri, Balamuthia infection is characterized by a more chronic granulomatous response with macrophages, lymphocytes, multinucleated giant cells and prominent vascular injury (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022). Balamuthia trophozoites accumulate around blood vessels (Wang et al. Reference Wang, Cheng, Li, Jian, Qi, Sun and Gao2020), where they induce endothelial injury, provoke inflammatory cell recruitment and initiate necrotizing vasculitis (Bhosale and Parija, Reference Bhosale and Parija2021; Zhang et al. Reference Zhang, Liang, Wei, Feng, Wang, Wang, Zhao, Yu, Gu and Yao2022). The mechanisms behind the tendency for Balamuthia to be restricted to the perivascular regions are not presently understood. One possibility however, is that Balamuthia may be less able than Naegleria and Acanthamoeba to tolerate anaerobic conditions. Balamuthia produces proteases and other cytotoxic factors that contribute directly to tissue destruction, while host-mediated immune responses amplify injury through granuloma formation and vascular compromise. Importantly, Balamuthia retains the capacity to encyst within mammalian tissue (Wang et al. Reference Wang, Cheng, Li, Jian, Qi, Sun and Gao2020; Bhosale and Parija, Reference Bhosale and Parija2021), and both trophozoites and cysts are consistently observed in affected brains (Figure 1). This ability to persist in a dormant form distinguishes Balamuthia from Naegleria and likely contributes to the chronicity of disease. Figure 1 summarizes the major features of B. mandrillaris infection, including environmental exposure, haematogenous dissemination, vascular invasion, granulomatous inflammation and tissue destruction.
Schematic overview of the conserved pathogenic sequence of Balamuthia mandrillaris infection. Environmental exposure to B. mandrillaris occurs primarily through soil or dust, with entry via skin breaches or inhalation. Following peripheral entry, amoebae disseminate haematogenously and circulate as trophozoites and cysts. Central nervous system invasion occurs through interactions with the cerebral vasculature, resulting in endothelial injury and penetration of the blood–brain barrier. Within the brain, B. mandrillaris exhibits marked angiocentric tropism, accumulating around blood vessels and inducing necrotizing vasculitis, thrombosis and perivascular inflammation. The ensuing host response is characterized by granulomatous inflammation with macrophages and multinucleated giant cells, leading to multifocal parenchymal necrosis, haemorrhage and infarction. This sequence of events is conserved across mammalian hosts despite interspecies differences in disease kinetics and immune context.

Pathology in humans and animals
Human disease
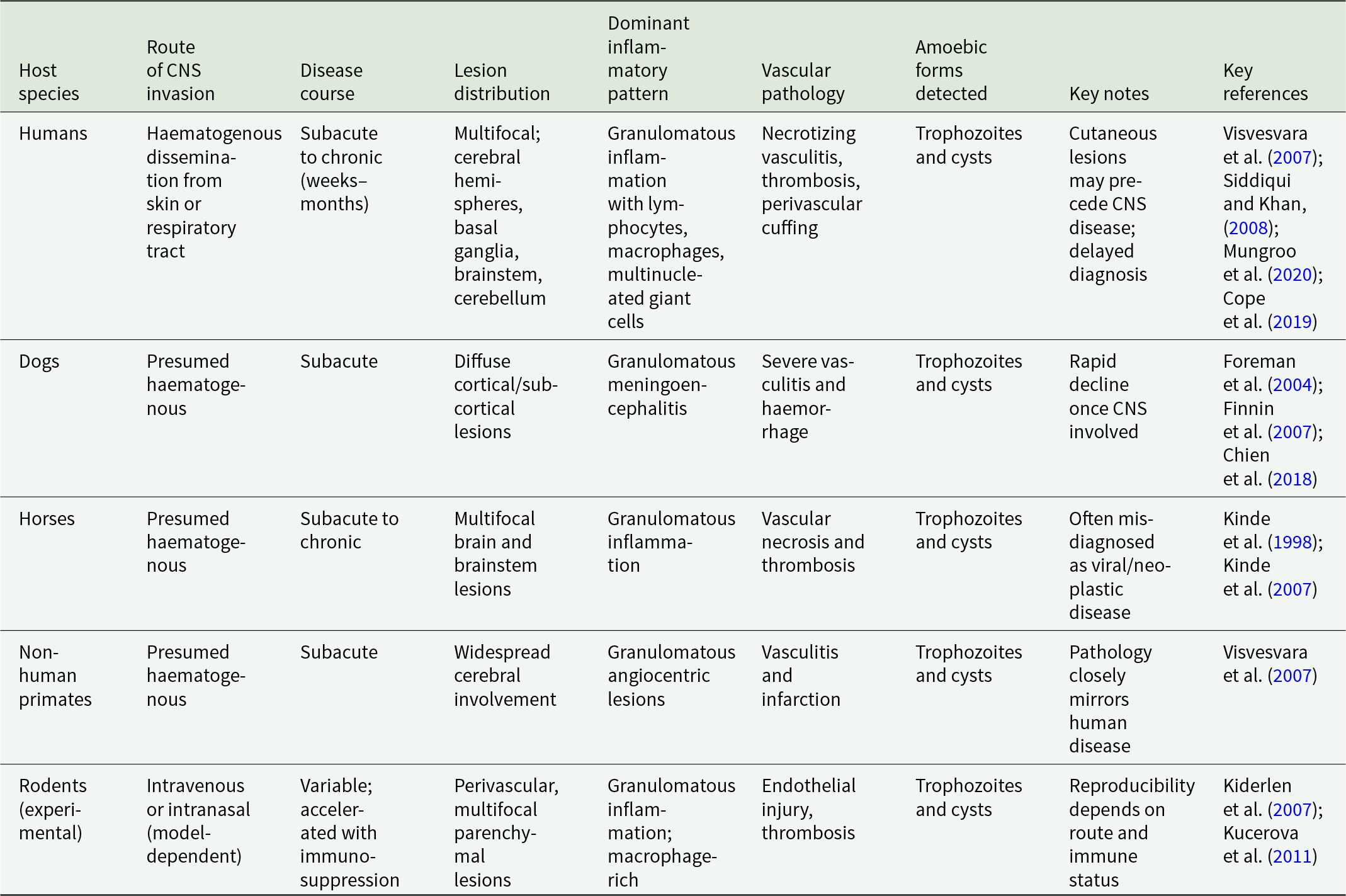
