



Introduction
Hypersaline environments represent unique ecological systems shaped by extreme abiotic conditions, where only highly specialized and uniquely adapted organisms can survive (Colletti, Reference Colletti2014). Despite their apparent simple and reduced biodiversity, these ecosystems play a crucial role in local ecological balance, hosting complex food webs in which a limited number of taxa support multiple trophic interactions (Saccò et al., Reference Saccò, White, Harrod, Salazar, Aguilar, Cubillos, Meredith, Baxter, Oren, Anufriieva, Shadrin, Marambio-Alfaro, Bravo-Naranjo and Allentoft2021). Among the key components of these systems are brine shrimp of the genus Artemia (Leach, 1819; Branchiopoda: Anostraca), which act as primary consumers and constitute an essential link between primary producers and higher trophic levels (Gajardo and Beardmore, Reference Gajardo and Beardmore2012). Brine shrimp are filter-feeders primarily consuming microalgae and planktonic bacteria, thus regulating their populations and contributing to nutrient cycling (Belovsky et al., Reference Belovsky, Stephens, Perschon, Birdsey, Paul, Naftz, Baskin, Larson, Mellison, Luft, Mosley, Mahon, Van Leeuwen and Allen2011; Sánchez et al., Reference Sánchez, Paredes, Lebouvier and Green2016). At the same time, they serve as important food resources for several waterbird species (Sánchez et al., Reference Sánchez, Green and Castellanos2006; Dolmans et al., Reference Dolmans, Partoens, Deboelpaep, Horváth and Vanschoenwinkel2025), thereby establishing intricate ecological interactions, including predator–prey dynamics and multiple host–parasite relationships (Sánchez et al., Reference Sánchez, Nikolov, Georgieva, Georgiev, Vasileva, Pankov, Paracuellos, Lafferty and Green2013). Notably, brine shrimp are recognized as intermediate hosts for numerous avian cyclophyllidean cestodes (Georgiev et al., Reference Georgiev, Sánchez, Green, Nikolov, Vasileva and Mavrodieva2005, Reference Georgiev, Sánchez, Vasileva, Nikolov and Green2007; Amarouayache et al., Reference Amarouayache, Derbal and Kara2009; Sánchez et al., Reference Sánchez, Nikolov, Georgieva, Georgiev, Vasileva, Pankov, Paracuellos, Lafferty and Green2013; Redón et al., Reference Redón, Berthelemy, Mutafchiev, Amat, Georgiev and Vasileva2015; Sivakumar et al., Reference Sivakumar, Hyland and Schuster2020). Although various studies on cestode–Artemia interactions originate from the American continent (Young, Reference Young1952; Redón et al., Reference Redón, Berthelemy, Mutafchiev, Amat, Georgiev and Vasileva2015, Reference Redón, Vasileva, Georgiev and Gajardo2019; Guagliardo et al., Reference Guagliardo, Graff, Gigola and Tanzola2020), most of them regard the Mediterranean area where the cestode Flamingolepis liguloides (Gervais, 1847) is particularly relevant in Artemia populations (Amat et al., Reference Amat, Illescas and Fernandez1991a; Georgiev et al., Reference Georgiev, Sánchez, Vasileva, Nikolov and Green2007, Reference Georgiev, Angelov, Vasileva, Sánchez, Hortas, Mutafchiev, Pankov and Green2014; Redón et al., Reference Redón, Amat, Hontoria, Vasileva, Nikolov and Georgiev2011; Sánchez et al., Reference Sánchez, Nikolov, Georgieva, Georgiev, Vasileva, Pankov, Paracuellos, Lafferty and Green2013). The parasite’s life cycle is closely intertwined with the ecology of brine shrimp and flamingos (Phoenicopteridae), which are among their avian predators. Artemia acquires infection while filter-feeding, ingesting cestode eggs containing an oncosphere larva (Robert and Gabrion, Reference Robert and Gabrion1991). The oncosphere penetrates the intestinal wall and develops into a cysticercoid in the hemocoel (Sánchez et al., Reference Sánchez, Georgiev and Green2007). Transmission to the definitive host occurs when infected brine shrimp are consumed by flamingos, where the parasite reaches maturity and releases eggs into the environment. Flamingolepis liguloides has been reported from multiple Artemia populations across the Mediterranean region – such as France (Robert and Gabrion, Reference Robert and Gabrion1991) and Spain (Amat et al., Reference Amat, Illescas and Fernandez1991a; Georgiev et al., Reference Georgiev, Sánchez, Vasileva, Nikolov and Green2007) – as well as from more distant regions, including Tengiz Lake in Kazakhstan (Maksimova, Reference Maksimova1973) and the Godolphin Lakes in Dubai (Sivakumar et al., Reference Sivakumar, Hyland and Schuster2020). In Italy, evidence for the presence of this cestode is scarce, with the only published records dating back to the early 1990s in southwestern Sardinia (Di Cave et al., Reference Di Cave, Cirillo, Dettori and Mura1990; Di Cave and Mura, Reference Di Cave and Mura1990; Mura, Reference Mura1995). To date, no additional occurrences have been reported elsewhere in the country. To fill this gap, the present study investigates the presence of F. liguloides in Artemia sp. within the hypersaline lagoon of ‘Saline dei Monaci’ (Apulia Region, Italy). The aim is to provide the first evidence of this parasite in the local Artemia population and to contribute new information on host–parasite interactions in Mediterranean hypersaline ecosystems.
Materials and methods
Study area
The study was conducted in the Salina dei Monaci (40°18′7.281″ N, 17°43′54.768″ E), a coastal hypersaline lagoon located in Manduria (Taranto, Apulia Region, Southern Italy). Formerly a saltwork, it is a shallow basin (20–70 cm) that depends mainly on rainfall for its filling, after which evaporation leads to progressive concentration and the creation of hypersaline conditions. Salinity and temperature vary seasonally, ranging from 40 gL−1 in winter to 290 gL−1 in summer and from 5–6 °C in winter to 37 °C in summer, respectively (Mura et al., Reference Mura, Baxevanis, Lopez, Hontoria, Kappas, Moscatello, Fancello, Amat and Abatzopoulos2005). Following the cessation of salt extraction in the mid-20th century, the area has developed into an ecologically valuable wetland supporting numerous resident and migratory waterbirds and is currently designated as a Special Area of Conservation (SAC IT IT9130001 ‘Torre Colimena’) due to its botanical and ecological relevance (Mastrolia et al., Reference Mastrolia, Fiocca, Pinna and Basset2017). Together with the adjacent Dunes of Torre Colimena, the area is protected by ‘Riserve Naturali Regionali Orientate del Litorale Tarantino Orientale’.
