



Introduction
Paleoparasitology is an interdisciplinary field dedicated to the study of parasites in archaeological, paleontological, or other contexts of preservation, providing valuable insights into health, ecology and the interactions between human populations and parasites in the past (Ferreira et al., Reference Ferreira, Araújo and Reinhard2011). The first documented evidence dates back to the pioneering work of Marc Armand Ruffer, who in 1910 described Schistosoma haematobium Bilharz, 1852 eggs in the tissues of Egyptian mummies dated to 1250–1100 BC (Ruffer, Reference Ruffer1910). In Brazil, this line of research became consolidated primarily at the Oswaldo Cruz Foundation (FIOCRUZ), where the term ‘Paleoparasitology’ was proposed by Luiz Fernando Ferreira in 1979, marking the beginning of the formal establishment of the discipline (Araujo et al., Reference Araujo, Reinhard, Ferreira and Gardner2008; Ferreira, Reference Ferreira, Ferreira, Araújo and Reinhard2014). Since then, analyses of coprolites, sediments and other archaeological materials have played an important role in identifying intestinal parasites, reconstructing diet and understanding cultural practices of ancient populations, as well as the ecological conditions in which they lived (Iñiguez et al., Reference Iñiguez, Reinhard, Araújo, Ferreira and Vicente2003; Leles et al., Reference Leles, Araújo, Ferreira, Vicente and Iñiguez2008; Iñiguez et al., Reference Iñiguez, Reinhard, Carvalho Gonçalves, Ferreira, Araújo and Paulo Vicente2006; Araújo et al., Reference Araújo, Reinhard, Ferreira, Pucu and Chieffi2013; Le Bailly et al., Reference Bailly, Landolt, Mauchamp and Dufour2014; Mitchell, Reference Mitchell and Mitchell2015; Sianto et al., Reference Sianto, de Miranda Chaves, Teixeira-Santos, Pereira, Godinho, Gonçalves and Santos2018; Guedes et al., Reference Guedes, Borba, Camacho, Neto, Dias and Iñiguez2020; Gurjão et al., Reference Gurjão, Dias, Neto and Iñiguez2024).
Within this framework, investigations of infectious diseases in past populations have advanced substantially through studies of ancient DNA (aDNA), including palaeogenetics and palaeogenomicss, which not only enrich and complement paleoparasitological findings but also expand the understanding of parasite, host and environment relationships (Jaeger et al., Reference Jaeger, Gijón-Botella, Del Carmen Del Arco-Aguilar, Martín-Oval, Rodríguez-Maffiotte, Del Arco-Aguilar, Araújo and Iñiguez2016; Petrigh et al., Reference Petrigh, Martínez, Mondini and Fugassa2019; Iñiguez, Reference Iñiguez, Hoon, Bianucci, Hoon and Bianucci2021; Ledger et al., Reference Ledger, Murchie, Dickson, Kuch, Haddow, Knüsel, Stein, Pearson, Ballantyne, Knight, Deforce, Carroll, Rice, Franconi, Šarkić, Redžič, Rowan, Cahill, Poblome, Palma, Brückner, Mitchell and Poinar2025). These approaches allow the identification of parasitic infections and the characterization of various host attributes, including the distinction between human and animal individuals (Gurjão et al., Reference Gurjão, Brito, Dias, Neto and Iñiguez2025), the determination of matrilineal and patrilineal haplogroups kinship relations (Jaeger et al., Reference Jaeger, Leles, Lima, da Silva, Dias and Iñiguez2012; Ferraz et al., Reference Ferraz, Suarez Villagran, Nägele, Radzevičiūtė, Barbosa Lemes, Salazar-García, Wesolowski, Lopes Alves, Bastos, Rapp Py-Daniel, Pinto Lima, Mendes Cardoso, Estevam, Liryo, Guimarães, Figuti, Eggers, Plens, Azevedo Erler, Valadares Costa, da Silva Erler, Koole, Henriques, Solari, Martin, Serafim Monteiro da Silva, Kipnis, Müller, Ferreira, Carvalho Resende, Chim, da Silva, Borella, Tomé, Müller Plumm Gomes, Barros Fonseca, Santos da Rosa, de Moura Saldanha, Costa Leite, Cunha, Viana, Ozorio Almeida, Klokler, Fernandes, Talamo, DeBlasis, Mendonça de Souza, de Paula Moraes, Elias Oliveira, Hünemeier, Strauss and Posth2023), biological sex and dietary aspects such as the last meal before death (Maixner et al., Reference Maixner, Turaev, Cazenave-Gassiot, Janko, Krause-Kyora, Hoopmann, Kusebauch, Sartain, Guerriero, O’Sullivan, Teasdale, Cipollini, Paladin, Mattiangeli, Samadelli, Tecchiati, Putzer, Palazoglu, Meissen, Lösch, Rausch, Baines, Kim, An, Gostner, Egarter-Vigl, Malfertheiner, Keller, Stark, Wenk, Bishop, Bradley, Fiehn, Engstrand, Moritz, Doble, Franke, Nebel, Oeggl, Rattei, Grimm and Zink2018). Moreover, they enable analyses of intestinal and external microbiota composition, as well as the reconstruction of past environments, providing an integrated view of biological, cultural and ecological interactions, while simultaneously, through palaeogenomicss, contributing to the reconstruction of ancient genomes (Santiago-Rodriguez et al., Reference Santiago-Rodriguez, Fornaciari, Luciani, Dowd, Toranzos, Marota and Cano2015; Granehäll et al., Reference Granehäll, Huang, Tett, Manghi, Paladin, O’Sullivan, Rota-Stabelli, Segata, Zink and Maixner2021; Majander et al., Reference Majander, Pla-Díaz, du Plessis, Arora, Filippini, Pezo-Lanfranco, Eggers, González-Candelas and Schuenemann2024).
Pre-Columbian coastal communities in Brazil exhibited large population aggregations, indicating that these environments provided adequate resources to support their subsistence (Gaspar et al., Reference Gaspar, DeBlasis, Fish, Fish, Silverman and Isbell2008). In Brazil, sambaquis are archaeological sites characterized by the intentional accumulation of shells and faunal remains that, over time, formed monumental structures in the landscape (Gaspar and Mendonça, Reference Gaspar, Mendonça and Gama2013). The recurrent presence of fish and mollusc remains suggests that fishing and mollusc gathering were central activities for these groups (Fish et al., Reference Fish, Blasis, Gaspar and Fish2000; Klokler and Figuti, Reference Klokler and Figuti2001). The availability of marine resources may also have influenced the mobility of coastal populations, as certain resources were seasonally restricted and their absence may have driven human groups to seek other areas (Figuti, Reference Figuti1993).