In humans, GAE caused by B. mandrillaris is characterized by multifocal lesions distributed throughout the cerebral hemispheres, basal ganglia, brainstem and cerebellum (Mungroo et al. Reference Mungroo, Khan and Siddiqui2020). Gross examination often reveals cerebral oedema, focal haemorrhage and areas of softening corresponding to infarction or necrosis. Lesions are frequently asymmetric and may mimic neoplastic, vascular or inflammatory conditions on neuroimaging (Javed et al. Reference Javed, Hussain, Ghanchi, Gilani and Enam2024; Chen et al. Reference Chen, Zhang and Lou2025). Microscopically, the defining feature is granulomatous inflammation centred on blood vessels. Perivascular cuffs composed of lymphocytes, macrophages, plasma cells and multinucleated giant cells are prominent (McKeever, Reference McKeever2012). Necrotizing vasculitis and thrombosis are common, leading to secondary ischaemic injury. Amoebae are identified within perivascular spaces, necrotic parenchyma and occasionally within vessel walls (Siddiqui and Khan, Reference Siddiqui and Khan2008). Both trophozoites and cysts are present, although organisms may be sparse and difficult to detect without immunohistochemical or molecular techniques (Kucerova et al. Reference Kucerova, Sriram, Wilkins and Visvesvara2011). Definitive identification of B. mandrillaris requires histopathology together with immunohistochemistry, polymerase chain reaction or sequencing-based approaches, particularly when amoebae are sparse or difficult to distinguish morphologically, although these are not discussed further as they are beyond the scope of this review (Kucerova et al. Reference Kucerova, Sriram, Wilkins and Visvesvara2011; McCormick-Baw et al. Reference McCormick-Baw, Pence and Filkins2023). Preceding cutaneous lesions and unexplained multifocal cerebral lesions should raise suspicion for Balamuthia infection (Alvarez et al. Reference Alvarez, Torres-Cabala, Gotuzzo and Bravo2022; Zhang et al. Reference Zhang, Liang, Wei, Feng, Wang, Wang, Zhao, Yu, Gu and Yao2022; Siddiqui et al. Reference Siddiqui, Maciver and Khan2026). The radiological and clinical features of Balamuthia encephalitis may mimic brain tumours, cerebral abscesses, tuberculosis, fungal infection, vasculitis or other amoebic encephalitides, particularly when multifocal ring-enhancing lesions are present. However, the coexistence of granulomatous inflammation, necrotizing vasculitis and both trophozoites and cysts within tissue favours B. mandrillaris infection (Ünal et al. Reference Ünal, Kuzucu, Demirdağ, Aykur, Kiliç, Güdeloğlu, Polat, Al, Kalkanci, Börcek and Tapisiz2024). The coexistence of active inflammation and tissue necrosis reflects a prolonged host–pathogen interaction rather than an overwhelming acute insult (Table 1).