Sample collection and parasitological analyses
A random sample of Artemia sp. individuals was collected in October 2023 from the Salina dei Monaci, using a plankton hand net with a 125 μm mesh size. Live specimens were transported to the CISMAR laboratories (Ichthyogenic Experimental Marine Centre, Tuscia University) in 30 L aerated plastic containers to ensure survival during transport. Within 24 h of sampling, 254 randomly selected Artemia individuals were identified to species level using morphological keys (Mura and Brecciaroli, Reference Mura and Brecciaroli2004), measured for total length to the nearest 0.1 mm under a stereomicroscope (Leica M205 FCA) and examined for parasitic infections. Cysticercoids were isolated and counted in each infected host and identified to species level based on diagnostic morphological characters described by Georgiev et al. (Reference Georgiev, Sánchez, Green, Nikolov, Vasileva and Mavrodieva2005).
Molecular identification of host and parasite
To confirm the species identity of both host and parasite, total genomic DNA was extracted from 15 brine shrimp individuals and 10 F. liguloides cysticercoids randomly selected from the overall sample using the Quick-gDNA Miniprep Kit (Zymo Research), following the manufacturer’s instructions. A partial fragment of the mitochondrial cytochrome c oxidase subunit I (cox1) gene was amplified from each brine shrimp specimen using the primers 1/2COI_Fol-F (5′-ATT CTA CGA ATC ACA AGG ATA TTG G-3′) and 1/2COI_Fol-R (5′-TAC ACT TCA GGA TGG CCA AAA AAT CA-3′) (Muñoz et al., Reference Muñoz, Gómez, Green, Figuerola, Amat and Rico2008). Polymerase chain reactions (PCRs) were carried out in a 25 μL volume containing 0.6 μL of each primer 10 μM (Promega), 2 μL of MgCl2 25 mM (Promega), 5 μL of 5× buffer (Promega), 0.6 μL of dNTPs 10 mM (Promega), 0.2 μL of Go-Taq Polymerase (5 U μL−1) (Promega) and 2 μL of total DNA. PCR temperature conditions were as follows: an initial denaturation at 94 °C for 3 min, followed by 40 cycles of denaturation at 94 °C for 45 sec, annealing at 55 °C for 1 min and extension at 72 °C for 1 min, with a final extension at 72 °C for 5 min.
For each F. liguloides cysticercoid, a partial fragment of the small ribosomal subunit gene (SSU, 18S) was amplified using the primers HymF1 (5′-GAT CAA TTG GAG GGC AAG T-3′) and HymR1 (5′-CTG TCA TGA CGG TGA TTG A-3′) (Redón et al., Reference Redón, Quiroz, Lukić, Green and Gajardo2024). The partial large ribosomal subunit (LSU, 28S) was also amplified for the first time using the primers ZX-1 (5′-ACCCGCTGAATTTAAGCATAT-3′) and 1500R (5′-GCT ATC CTG AGG GAA ACT TCG-3′) (Van der Auwera et al., Reference Van der Auwera, Chapelle and De Wächter1994; Palm et al., Reference Palm, Waeschenbach, Olson and Littlewood2009). Both PCRs were carried out in a 25 μL volume containing 0.6 μL of each primer 10 μM (Promega), 2 μL of MgCl2 25 mM (Promega), 5 μL of 5× buffer (Promega), 0.6 μL of dNTPs 10 mM (Promega), 0.2 μL of Go-Taq Polymerase (5 U μL−1) (Promega) and 2 μL of total DNA. PCR temperature conditions were as follows: an initial denaturation at 95 °C for 5 min, followed by 40 cycles of denaturation at 95 °C for 30 sec, annealing at 55 °C (28S) and 54 °C (18S) for 30 sec and extension at 72 °C for 2 min, with a final extension at 72 °C for 7 min. The successful PCR products were purified, and Sanger sequenced through an Automated Capillary Electrophoresis Sequencer 3730 DNA Analyzer (Applied Biosystems), using the Big-Dye® Terminator v3.1 Cycle Sequencing Kit (Life Technologies). Forward and reverse sequences obtained were assembled using Sequencher v. 5.0.1 (GeneCodes Co.) and compared with reference sequences using BLASTn (Morgulis et al., Reference Morgulis, Coulouris, Raytselis, Madden, Agarwala and Schäffer2008).
Statistical analyses
Prevalence (P: proportion of infected individuals), mean abundance (MA: mean number of cysticercoids per host examined) ± standard deviation (± SD) and mean intensity (MI: mean number of cysticercoids in infected hosts) ± SD were calculated following Bush et al. (Reference Bush, Lafferty, Lotz and Shostak1997) on the total sample of individuals examined (N = 254).
The relationship between brine shrimp total length and parasite abundance was assessed using linear regression. Based on a preliminary graphical analysis, we fitted 2 models: one with parasite abundance as a linear term (y ∼ x) and one including a quadratic term (y ∼ x + x 2). These models were compared using the second-order Akaike’s information criterion (AICc; Burnham and Anderson, Reference Burnham and Anderson2002). The model with the lowest AICc was considered the best, provided that its AICc differed from competing models by at least 2 units (Burnham and Anderson, Reference Burnham and Anderson2002).
In addition, the relationship between the number of cysticercoids in each body part (head, thorax, abdomen, ovisac) and the total parasite abundance was assessed using a generalized linear model with a negative binomial error distribution (Zuur et al., Reference Zuur, Ieno and Smith2007; Lindén and Mäntyniemi, Reference Lindén and Mäntyniemi2011). Specifically, we modelled the number of cysticercoids per body part as the response variable, including the total parasite abundance, the body length and interactions between total parasite abundance × body part and body length × body part as predictors. Model support was evaluated by comparing the AICc of this model with that of a null model (i.e. without predictors, y ∼ 1; Burnham and Anderson, Reference Burnham and Anderson2002). Multicollinearity was assessed using variance inflation factors (VIFs), model fit was quantified using pseudo R 2 (Nakagawa and Schielzeth, Reference Nakagawa and Schielzeth2013) and the normality of residuals was checked with a Kolmogorov–Smirnov test. All statistical analyses were performed in R version 4.2.2 (R Core Team, 2022) and the packages MuMIn (Bartoń, Reference Bartoń2018), glmmTMB (Brooks et al., Reference Brooks, Kristensen, van Benthem, Magnusson, Berg, Nielsen, Skaug, Machler and Bolker2017) and DHARMa (Hartig, Reference Hartig2022).