In Southern and Southeastern Brazil, pre-Columbian populations have traditionally been referred to as ‘fisher-gatherers’, a term often reduced to mollusc collection, which reflected the greater emphasis given to animal remains compared to plant remains (Scheel-Ybert and Boyadjian, Reference Scheel-Ybert and Boyadjian2020). Recent studies suggest that these societies displayed intermediate lifeways, combining diverse subsistence strategies that do not entirely conform to the classical categories of fisher or agricultural communities (Scheel-Ybert et al., Reference Scheel-Ybert, Boyadjian and Capucho2022). Sambaqui archaeological sites in Brazil recently become areas for aDNA research aimed at understanding the health conditions and lifeways of pre-Columbian populations (Cruz Dávalos et al., Reference Cruz Dávalos, Arizmendi Cárdenas, Bravo-Lopez, Neuenschwander, Reis, Bastos, Stenderup, Yediay, Villa-Islas, Reyna-Blanco, Rodrigues-Carvalho, Hünemeier, Allentoft, Amorim, Moreno-Mayar, Ávila-Arcos and Malaspinas2022; Iñiguez et al., Reference Iñiguez, Brito, Guedes and Chaves2022; Ferraz et al., Reference Ferraz, Suarez Villagran, Nägele, Radzevičiūtė, Barbosa Lemes, Salazar-García, Wesolowski, Lopes Alves, Bastos, Rapp Py-Daniel, Pinto Lima, Mendes Cardoso, Estevam, Liryo, Guimarães, Figuti, Eggers, Plens, Azevedo Erler, Valadares Costa, da Silva Erler, Koole, Henriques, Solari, Martin, Serafim Monteiro da Silva, Kipnis, Müller, Ferreira, Carvalho Resende, Chim, da Silva, Borella, Tomé, Müller Plumm Gomes, Barros Fonseca, Santos da Rosa, de Moura Saldanha, Costa Leite, Cunha, Viana, Ozorio Almeida, Klokler, Fernandes, Talamo, DeBlasis, Mendonça de Souza, de Paula Moraes, Elias Oliveira, Hünemeier, Strauss and Posth2023). However, as open-air formations, these sites are particularly vulnerable to taphonomic processes that compromise the preservation of archaeological materials. Key factors include temperature fluctuations, solar exposure, high microbial activity in the soil, elevated rainfall, and water percolation through archaeological layers, all of which can damage parasitic structures and degrade aDNA (Camacho et al., Reference Camacho, Leles, Santiago, Ramos, Uchôa, Bastos, Nunes, Mendonça de Souza and Araújo2016). In this context, palaeogenomicss is the approach on the frontiers of knowledge, in terms of sensitivity, volume and diversity of data recovered, and also, by adopting controlled sampling protocols to minimize contamination risks and ensure the reliability of results (Iñiguez, Reference Iñiguez, Hoon, Bianucci, Hoon and Bianucci2021). The application of multiple analytical methods has optimized the recovery of preserved aDNA fragments, thereby enhancing the informative potential regarding human health and subsistence in archaeological contexts.
Previous paleoparasitological and palaeogenetic studies from our laboratory reported evidence of helminth infection in an individual from the Jabuticabeira II (JABII) sambaqui, located in Santa Catarina, Brazil (Iñiguez et al., Reference Iñiguez, Brito, Guedes and Chaves2022). Palaeogenetic analyses confirmed the presence of Ascaris sp., representing the first molecular record of parasitic infection in a Brazilian sambaqui. Complementary microscopic analysis revealed plant microremains in six individuals, including starch granules of sweet potato and maize, as well as algal spores, fish scales, and diatoms, which are commonly recovered in coastal archaeological contexts. The presence of pollen from the genus Podocarpus, characteristic of Araucaria forests, suggested that these human groups may have moved between coastal and highland areas in search of resources. Such mobility patterns may indicate the complexity of subsistence strategies and the adaptive capacity of these populations in relation to the environmental diversity of the pre-Columbian period (Iñiguez et al., Reference Iñiguez, Brito, Guedes and Chaves2022). These previous findings on parasites and plant microremains at JABII are further complemented by the palaeogenomics approach of Brito and coauthors (Reference Brito, Pires, Parente, Furtado, Wesolowski and Iñiguez2024), which applied high-throughput sequencing (HTS) and computational biological analyses to expand the identification of plant taxa through aDNA analyses, reinforcing the complexity of plant use within the JABII archaeological population.
Recent palaeogenomics investigations of human remains from JABII archaeological site revealed the presence of Treponema pallidum subspecies endemicum Smibert 1984, the causative agent of bejel (Majander et al., Reference Majander, Pla-Díaz, du Plessis, Arora, Filippini, Pezo-Lanfranco, Eggers, González-Candelas and Schuenemann2024). In the study, the authors identified a lineage closely related to the species responsible for bejel, challenging previous paleopathological interpretations regarding the diversity of treponemal subspecies and their traditional geographic distributions. These findings suggest that treponemal infections were already circulating in the Americas before European contact and highlight the importance of HTS for detecting Treponema spp. aDNA. However, the presence of the bacteria that cause syphilis was not confirmed in pre-Columbian populations.
The study of sedimentary ancient DNA (sedaDNA) represents a powerful approach for reconstructing ancient marine ecosystems, enabling the detection of a wide range of taxa and providing an integrated perspective of past biodiversity, while overcoming the limitations of traditional fossil proxies (Nguyen et al., Reference Nguyen, Devendra, Szymańska, Greco, Angeles, Weiner, Ray, Cordier, De Schepper, Pawłowski and Pawłowska2023). As part of an effort to expand the paleoepidemiological knowledge of the Brazilian sambaquis population, the present palaeogenomics study was conducted by applying HTS technique to sedaDNA from the JABII individuals, aimed to recover additional information on the helminth infection and its associations with subsistence strategies and health in pre-Columbian groups.