Cross-species neuropathological features of Balamuthia mandrillaris infection

Animal infections and experimental models
Naturally occurring Balamuthia infections have been reported in a range of mammals, including dogs, horses and non-human primates (Kinde et al. Reference Kinde, Visvesvara, Barr, Nordhausen and Chiu1998, Reference Kinde, Read, Daft, Manzer, Nordhausen, Kelly, Fuerst, Booton and Visvesvara2007; Foreman et al. Reference Foreman, Sykes, Ball, Yang and De Cock2004; Finnin et al. Reference Finnin, Visvesvara, Campbell, Fry and Gasser2007; Chien et al. Reference Chien, Telford, Roy, Ali, Shieh and Confer2018; Niedringhaus et al. Reference Niedringhaus, Gordon, Yabsley, Gai, Uzal and Woolard2023). Clinical presentation in animals parallels human disease, with progressive neurological dysfunction and variable extra-cranial involvement. Post-mortem examination characteristically reveals multifocal granulomatous meningoencephalitis with angiocentric distribution (Kinde et al. Reference Kinde, Visvesvara, Barr, Nordhausen and Chiu1998, Reference Kinde, Read, Daft, Manzer, Nordhausen, Kelly, Fuerst, Booton and Visvesvara2007; Foreman et al. Reference Foreman, Sykes, Ball, Yang and De Cock2004; Finni et al. Reference Finnin, Visvesvara, Campbell, Fry and Gasser2007; Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Chien et al. Reference Chien, Telford, Roy, Ali, Shieh and Confer2018; Niedringhaus et al. Reference Niedringhaus, Gordon, Yabsley, Gai, Uzal and Woolard2023). Necrotizing vasculitis, haemorrhage and parenchymal necrosis are common, and amoebae are detected within lesions as both trophozoites and cysts. Experimental infection models, primarily in rodents, reproduce key pathological features of human disease, although outcomes depend strongly on inoculation route and host immune status (Kiderlen et al. Reference Kiderlen, Laube, Radam and Tata2007). Intravenous or intranasal inoculation results in haematogenous dissemination and angiocentric brain lesions resembling those seen in natural infection (Siddiqui and Khan, Reference Siddiqui and Khan2008; Mungroo et al. Reference Mungroo, Khan and Siddiqui2020). Granulomatous inflammation, perivascular amoebae and vascular injury are consistently observed (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Mungroo et al. Reference Mungroo, Khan and Siddiqui2020). Some models require immunosuppression to establish sustained infection (Kiderlen et al. Reference Kiderlen, Laube, Radam and Tata2007), highlighting the importance of host immune surveillance in controlling disease progression. Despite these limitations, experimental models capture the essential features of Balamuthia neuropathology and provide valuable platforms for mechanistic and therapeutic studies (Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022; Siddiqui et al. Reference Siddiqui, Rizvi, Maciver and Khan2025). More recently, human cerebral organoid systems have also been used to model Balamuthia infection and investigate host–pathogen interactions in human neural tissue (Tongkrajang et al. Reference Tongkrajang, Kobpornchai, Dubey, Chaisri and Kulkeaw2024).
Cross-species comparison
Comparison of pathological features across species reveals a striking conservation of disease architecture (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022). In humans, companion animals, livestock and experimental models, B. mandrillaris consistently produces angiocentric lesions characterized by granulomatous inflammation, necrotizing vasculitis and secondary parenchymal injury. The presence of both trophozoites and cysts within neural tissue is a universal finding, underscoring the organism’s capacity for persistence within the mammalian brain (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Bhosale and Parija, Reference Bhosale and Parija2021). Despite this conserved pathological signature, meaningful differences exist in disease kinetics and lesion distribution. Humans often experience prolonged subclinical or prodromal phases before neurological deterioration (Cope et al. Reference Cope, Landa, Nethercut, Collier, Glaser, Moser, Puttagunta, Yoder, Ali and Roy2019; McCormick-Baw et al. Reference McCormick-Baw, Pence and Filkins2023), whereas some animal infections progress more rapidly once central nervous system involvement occurs (Chien et al. Reference Chien, Telford, Roy, Ali, Shieh and Confer2018; Niedringhaus et al. Reference Niedringhaus, Gordon, Yabsley, Gai, Uzal and Woolard2023). Variability in immune competence influences the balance between parasite proliferation and host-driven tissue damage, with immunosuppressed hosts exhibiting more extensive dissemination and reduced granuloma formation (Kiderlen et al. Reference Kiderlen, Laube, Radam and Tata2007; Cope et al. Reference Cope, Landa, Nethercut, Collier, Glaser, Moser, Puttagunta, Yoder, Ali and Roy2019; Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022). Nonetheless, the fundamental mechanisms of vascular invasion, immune-mediated injury and tissue necrosis remain consistent across species, although disease course, lesion distribution and inflammatory pattern vary between hosts (Table 1).