Results
Identification of parthenogenetic Artemia and F. liguloides
Morphological analyses of the brine shrimp collected from the Salina dei Monaci confirmed their assignment to the genus Artemia based on diagnostic characters, including the typical anostracan body plan, 11 pairs of thoracopods and the absence of males, all consistent with parthenogenetic populations (Browne et al., Reference Browne, Sorgeloos and Trotman1991; Figure 1A). Partial sequences of the mitochondrial cox1 gene were successfully obtained from all 15 Artemia individuals analysed (fragment length: 600 bp). BLAST comparisons revealed that all sequences shared 100% identity with reference sequences of Artemia parthenogenetic lineage deposited in GenBank (MT791655; Rode et al., Reference Rode, Jabbour-Zahab, Boyer, Flaven, Hontoria, Van Stappen, Dufresne, Haag and Lenormand2022), confirming their molecular identification.
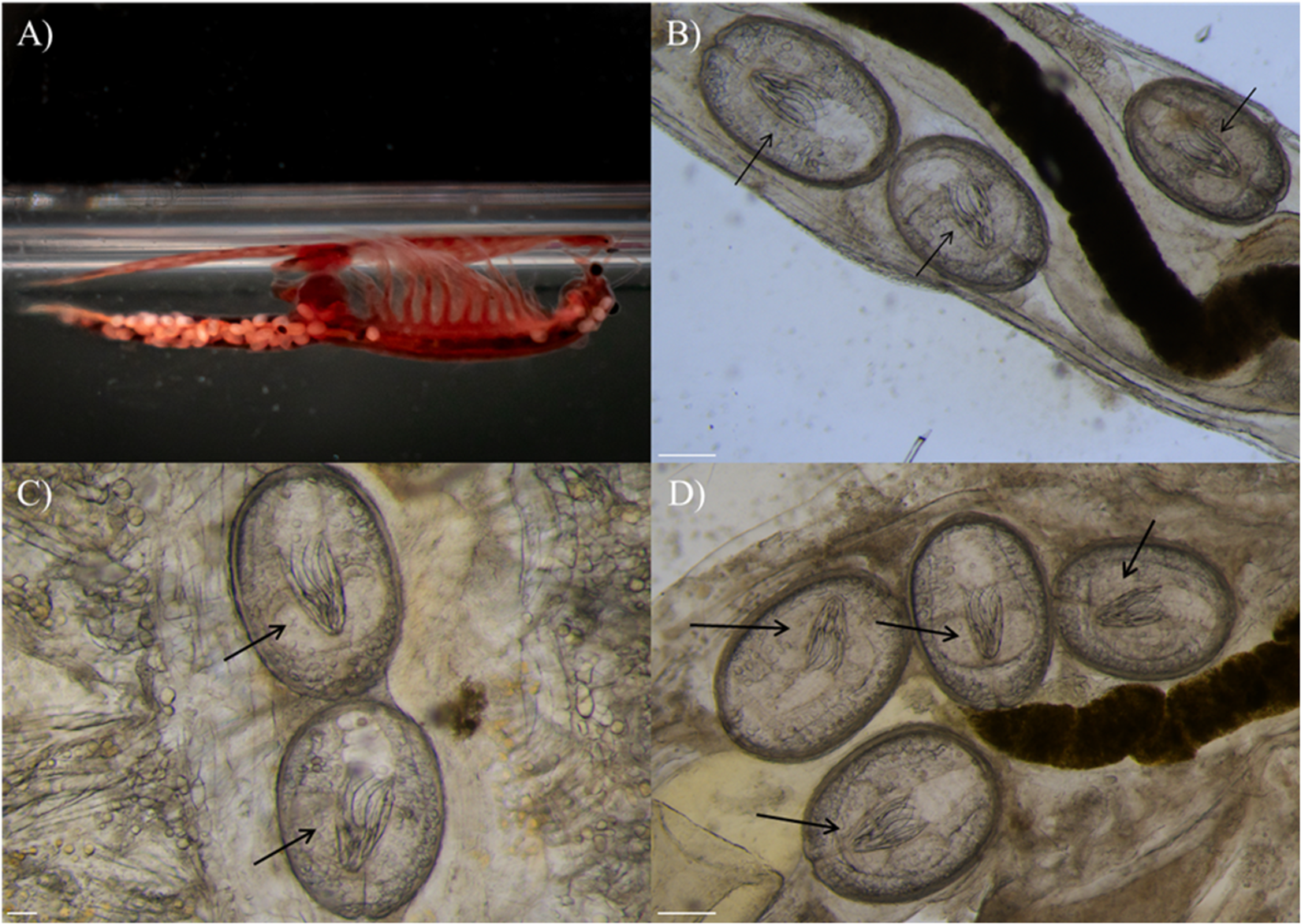
Spatial distribution of Flamingolepis liguloides cysticercoids in parthenogenetic Artemia. (A) General view illustrating cysticercoids distributed throughout the whole body, captured with a Nikon D3500 camera, equipped with a 55 mm 3.5 Micro-Nikkor Macro lens. (B–D) Examples of cysticercoids in specific anatomical regions with the rostellar apparatus visible and indicated by black arrows, captured using a Leica M205 FCA stereomicroscope: (B) abdomen (white scale bar = 250 µm); (C) thorax (white scale bar = 100 µm) and (D) near the ovisac (white scale bar = 250 µm).

Figure 1 Long description
The image A shows a specimen in a tube. The image B shows a microscopic view with arrows pointing to oval shapes. The image C shows a microscopic view with arrows pointing to oval shapes. The image D shows a microscopic view with arrows pointing to oval shapes.