Materials and methods
Jabuticabeira II sambaqui
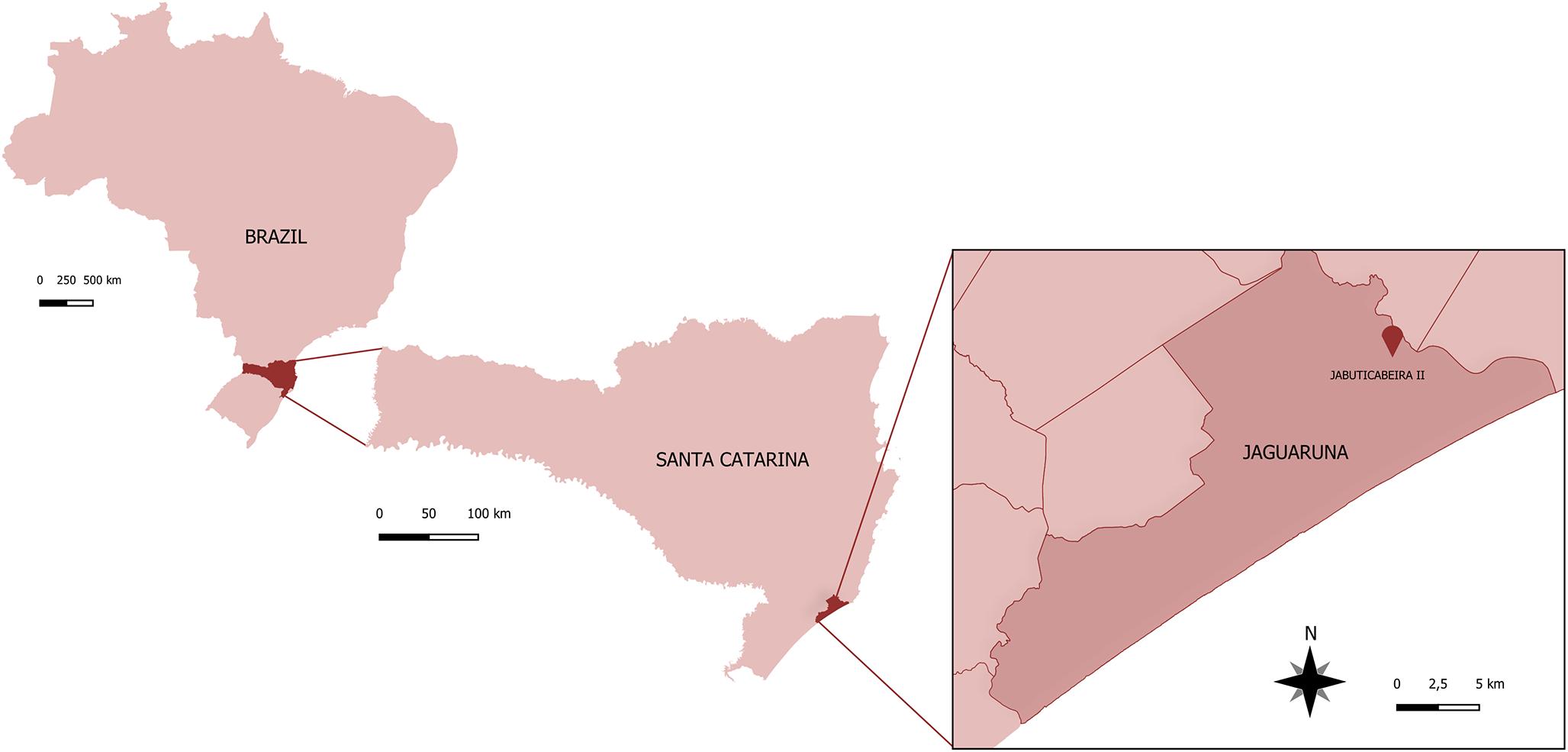
The JABII sambaqui is an archaeological site located in Santa Catarina state, Southern Brazil (Figure 1), with radiocarbon dates indicating an occupation period between 3208–3469 cal yr BP (Beta 270321) and 988–1194 cal yr BP (Beta 234201) (Kneip et al., Reference Kneip, Farias and DeBlasis2018). The archaeological site has a complex stratigraphy, characterized by overlapping and interdigitated layers of shells, sand, and cover layers overlying the funerary deposits (Bianchini et al., Reference Bianchini, Gaspar, DeBlasis and Scheel-Ybert2011). Initial archaeological investigations at JABII began in 1997 through the multidisciplinary project ‘Sambaquis e Paisagens’ (shellmounds and landscapes) led by archaeologists and bioarchaeologists (Blasis et al., Reference Blasis, Kneip, Scheel-Ybert, Giannini and Gaspar2007). The site has yielded a large number of burials, accompanied by numerous artefacts produced or modified by humans. The recovered material culture indicates that JABII functioned predominantly as a funerary space (Bianchini, Reference Bianchini2008).
Geographic location of the archaeological site JABII sambaqui in Jaguaruna municipality, Santa Catarina state, Brazil.

Precautions in working with ancient DNA
Procedures involving aDNA were conducted following strict protocols to ensure authenticity and prevent contamination, as established by previously published guidelines (Iñiguez, Reference Iñiguez, Luna, Aranda and Suby2014, Reference Iñiguez, Hoon, Bianucci, Hoon and Bianucci2021). In 2014, samples from the archaeological site JABII were collected from the osteological collection curated by archaeologist Sabine Eggers at the Instituto de Biociências, Universidade de São Paulo. Sampling was performed using personal protective equipment (PPE), including gloves, lab coats, masks, caps and disposable shoe covers, all replaced for each new collection. Only sterile, disposable instruments were used. Samples were kept protected from light exposure and refrigerated until laboratory processing.
palaeogenetic and palaeogenomics analyses were conducted at the Palaeogenetic Laboratory (PL) from the Laboratório de Parasitologia Integrativa e Paleoparasitologia (LPIP), Instituto Oswaldo Cruz – FIOCRUZ, Rio de Janeiro, Brazil, a facility dedicated exclusively to aDNA studies. The PL facility maintains strict entrance control and has a pre-room for the dressing of all necessary PPE. The PL laboratory is physically compartmentalized into isolated sectors for sample preparation, DNA extraction and PCR reactions, with post-PCR procedures conducted in a separate laboratory to minimize cross-contamination risks. Work areas undergo weekly decontamination protocols using 1% sodium hypochlorite (NaClO) and subsequent ultraviolet light irradiation for at least 1 hour, which may be extended overnight if necessary (Iñiguez et al., Reference Iñiguez, Leles, Jaeger, Carvalho-Costa, Araújo and Group2012). To control and monitor access to research materials and the laboratory, we maintain an mtDNA haplotype ancestry database of each member of the PL staff or archaeologist who is part of the project in progress.