Environmental niche and exposure pathways
Although B. mandrillaris is widely distributed in soil and dust, clinically apparent infection is exceedingly rare (Dunnebacke et al. Reference Dunnebacke, Schuster, Yagi and Booton2004; Parija and Giri, Reference Parija and Giri2012; Yamanouchi et al. Reference Yamanouchi, Arima, Sakamoto, Kanto, Kasai, Ito and Inaba2018). The environmental reservoir and ecological niche of the organism remain incompletely defined, and the factors governing transition from environmental exposure to invasive disease are poorly understood. The frequent association of human infection with cutaneous lesions and the demonstration of haematogenous spread across species suggest that skin and respiratory exposure represent key portals of entry (Siddiqui and Khan, Reference Siddiqui and Khan2008; Alvarez et al. Reference Alvarez, Torres-Cabala, Gotuzzo and Bravo2022; Zhang et al. Reference Zhang, Liang, Wei, Feng, Wang, Wang, Zhao, Yu, Gu and Yao2022). The rarity of disease despite widespread environmental presence indicates that host susceptibility, immune status and possibly genetic factors play critical roles in determining outcome (Cope et al. Reference Cope, Landa, Nethercut, Collier, Glaser, Moser, Puttagunta, Yoder, Ali and Roy2019; Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022; Siddiqui and Khan, Reference Siddiqui and Khan2025).
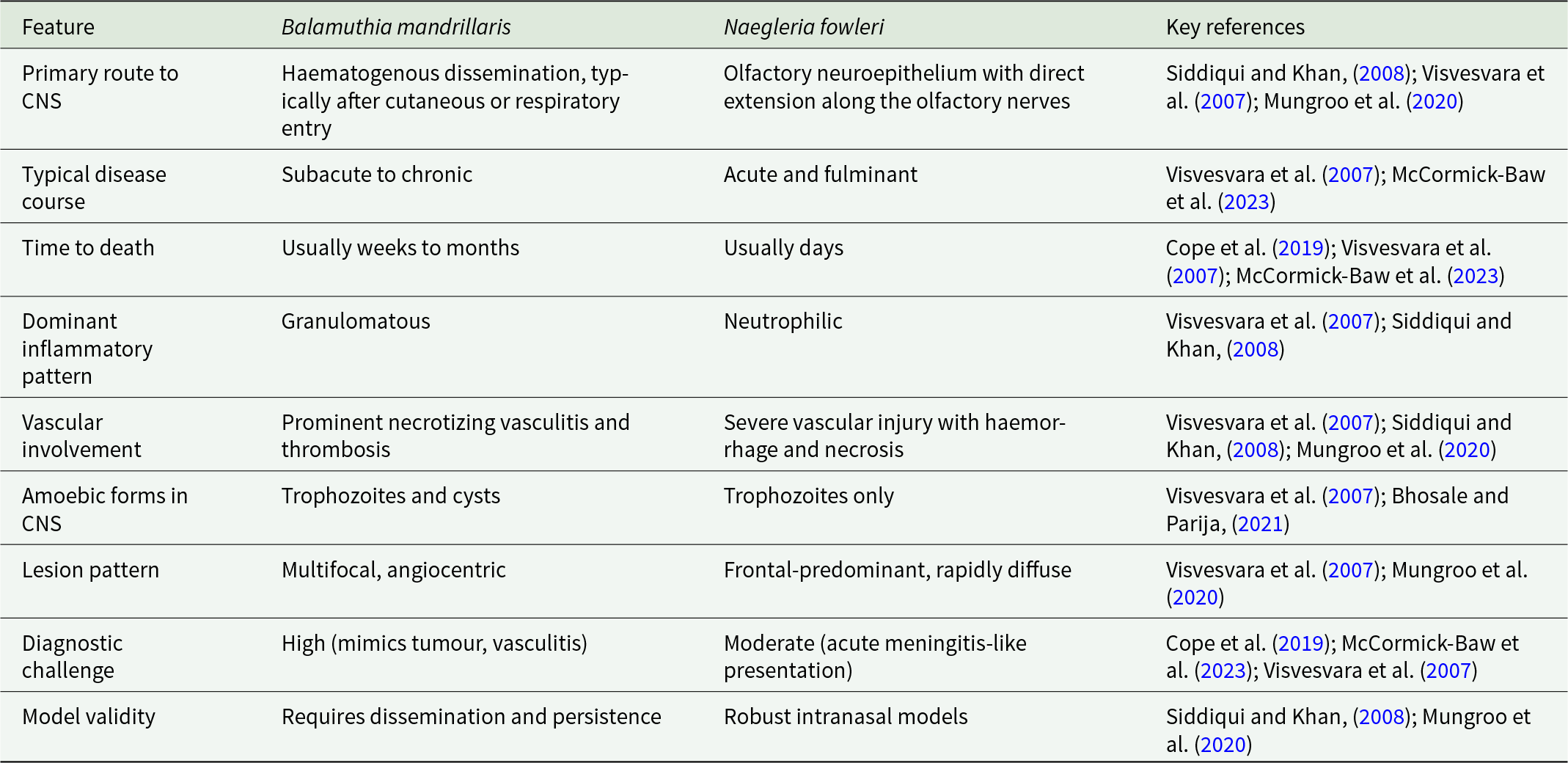
The remarkable consistency of Balamuthia neuropathology across species confirms the translational relevance of animal models for studying GAE (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007; Mungroo et al. Reference Mungroo, Khan, Maciver and Siddiqui2022). Conserved features such as angiocentric invasion, necrotizing vasculitis and cyst persistence identify vascular injury and chronic host–pathogen interaction as central drivers of disease (Siddiqui and Khan, Reference Siddiqui and Khan2008; Mungroo et al. Reference Mungroo, Khan and Siddiqui2020; Bhosale and Parija, Reference Bhosale and Parija2021). These mechanisms differ fundamentally from the acute cytolytic pathology of Naegleria infection (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007) and have important implications for diagnosis and therapy. These differences between B. mandrillaris and N. fowleri are summarized in Table 2. Future studies should prioritize models that faithfully reproduce haematogenous dissemination, granulomatous inflammation and cyst biology. Improved understanding of endothelial interactions, immune modulation and cyst resilience may identify therapeutic vulnerabilities that can be exploited across species, as illustrated by efforts to