Morphological examination of cestode specimens identified diagnostic features consistent with F. liguloides according to Georgiev et al. (Reference Georgiev, Sánchez, Green, Nikolov, Vasileva and Mavrodieva2005), including a rostellar apparatus with 8 slender hooks (∼130 μm total length; Figure 1B–D). Partial sequences of the ribosomal 18S and 28S genes were successfully obtained from all 10 F. liguloides individuals analysed (fragment length: 1100 bp and 1516 bp, respectively). BLAST comparisons revealed that all 18S sequences shared 100% identity with reference sequences of F. liguloides deposited in GenBank (OR995639; Redón et al., Reference Redón, Quiroz, Lukić, Green and Gajardo2024). BLAST comparisons of 28S sequences shared 88.6% identity with the cestode Hymenolepis microstoma deposited in GenBank (LC064144; Nkouawa et al., Reference Nkouawa, Haukisalmi, Li, Nakao, Lavikainen, Chen, Henttonen and Ito2016). The newly obtained sequences for parthenogenetic Artemia and F. liguloides have been deposited in GenBank under accession numbers PZ315227 (cox1) and PZ315369–PZ315425 (18S–28S, respectively).
Parasitic infection and site preferences
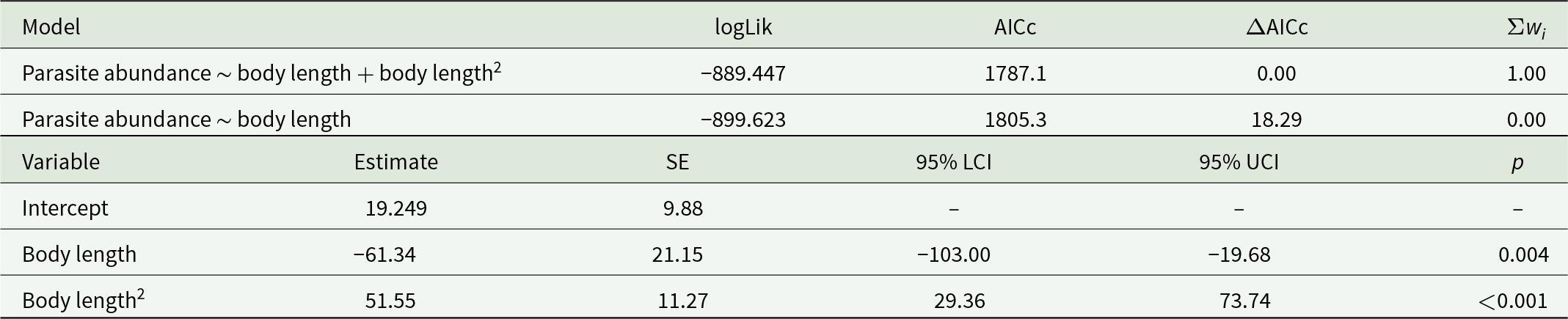
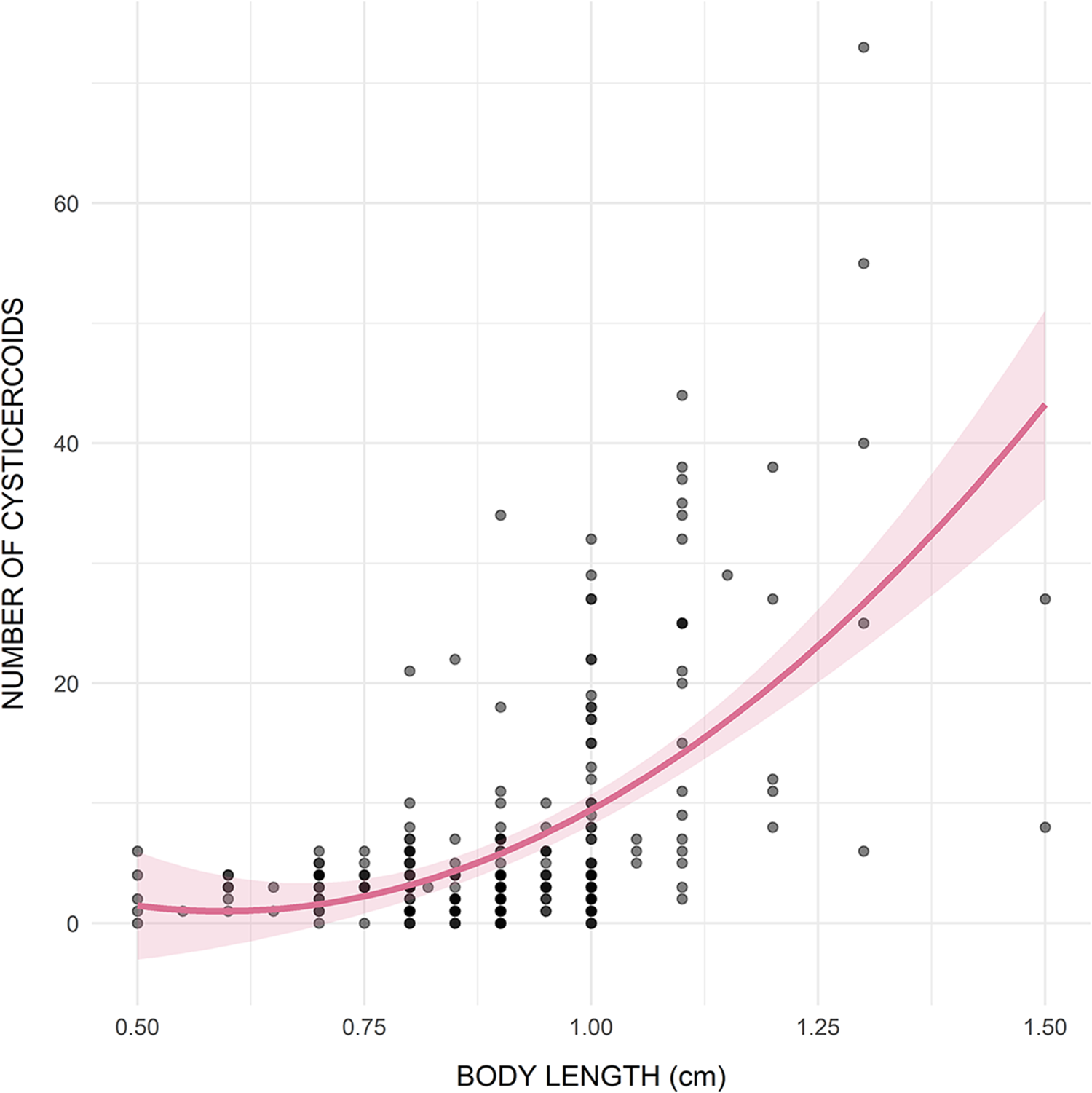
Infection with F. liguloides was detected in 93.3% of the examined brine shrimp individuals. Mean abundance was 7.3 ± 10.1 cysticercoids per host, while mean intensity reached 7.8 ± 10.3 cysticercoids per infected host (range: 1–73), with a maximum of 73 cysticercoids in a single individual. Parasite abundance significantly increased with host body length, with the best-fitting model including a positive quadratic term (Table 1; R 2 = 0.343) (Figure 2).