Palaeogenetic and palaeogenomics analyses
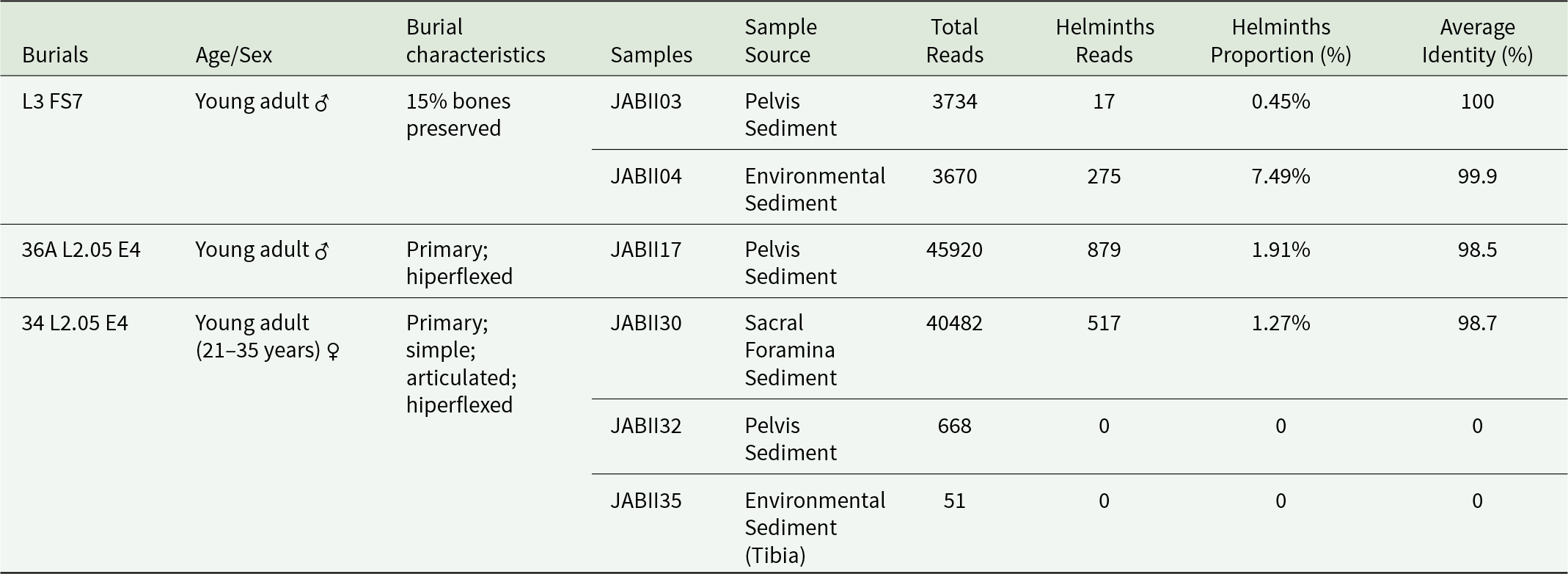
Six sediment samples were collected from the pelvic region and sacral foramina of JABII individuals, along with soil environmental samples (Table 1). These samples originated from 3 burials, encompassing both single and multiple interments, with individuals found in articulated or hyperflexed positions (Table 1). All samples were subjected to appropriate cleaning and decontamination protocols for archaeological materials to minimize contamination risks. Sediment samples were rehydrated in a 0.5% aqueous trisodium phosphate (Na₃PO₄) solution for 72 hours (Callen and Cameron, Reference Callen and Cameron1960) at 4 ºC. Approximately 2–5 g of sediment from each sample was combined with 25 ml of Na₃PO₄ solution. The rehydrated samples were subsequently processed using the spontaneous sedimentation technique (Lutz, Reference Lutz1919), and 200 μL of the resulting sediment was reserved for analysis.
Characteristics of JABII individuals, samples, and results of the Paleogenomic dataset analysis

The sedaDNA extraction was performed using the QIAmp DNA Investigator kit (Qiagen®) following the manufacturer’s protocol with specific modifications tailored for aDNA samples. These modifications included both chemical digestion steps provided by the kit and additional physical digestion, whereby samples were immersed in liquid nitrogen or dry ice and subsequently vortexed to disrupt parasitic structures and enhance DNA release, thereby improving extraction efficiency. Samples were digested overnight with proteinase K (Invitrogen, 20 mg/μL) at 56 °C. aDNA was eluted in two aliquots, yielding a final volume of 80 μL. Extracted DNA samples were then subjected to the reconstructive polymerization technique, a pre-treatment designed to repair and restore fragmented aDNA before amplification (Golenberg et al., Reference Golenberg, Bickel and Weihs1996; Iñiguez et al., Reference Iñiguez, Reinhard, Araújo, Ferreira and Vicente2003). The reactions comprised 1X Buffer, 3.0 mM MgCl2, 0.4 mM dNTP, 2 U of GoTaq G2 Hot Start (Promega, Madison, USA) or Platinum Taq DNA Polymerase (Invitrogen), and 5–10 ng DNA in 25 μL of final volume. The thermocycling was performed in Applied Biosystems™ SimpliAmp™ Thermal Cycler as 94 ºC for 10 min, with 20 cycles of 8 s at 94 °C, 8 s at 56 °C, and 8 s at 72 °C, with an extension of 20 s at 72 °C. Then, nested PCR amplifications were carried out targeting the mitochondrial cox1 gene as the exclusive molecular marker for species identification using the primers described (Botella et al., Reference Botella, Vargas, de la Rosa, Leles, Reimers, Vicente and Iñiguez2010). Reactions were carried out in a final volume of 50 μL, containing 1X Buffer, 2.5 mM MgCl₂, 0.4 mM dNTPs, 100 ng of each primer, 2.5 U of Platinum™ Taq DNA Polymerase (Invitrogen), and 5 μL of aDNA or 0.2 ng/μL of DNA. The cycling conditions for both reactions consisted of an initial denaturation at 96 °C for 5 min, followed by 40 cycles of 40 s at 96 °C, 50 s at 45 °C, and 50 s at 72 °C, with a final extension of 5 min at 72 °C, performed in Applied Biosystems™ SimpliAmp™ Thermal Cycler. PCR amplicons were purified using the ExoSAP-IT™ Express PCR Product Cleanup Kit (Thermo Fisher Scientific), and DNA was quantified with the Quantus™ Fluorometer (Promega).