develop Balamuthia-specific antibody fragments for detection and potential targeting of the parasite (Siddiqui et al. Reference Siddiqui, Kulsoom, Lalani and Khan2016). Current treatment relies on prolonged multidrug therapy, most commonly involving pentamidine, sulfadiazine, flucytosine, azithromycin or clarithromycin, fluconazole or itraconazole, and miltefosine, sometimes combined with neurosurgical intervention, although outcomes remain poor in most cases (Mungroo et al. Reference Mungroo, Khan and Siddiqui2020; Siddiqui and Khan, Reference Siddiqui and Khan2020; McCormick-Baw et al. Reference McCormick-Baw, Pence and Filkins2023). Early recognition of extra-cranial disease, particularly cutaneous manifestations, may also offer opportunities for timely intervention before irreversible neurological damage occurs (Alvarez et al. Reference Alvarez, Torres-Cabala, Gotuzzo and Bravo2022; Zhang et al. Reference Zhang, Liang, Wei, Feng, Wang, Wang, Zhao, Yu, Gu and Yao2022).
Key neuropathological and clinical contrasts between Balamuthia mandrillaris and Naegleria fowleri

In conclusion, comparative analysis demonstrates that B. mandrillaris produces a conserved pattern of granulomatous, angiocentric encephalitis across mammalian hosts (Visvesvara et al. Reference Visvesvara, Moura and Schuster2007). Regardless of species, the organism disseminates haematogenously; targets the cerebral vasculature; and induces chronic inflammation, vascular injury and multifocal parenchymal destruction. The consistent presence of trophozoites and cysts highlights sustained tissue persistence as a defining feature of disease. These shared pathological features validate the use of animal models for mechanistic investigation and therapeutic development and provide a framework for advancing diagnosis and treatment of GAE.
Acknowledgements
RS, NAK and SKM are supported by the Air Force Office of Scientific Research (AFOSR), USA. An AI-based language editing tool was used to improve the grammar and readability of this manuscript. All scientific content, analysis and interpretation were performed by the authors, who take full responsibility for the final manuscript.
Author contributions
RS and NAK conceptualized the study amid discussions with DL and SKM. RS and NAK carried out the study and prepared the first draft of the manuscript. DL and SKM added to the manuscript. All authors approved the final manuscript.
Financial support
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Competing interests
The authors have stated explicitly that there are no conflicts of interest in connection with this article.
Ethical standards
Not applicable.

 Open access
Open access