Comparison of models investigating the relationship between the total number of cysticercoids of F. liguloides and the body size of parthenogenetic Artemia. Comparison between linear and quadratic models showed that the latter provided the best fit

Table 1 Long description
The table compares two models linking parasite abundance to host body length: a linear model and a model that also includes body length squared. Model fit metrics show the body length plus squared term model is strongly supported, with log-likelihood minus 889.447, AICc 1787.1, delta AICc 0.00, and Akaike weight 1.00. The linear-only model fits much worse, with log-likelihood minus 899.623, AICc 1805.3, delta AICc 18.29, and weight 0.00. For the best-supported model, the intercept estimate is 19.249 with standard error 9.88, and confidence limits and p-value are not reported. The body length term is negative (estimate minus 61.34, standard error 21.15) with a 95% confidence interval from minus 103.00 to minus 19.68 and p-value 0.004. The body length squared term is positive (estimate 51.55, standard error 11.27) with a 95% confidence interval from 29.36 to 73.74 and p-value less than 0.001. Together, the negative length term and positive squared term indicate a curved relationship rather than a straight-line trend.
Reported are the log-likelihood (logLik), second-order Akaike’s information criterion (AICc), differences in AICc (ΔAICc) and Akaike weights (Σwi). Summary statistics of the best-supported model include parameter estimates, standard errors (SEs), 95% confidence intervals (LCI: lower, UCI: upper) and their significance (P).
Relationship between host body length (cm) and cysticercoid abundance.

Figure 2 Long description
The horizontal axis represents BODY LENGTH (cm), ranging from 0.50 to 1.50. The vertical axis represents NUMBER OF CYSTICERCOIDS, ranging from 0 to 60. The plot shows a positive correlation, where the number of cysticercoids generally increases with body length. A curved line indicates the trend, with a shaded band representing the confidence interval. Data points are densely clustered around body lengths of 0.75 to 1.00 cm with lower cysticercoid counts, while higher counts are observed at greater body lengths. Notable outliers appear at body lengths above 1.25 cm with cysticercoid counts exceeding 50. The plot aims to demonstrate that larger body lengths are associated with higher cysticercoid abundance.
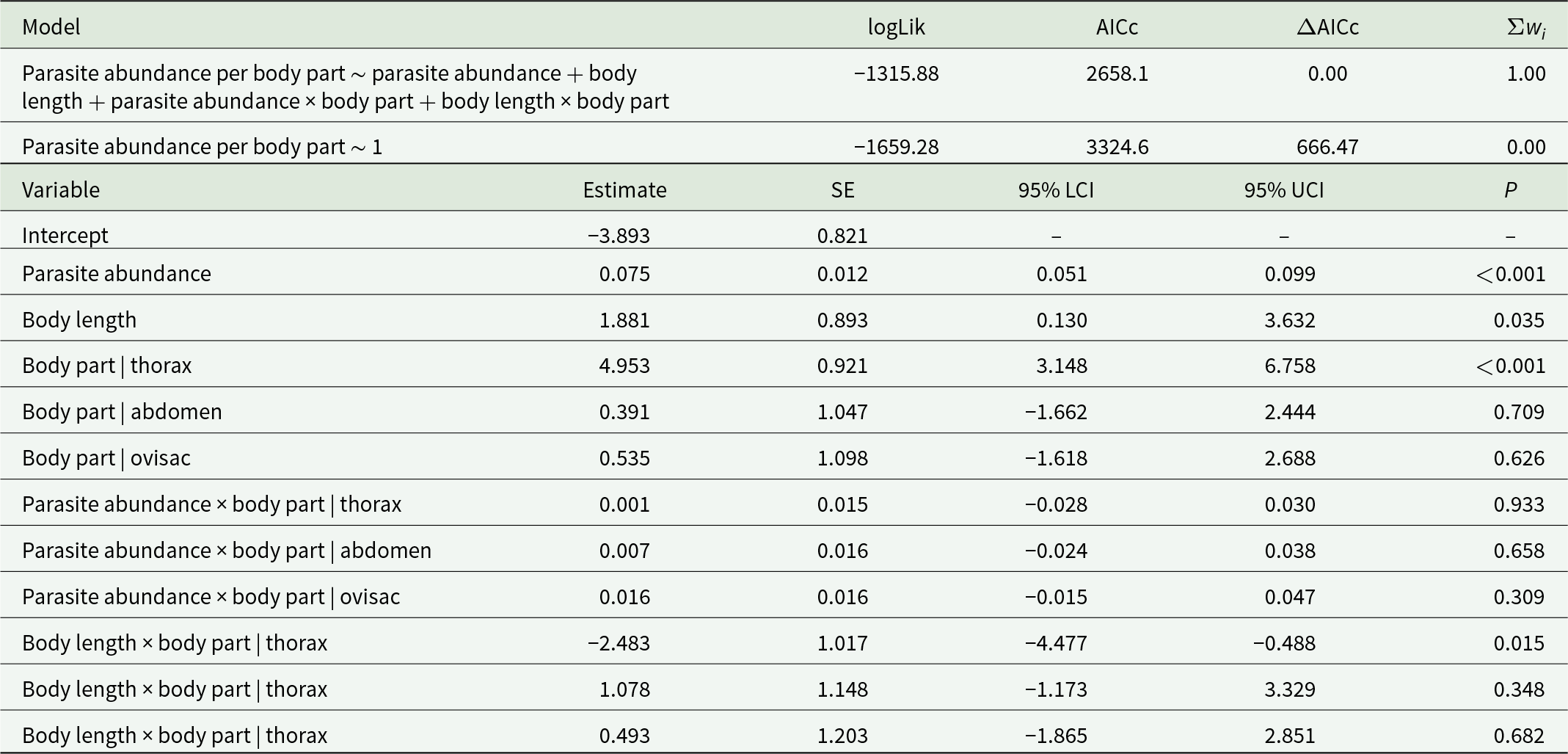
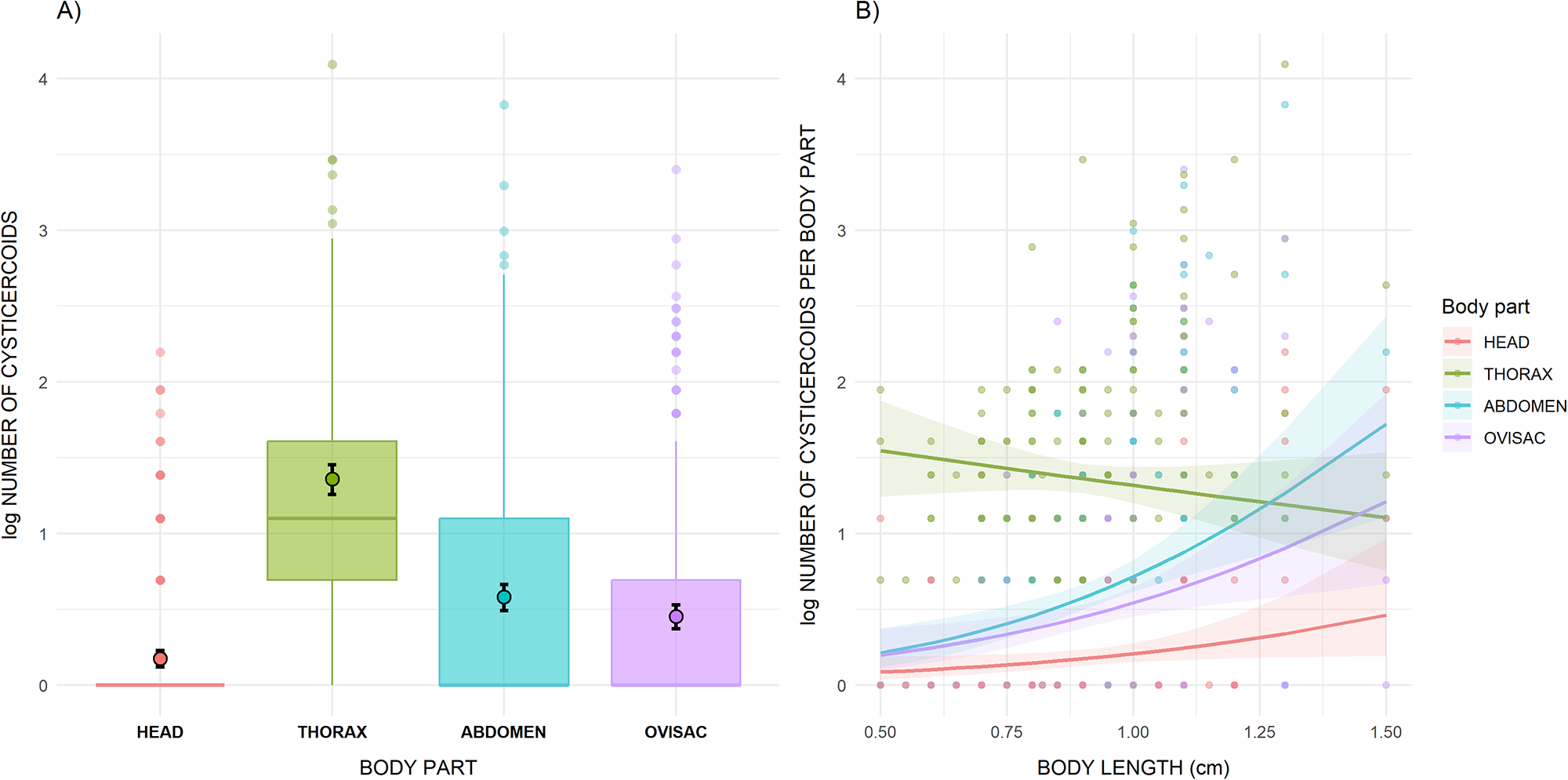