HTS libraries were prepared using an amplicon metabarcoding approach, in collaboration with the Genomics Unit at the National Cancer Institute (INCA, Brazil). Ancient DNA amplicons were used for library preparation with the TruSeq® Nano DNA Library Prep Kit (Illumina), with protocol adaptations excluding the initial DNA fragmentation step. Sequencing was carried out on the Illumina MiSeq platform. Amplicons from each sample were pooled in equimolar amounts, resulting in a total volume of 30 μL containing 50ng of DNA. Following end-repair, sequencing adapters containing dual indexes were ligated to the fragments, and paired-end libraries were enriched by PCR. The resulting libraries were verified by 1% agarose gel electrophoresis, quantified with a Qubit fluorometer using the High Sensitivity dsDNA Assay Kit (Thermo Fisher Scientific), and diluted to a concentration of 4 nM. Subsequently, the libraries were denatured with NaOH and further diluted to a final concentration of 8 pM. All libraries were then pooled into a single tube and loaded onto the Illumina MiSeq platform for paired-end sequencing (2 × 250 cycles) using the MiSeq Reagent Kit v2, following the protocol of the manufacturer.
In the bioinformatics analyses, the raw paired sequence archives were submitted to a customized script as described (Brito et al., Reference Brito, Pires, Parente, Furtado, Wesolowski and Iñiguez2024; Pires et al., Reference Pires, Furtado, Dias, Neto, Parente and Iñiguez2024), which was developed in the FIOCRUZ Bioinformatics Platform (RPT04A https://plataformas.fiocruz.br/). The quality status of the raw reads was observed with FastQC. The adapters were trimmed, and reads were filtered by 22 minimum quality with AdapterRemoval Version 2. Remaining reads were merged using PEAR Version 0.9.6, and the alignment was performed with BLASTn Version 2.16.0. The alignment was made against a customized database for the cox1 gene, with sequence data available in the GenBank database. The taxonomic identification, alignment cover, and identity were recovered for each read using a script developed in Python language with the pandas package for dataframe manipulation. The Python script produces tables from BLAST output data and calculates proportions based on the best-hit taxa. The analysis of these tables was performed in the R environment using the taxonomizr , dplyr, and tidyr packages. Graphical results were made with ggplot2 package. The aDNA authentication software was not applied since the amplicon metabarcoding approach does not allow the verification of DNA characteristic damage patterns. aDNA authentication methodologies will be performed in the future Jabuticabeira II analyses. The taxonomic groups present in samples were analysed in terms of the burials associated with the JABII sambaqui archaeological site, considering the marine and lagoon environment context and the parasite life cycle of taxa recovered.
Results
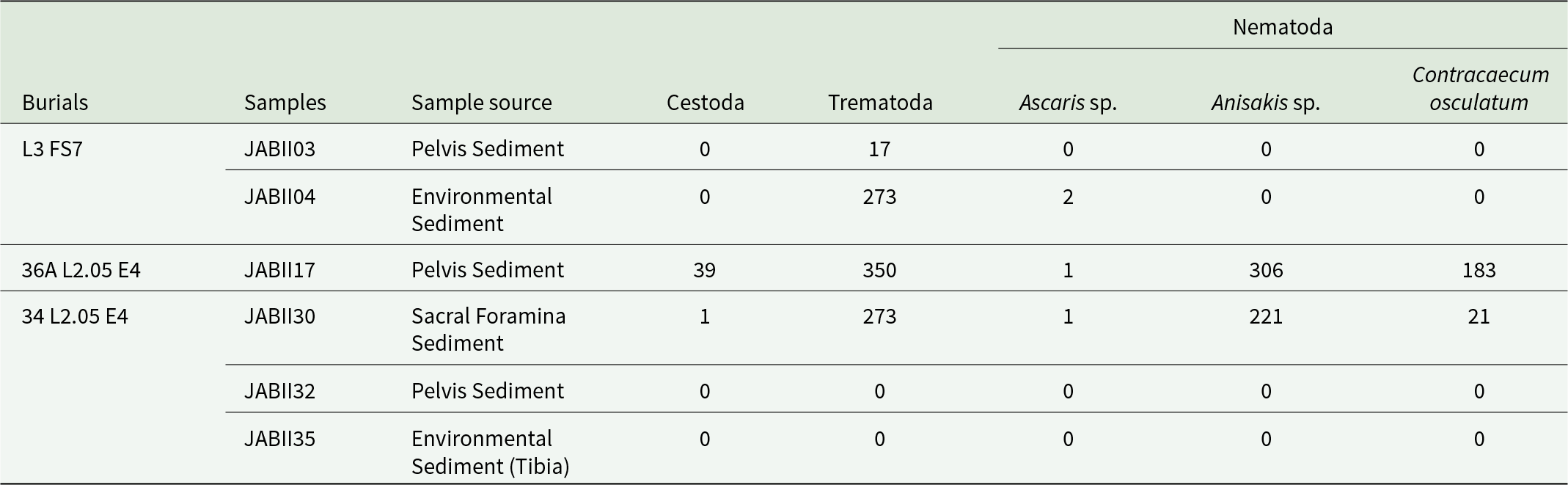
In all JABII samples, a total of 652 219 raw reads were generated, 94 525 reads (∼14.5%) were associated taxonomically by the cox1 gene (see the Supplementary Material). The count of cox1 gene reads for sample was 3724 (JABII03), 3670 (JABII04), 45 920 (JABII17), 40 482 (JABII30), 668 (JABII32) and 51 reads (JABII35) (see the Supplementary Material). The large decrease in the proportion of cox1 gene dataset is justified by the strategy of amplicon pool sequencing using different molecular targets for the same samples, as the objective of other LPIP projects. The most reads were mapped against Streptophyta species (Plantae), with 94.525 sequences (∼97%) (see Supplementary Material). A total of 764 human reads (∼0.8%) were recovered, and a total of 1688 reads of helminths (∼1.7%), where 735 reads of Nematoda phyla (∼43.5%), 34 reads (∼2%) of Cestoda and 913 reads of Trematoda (∼54%). Other low-proportion taxa were identified, such as Fungi (∼0.09%), Bacteria (∼0.06%), Cnidaria (∼0.005%), Haptophyta (green algae) (∼0.004%) and Arthropoda (∼0.001%). Two of the six samples (JABII32 and JABII35) exhibited a low quantity of raw reads, and no helminth reads were recovered from them, with most of the reads mapped against Homo sapiens (see Tables 1–2 and the Supplementary Material).
The Paleogenomic dataset analysis results with read counts of helminth taxa identified in JABII samples

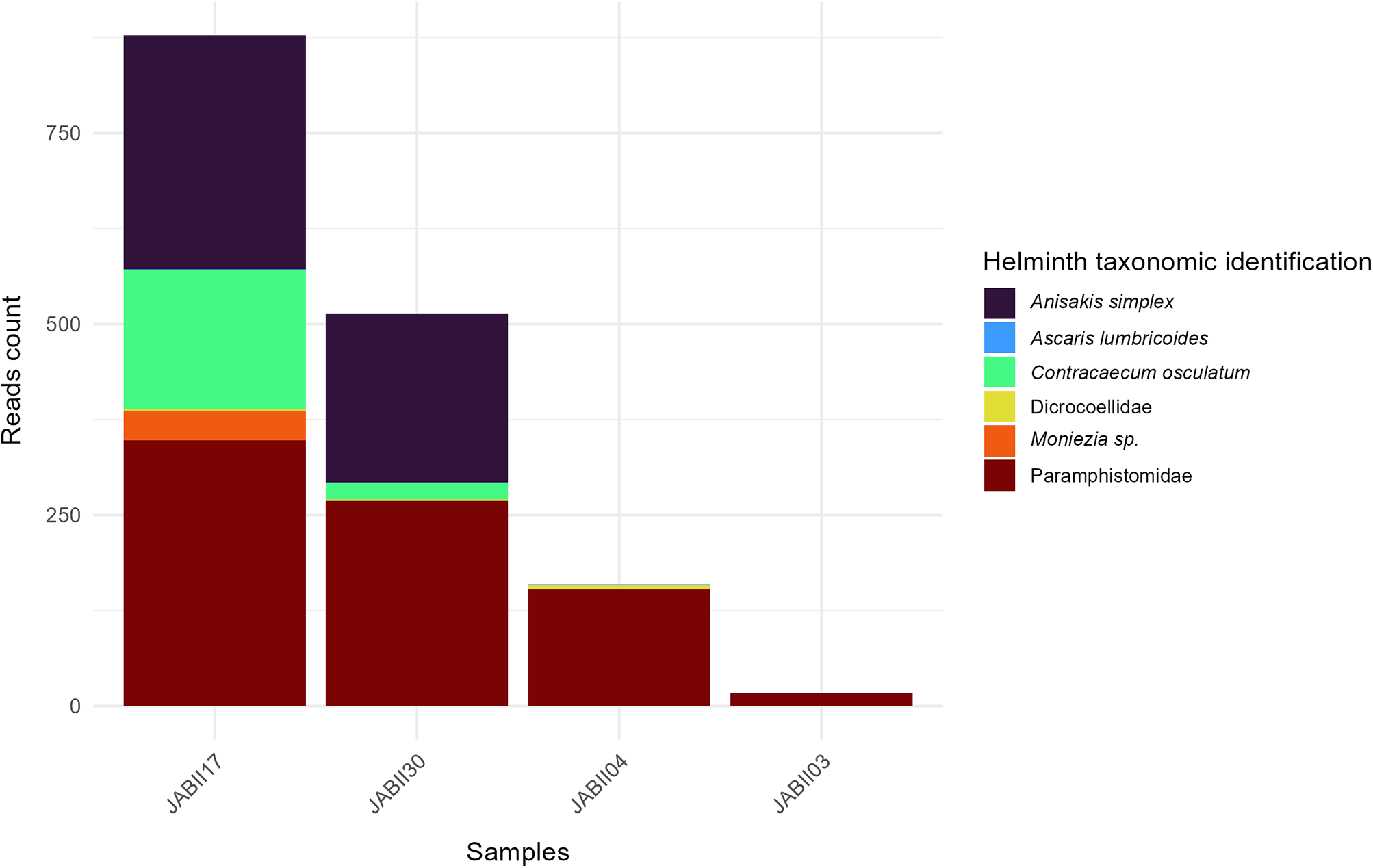
Regarding the Nematoda findings, samples JABII17 and JABII30 show more Nematoda reads, and consequently more species (Figure 2). Two species stand out in terms of read count: Anisakis simplex (Rudolphi, 1809) and Contracaecum osculatum (Rudolphi, 1802) (Figure 2a and Table 2). Despite the relatively low coverage of A. simplex and C. osculatum alignment (∼34×), we observed a high frequency of reads identified as these species, with consistent values of identity (>97%) and E value averages of 7.05 × 10−6, and 3.55 × 10−9, respectively (see Tables 1–2 and the Supplementary Material). Few sequences of Ascaris lumbricoides Linnaeus, 1758 (Nomenclature used in this study as Ascaris sp.) were recovered in JABII04, JABII17, and JABII30 samples (Table 2). However, all these reads were mapped with high identity (>99%) with alignment coverage of 153 (JABII04 and JABII30), and E-value equal to 1.31 × 10−5.
Barplot showing helminths diversity of JABII samples.

Most reads (56%) were mapped to Trematoda and Cestoda classes (see Figure 2, Table 2 and the Supplementary Material). In the Trematoda class, the only order present was Plagiorchiida, with most reads spread across two families, Paramphistomidae and Dicrocoeliidae, in samples JABII04, JABII17, JABII30 and less so in JABII03. From the family Paramphistomidae, the genus Calicophoron sp. was the only one identified, a known cosmopolitan parasite of ruminants, with 100% identity and relatively low cover (∼30×). Regarding Dicrocoeliidae, no species identification was provided, and only a few reads were from JABII04, JABII17 and JABII30 samples. A total of 97.5% of reads of Cestoda were from Anoplocephalidae, indicating the genus Moniezia sp. only in sample JAB17, with 100% identity and 28× cover (see the Supplementary Material).