Cysticercoids were distributed within the host body, occurring predominantly in the thorax (55.2%), followed by the abdomen (22.6%), the ovisac region (17.0%) and the head (5.3%). According to the negative binomial model (Table 2), the thorax harboured significantly (p < 0.001) more cysticercoids than the head, abdomen or ovisac (Figure 3A). However, the significant interaction between body length and body part indicates that in larger brine shrimp, which hosted a higher cysticercoid abundance, the relative number of cysticercoids in the thorax decreases compared to the other body parts (Figure 3B). The model showed no collinearity (VIF < 3), had a good level of explained variance (R 2 = 0.604) and exhibited normally distributed residuals (Kolmogorov–Smirnov test, p = 0.544). None of the infected brine shrimp, at any level of infection, contained cysts or nauplii in the ovisac, whereas cysts were observed exclusively in uninfected individuals. Because the number of cyst-bearing individuals was very low, statistical significance could not be assessed.
Model comparison assessing the effects of body part (head, thorax, abdomen, ovisac), total cysticercoid abundance, body length of parthenogenetic Artemia and their interactions on the number of F. liguloides cysticercoids per body part

Table 2 Long description
The table compares two statistical models predicting the number of cysticercoids per body part using body part, total cysticercoid abundance, body length, and their interactions. The full model had log-likelihood minus 1315.88 and AICc 2658.1, while the null model had log-likelihood minus 1659.28 and AICc 3324.6, a very large AICc difference of 666.47; the full model carried essentially all model weight. In the best model, total abundance was positively associated with per-part counts (estimate 0.075, confidence interval 0.051 to 0.099, p less than 0.001). Body length also showed a positive main effect (estimate 1.881, confidence interval 0.130 to 3.632, p 0.035). Relative to the head reference, the thorax had much higher counts (estimate 4.953, confidence interval 3.148 to 6.758, p less than 0.001), while abdomen and ovisac differences were not statistically supported (p 0.709 and 0.626). Interactions between total abundance and body part were small and not significant for thorax, abdomen, or ovisac (all p values above 0.30). The body length interaction for the thorax was negative and significant (estimate minus 2.483, confidence interval minus 4.477 to minus 0.488, p 0.015), indicating the length effect differs in the thorax compared with the head. The final two body-length-by-body-part rows appear to be mislabeled as thorax; interpret those specific labels cautiously.
Comparison with the null model indicated that several predictors contributed to explaining variation in parasite counts. Reported are the log-likelihood (logLik), second-order Akaike’s information criterion (AICc), differences in AICc (ΔAICc) and Akaike weights (Σwi). For the best-supported model, we present parameter estimates, standard errors (SEs), 95% confidence intervals (LCI: lower; UCI: upper) and significance levels (p). The head is the reference level for the body part factor.
(A) Abundance of F. liguloides cysticercoids in each body part (head, thorax, abdomen, ovisac) of parthenogenetic Artemia. Filled dots represent values predicted by the model. (B) Relationship between cysticercoid abundance per body part, total cysticercoid number per individual and the body length of parthenogenetic Artemia. Lines represent values predicted by the model, whereas the shaded areas indicate 95% confidence intervals. In both panels, the y-axis is log-transformed to improve visualization.

Figure 3 Long description
The image A showing a box plot labeled A). The horizontal axis is labeled BODY PART with categories HEAD, THORAX, ABDOMEN and OVISAC. The vertical axis is labeled LOG NUMBER OF CYSTICERCOIDS with tick labels 0, 1, 2, 3 and 4. The THORAX box is positioned higher than the other body parts, with its median above 1. The HEAD box is at 0 with points clustered at 0 and a small number of points extending upward to around 2. The ABDOMEN box has a median below 1 with points extending upward to around 4. The OVISAC box has a median below 1 with points extending upward to around 3. The image B showing a scatter plot labeled B). The horizontal axis is labeled BODY LENGTH (mm) with tick labels 0.50, 0.75, 1.00, 1.25 and 1.50. The vertical axis is labeled LOG NUMBER OF CYSTICERCOIDS PER BODY PART with tick labels 0, 1, 2, 3 and 4. A legend titled Body part lists HEAD, THORAX, ABDOMEN and OVISAC, with separate point and line styles for each body part. Multiple points are plotted for each body part across the body-length range. Four smooth lines are shown with shaded bands around them. The THORAX line slopes downward from higher values at shorter body length toward lower values at longer body length. The ABDOMEN line slopes upward across the body-length range. The OVISAC line slopes upward across the body-length range. The HEAD line stays near 0 and rises slightly at longer body length. Across both images, A shows the distribution of log number of cysticercoids by BODY PART and B shows how LOG NUMBER OF CYSTICERCOIDS PER BODY PART varies with BODY LENGTH (mm) for the same body-part categories.
Discussion
This study provides the first confirmed evidence, supported by morphological and molecular data, of a host–parasite association between parthenogenetic Artemia populations – considered a complex of genetically heterogeneous, often polyploid lineages (Asem et al., Reference Asem, Gajardo, Hontoria, Yang, Shen, Rastegar-Pouyani, Padhye and Sorgeloos2024 and references therein) – and F. liguloides in the hypersaline lagoon of Salina dei Monaci (Apulia, Italy), with newly generated 28S rDNA sequences obtained for the parasite. Previous investigations in the Salina dei Monaci area reported the presence of parthenogenetic Artemia but did not document any associated parasite fauna (Moscatello et al., Reference Moscatello, Belmonte and Mura2002; Moscatello and Belmonte, Reference Moscatello and Belmonte2004; Mura et al., Reference Mura, Baxevanis, Lopez, Hontoria, Kappas, Moscatello, Fancello, Amat and Abatzopoulos2005). However, our findings represent the first verified occurrence of F. liguloides in Italy since the early reports from southwestern Sardinia in the 1990s, where the parasite was recorded in bisexual Artemia populations from Su Pallosu pond with low prevalence and abundance (approximately 2–3%; Di Cave et al., Reference Di Cave, Cirillo, Dettori and Mura1990; Di Cave and Mura, Reference Di Cave and Mura1990; Mura, Reference Mura1995).
In contrast to those earlier observations, the population examined in the present study exhibited high infection levels, with a prevalence of approximately 90% and a significant increase in parasite burden with host body size. Such high infection levels strongly suggest elevated environmental parasite pressure and indicate that the life cycle of F. liguloides is fully maintained within the Salina dei Monaci ecosystem. The coexistence of dense parthenogenetic Artemia populations with large aggregations of the definitive host, the greater flamingo, likely plays a key role in sustaining parasite transmission (Sánchez et al., Reference Sánchez, Nikolov, Georgieva, Georgiev, Vasileva, Pankov, Paracuellos, Lafferty and Green2013). During the sampling period (October), approximately 200 flamingos were observed in the lagoon (GC, pers. obs.), consistent with the species’ main seasonal presence and aggregation in Mediterranean wetlands during post-breeding and post-fledging movements (Johnson, Reference Johnson1989; Spina and Volponi, Reference Spina and Volponi2008). Indeed, as flamingo faeces represent the primary source of infective cestode eggs, persistent bird aggregations are expected to substantially increase environmental infection pressure on Artemia populations. This scenario likely did not apply to the studies conducted in the early 1990s (Di Cave et al., Reference Di Cave, Cirillo, Dettori and Mura1990; Di Cave and Mura, Reference Di Cave and Mura1990; Mura, Reference Mura1995). Flamingos were absent from Su Pallosu pond in 1991 (Mura, Reference Mura1995), sampling was carried out between January and March (i.e. outside the main period of flamingo movements) and the species was still relatively uncommon in the Mediterranean at that time, with no breeding colonies in Italy (the first breeding event in Sardinia was recorded in 1993; Grussu, Reference Grussu1993). Consequently, the probability of flamingo movements, and thus parasite dispersal, across Mediterranean wetlands was likely low.