JABII17 sample harboured the most JABII sequences, and consequently, showed a rich variety of organisms, including marine-associated taxa (see the Supplementary Material). Reads mapped to Chlorophyta and Rhodophyta phyla. The species Phaeocystis globosa (Scherffel, 1899), with 7 unique reads of high coverage (273×), and Hypnea spp., with 91% of identity. Other marine invertebrate organisms were identified, such as Desmophyllum pertusum (Linnaeus, 1758), a cnidarian representative with high coverage (∼200×) and 430 associated reads as Phoronis spp.
Discussion
The huge recent advances in genomics based on HTS have profoundly increased the volume of data recovered from ancient remains (Spyrou et al., Reference Spyrou, Bos, Herbig and Krause2019), making Paleogenomics the ideal approach to study parasite vestiges from sambaquis, due to the drastic taphonomic conditions of these archaeological sites. In this study, we applied palaeogenomics analyses to expand our knowledge about helminth infection in the sambaqui JABII people. The dataset results indicate the Nematoda, Trematoda, and Cestoda parasites in the 3 individuals analysed, confirming Ascaris sp. infection in JABII, and showing new parasite records.
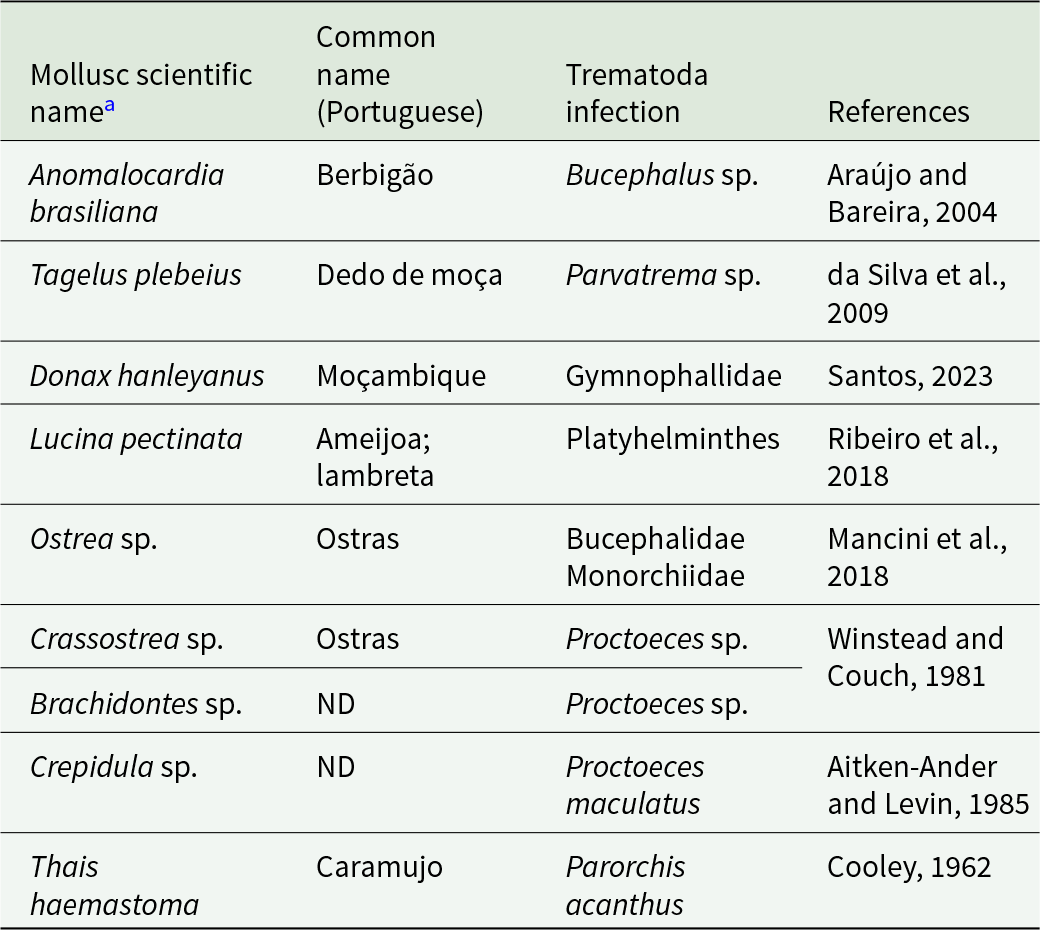
The presence of class Trematoda in JABII is congruent with the lake-marine complex environment and characteristics of a sambaqui archaeological site. It comprises two subclasses: Aspidogastrea, which parasitize freshwater and marine molluscs, and cartilaginous or bony fishes and turtles, and Digenea, with a life cycle of one invertebrate intermediate host, usually a mollusc, and a vertebrate definitive host (Toledo and Fried, Reference Toledo and Fried2017). Especially Digeneans, the largest group, have medical and veterinary relevance, with over 100 species reported infecting humans. The review of literature about the mollusc identified in JABII revealed that 9/14 taxa were found infected by trematodes (Table 3). Paramphistomidae transmission includes livestock and wildlife ruminants as definitive hosts and freshwater snails as intermediate hosts, but the species present in Brazil, especially from wild ruminants, are not completely clarified (Lopes et al., Reference Lopes, López-Hernández and Pinto2024). Following the catalogue of trematode parasites from South America (Fernandes et al., Reference Fernandes, Justo, Cardenas and Cohen2015), Paramphistomum spp. have been reported in several species of wild deer from Brazil, including Mazama spp., the brocket deer reported by Klokler (Reference Klokler2008) as part of the terrestrial fauna of JABII. These results should be considered with caution, since the finding in JABII individuals probably represents an accidental passage of the parasite due to the consumption of bushmeat. In the same way, the evidence of Dicrocoeliidae in two JABII individuals should be considered. Although the infection has been reported in humans, there is no paleoparasitological record in the New World, and the parasite in human samples is often a sign of pseudo-parasitosis due to the ingestion of animal liver infected (Le Bailly and Bouchet, Reference Le Bailly and Bouchet2010), possibly the wild deer ruminant, as part of the diet. Despite the key food of animal source consumed at JABII was fish, the mammals and birds were also hunted for offerings, and the materials were brought to the site. Bird and mammal offerings may have been cooked, but no clear evidence of this activity has been identified (Klokler, Reference Klokler2008). The findings of trematode false parasitism are consistent with the JABII sambaqui faunal assemblages.
Mollusc taxa identified at Jabuticabeira II and reported with trematode infections

a Common mollusc in the JABII archaeological site from Klokler et al., 2010.