The high infection levels observed in this population revealed a positive relationship between host body size and parasite burden, in line with patterns described for trophically transmitted endoparasites and supporting the hypothesis of parasite accumulation over time (e.g. Cipriani et al., Reference Cipriani, Sbaraglia, Paoletti, Giulietti, Bellisario, Palomba, Bušelić, Mladineo, Nascetti and Mattiucci2018; Gay et al., Reference Gay, Bao, MacKenzie, Pascual, Buchmann, Bourgau, Couvreur, Mattiucci, Paoletti, Hastie, Levsen and Pierce2018; Levsen et al., Reference Levsen, Cipriani, Palomba, Giulietti, Storesund and Bao2022; Palomba et al., Reference Palomba, Marchiori, Tedesco, Fioravanti, Marcer, Gustinelli, Aco-Alburqueque, Belli, Canestrelli, Santoro, Cipriani and Mattiucci2023). In Artemia, which has a relatively short life cycle (approximately 30–40 days; Browne et al., Reference Browne, Sallee, Grosch, Segreti and Purser1984) but feeds continuously by filtration, larger individuals are expected to experience longer exposure times and higher probabilities of ingesting infective stages. Previous studies have shown that susceptibility to infection in Artemia is developmentally constrained by particle ingestion capacity. Early larval stages (stages III and IV, <3 mm) are able to ingest only particles up to 25–30 μm in diameter (Dobbeleir et al., Reference Dobbeleir, Adam, Bossuyt, Bruggeman, Sorgeloos, Persoone, Sorgeloos, Roels and Jaspers1980; Mura, Reference Mura1995; Fernández, Reference Fernández2001), a size range that excludes F. liguloides eggs (typically 40–50 μm). Accordingly, infection is expected to occur only after individuals reach a minimum body size (minimum reported size of infected individuals: 3–4 mm). In the present study, parasitized hosts had a minimum body length of 5 mm, and parasite burden increased progressively with host size above this threshold. Although smaller individuals were not sampled, precluding direct inference on infection at earlier stages, the observed size-dependent increase in parasite load is consistent with cumulative exposure.
As parasite loads increased, spatial constraints within the host appeared to influence cysticercoid distribution. Cysticercoids were predominantly located in the thoracic region, which harboured approximately 55% of the total parasite load, with a maximum of 73 cysticercoids recorded per host, in agreement with previous studies on F. liguloides in Artemia spp. (Mura, Reference Mura1995; Koru, Reference Koru2022). With increasing host size and parasite burden, cysticercoids were increasingly detected in other body regions, including the abdomen, ovisac and head. This progressive expansion across body regions suggests that the thorax represents the primary site of infection, whereas other regions act as secondary sites once the thoracic cavity becomes saturated, as previously proposed for this host–parasite system (Amarouayache et al., Reference Amarouayache, Derbal and Kara2009). Beyond spatial effects, infections were also associated with marked impacts on host reproduction. All parasitized adult exhibited empty ovarian sacs, indicating functional castration. Parasite-induced castration has been repeatedly documented in Artemia infected by cestode cysticercoids (Amat et al., Reference Amat, Illescas and Fernandez1991a,Reference Amat, Illescas and Fernandezb; Sánchez et al., Reference Sánchez, Rode, Flaven, Redón, Amat, Vasileva and Lenormand2012) and is widely interpreted as an adaptive strategy that enhances parasite fitness. By diverting host resources from reproduction to somatic growth and maintenance, castration may increase host longevity and body size, thereby improving parasite survival and increasing the likelihood of trophic transmission to definitive hosts (Amat et al., Reference Amat, Gozalbo, Navarro, Hontoria and Varó1991b).
Taken together, these results indicate the presence of a stable trophic pathway linking intermediate and definitive hosts in the Salina dei Monaci lagoon. Parasites with complex life cycles depend on intact trophic interactions and stable host communities, and disruptions caused by pollution or habitat degradation may interrupt parasite transmission (Marcogliese, Reference Marcogliese2004; Sures, Reference Sures2004; Hudson et al., Reference Hudson, Dobson and Lafferty2006; Palm and Rückert, Reference Palm and Rückert2009). Conversely, high parasite burdens generally occur only in systems where food webs remain structurally intact. In this context, the infection patterns of F. liguloides reinforce the view that trophically transmitted helminths, when characterized by well-resolved life cycles, host specificity and stable occurrence, can act as sensitive bioindicators of ecosystem health and ecological integrity, rather than merely as stressors (Sures, Reference Sures2004, Reference Sures2008; Hudson et al., Reference Hudson, Dobson and Lafferty2006; Palm and Rückert, Reference Palm and Rückert2009; Palomba et al., Reference Palomba, Marchiori, Tedesco, Fioravanti, Marcer, Gustinelli, Aco-Alburqueque, Belli, Canestrelli, Santoro, Cipriani and Mattiucci2023).
Acknowledgements
We thank Alessandro Mariggiò, Gregorio Pecoraro, Patrizio Raffaele Fontana and the staff of the Riserve Naturali Regionali Orientate del Litorale Tarantino Orientale for granting permission to conduct the sample collection. We are also grateful to Dr Claudio Pardo for providing the photograph used in Figure 1A.
Author contributions
M.F.: Conceptualization, methodology, formal analysis, investigation, data curation, writing – original draft, writing – review & editing, visualization. G.C.: Software, formal analysis, data curation, writing – review & editing, visualization. E.D.B.: Methodology, investigation, writing – review & editing. M.M.: Data curation, writing – original draft, writing – review & editing. G.P.: Resources, writing – review & editing, funding acquisition. M.P.: Conceptualization, methodology, validation, investigation, resources, data curation, writing – original draft, writing – review & editing, supervision, project administration, funding acquisition. D.C.: Resources, writing – review & editing, project administration, funding acquisition.
Financial support
The project was implemented under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4, Call for tender No. 3138 of 16 December 2021, rectified by Decree n. 3175 of 18 December 2021 of the Italian Ministry of University and Research funded by the European Union-Next Generation EU. Project code CN_00000033, Concession Decree No.1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP J83C22000860007, Project title ‘National Biodiversity Future Centre-NBFC’; and Mission 4 Component 2 Investment 3.1, Italian Ministry of University and Research funded by the European Union, NextGenerationEU; Project code IR0000035, CUP C63C22000570001, Project title ‘Unlocking the Potential for health and food from the seas’, EMBRC UP.
G.C. acknowledges the support of University of Tuscia, project ECS00000024 Rome Technopole (CUP B83C22002820006, PNRR Missione 4 Componente 2 Investimento 1.5), funded by the European Union – NextGenerationEU.
Competing interests
The authors declare none.
Ethical standards
Not applicable.

 Open access
Open access