ND: No Data.
Regarding Cestoda results, only Moniezia sp. (Anoplocephalidae), an herbivorous parasite, was found in one individual (sample JAB17). There are paleoparasitological records of this cestode in coprolites from South American camelids (Tietze et al., Reference Tietze, Urquiza and Beltrame2021; Ramirez et al., Reference Ramirez, Herrera‐Soto, González Andreu, Santana‐Sagredo, Uribe Rodríguez and Nores2023) and in coprolites consistent with a human or a canid source, both from the Holocene (Fugassa et al., Reference Fugassa, Denegri, Sardella, Araújo, Guichón, Martinez, Civalero and Aschero2006). The cestode findings in camelid populations during the pre-Hispanic period likely reflect long-standing host–parasite associations in Andean environments. The genus Moniezia is nowadays the most frequent tapeworm in cattle, causing significant economic losses, and the infection in wild camelids was interpreted to be due to domestic ruminants sharing the same foraging spaces (Tietze et al., Reference Tietze, Urquiza and Beltrame2021). The Moniezia sp. findings in native artiodactyls before European contact, and consequently the cattle introduction to America, constitute a relevant shift in the enzootic scenario. Regarding Moniezia sp. in the canine or human coprolite, coprophagia and/or herbivorous intake were suggested by the authors (Fugassa et al., Reference Fugassa, Denegri, Sardella, Araújo, Guichón, Martinez, Civalero and Aschero2006). Since it is not a human parasite, the only explanation for Moniezia sp. in the context of the JABII site is pseudo-parasitosis. In the present day, Moniezia spp. was found in Pecari tajacu (Linnaeus, 1758), collared peccary, from Brazil (Sampaio et al., Reference Sampaio, Sianto, Chame, Saldanha and Brener2023), which was indicated as part of the terrestrial habitat fauna of JABII (Klokler, Reference Klokler2008). In general, the genus was found parasitizing a diversity of wild ruminants and has also been documented in primates (Phetla et al., Reference Phetla, Chaisi and Malatji2024). Tayassu spp. (Pecari tajacu), Mazama sp. (the brocket deer) and monkeys (Allouata spp. and Cebus sp.) were reportedly cited as part of the JABII faunal assemblages, and their use as food or pets was suggested (Klokler, Reference Klokler2008). Cestodes, like the trematode findings, are consistent with the environmental context of JABII.
In a previous JABII study, we demonstrated Ascaris sp. infection, based on nad1 gene fragment detection in a pelvic region sediment (JAB03) from the individual excavated at L3 FS7 burial (Iñiguez et al., Reference Iñiguez, Brito, Guedes and Chaves2022). We discussed that this isolated case of ascariasis probably denotes an underestimation of the true rate of soil-transmitted helminthiasis, and a higher diversity of parasites could be infecting the JABII people. We pondered that only a high parasite burden in this individual and/or a well-established transmission through the intra-community could explain the positive record. In this study, Ascaris lumbricoides cox1 reads were recovered from samples of the pelvic (JABII17) and sacral foramina (JABII30) sediments of two additional individuals, 36A L2.05 E4 and 34 L2.05 E4, respectively. In time, we adopted the Ascaris sp. nomenclature, since molecular taxonomic studies revealed a lack of distinction between A. lumbricoides and A. suum Goeze, 1782, to represent a single species or genetic complex infecting humans capable of interbreeding (Iñiguez et al., Reference Iñiguez, Leles, Jaeger, Carvalho-Costa, Araújo and Group2012; Leles et al., Reference Leles, Gardner, Reinhard, Iñiguez and Araujo2012; Easton et al., Reference Easton, Gao, Lawton, Bennuru, Khan, Dahlstrom, Oliveira, Kepha, Porcella, Webster, Anderson, Grigg, Davis, Wang and Nutman2020).
In the individual L3FS7, the abdominal sample (JABII03) tested positive for Trematoda but negative for Ascaris sp., whereas the environmental sample (JABII04) tested positive for both Trematoda and Ascaris sp. In this case, the presence of Trematoda sequences in the environmental sample likely originated from an intestinal infection affecting the L3FS7 individual. The presence of Ascaris sp. sequences in the environmental sample could be explained in the same way, as we had identified the geohelminth in a previous study (Iniguez et al., Reference Iñiguez, Brito, Guedes and Chaves2022). Even though Ascaris sp. sequences were absent from the pelvic sample in the present study, their presence in the environmental sample is understandable, given the soil phase of the life cycle. Given the presence of a geohelminth, it is possible to infer that the parasite infection was established in the JABII population. This interpretation is supported by the identification of Ascaris sp. sequences in two individuals associated with JABII17 and JABII30 samples. The inclusion of environmental context samples in analyses contributes to understanding the surrounding environment and its relationship with the burial context.
On the other hand, in the burial L2.05 E4, the environmental sample showed no helminths, whereas the pelvic and sacral foramina samples tested positive. It is also relevant to note that sediment collection for sacral samples is performed directly from the sacral foramina, which are not freely exposed to the surrounding soil, thereby reducing environmental input.
Previously, a structure suggestive of Ascaris sp. eggs was found in sediment JABII35 associated with the tibia of the 34 L2.05 E4 individual (Iñiguez et al., Reference Iñiguez, Brito, Guedes and Chaves2022). This sediment was initially proposed to be used as a negative control of infection, as sediment close to inferior members is supposed to be out of the gastrointestinal infection site. However, since the individual was buried in a hyperflexed/crouched position, it could indicate true parasite infection, with the tibia bone in contact with the abdominal region. We decided at the time not to consider the suggestive ascarid egg as a conclusive paleoparasitological positive result, because no parasite evidence was observed in abdominal sediments, which is now confirmed in this work (JABII30). It is important to mention that no reads from Ascaris sp. were retrieved from the environmental sample JABII 35 (34 L2.05 E4 individual), but from the environmental sample JABII04, associated with long bones of a young adult, with only 15% of bones preserved (L3 FS7 individual). A limitation of our studies was the use of soil environmental sediments from individuals buried in a hyperflexed position or in poor preservation as control samples, as previously noted (Iñiguez et al., Reference Iñiguez, Brito, Guedes and Chaves2022). Unfortunately, it was not possible to collect additional reliable control samples of infection from JABII site, and these sediment samples should be considered as environmentally informative, which is comprehensible in the case of ascariasis, as a soil-transmitted helminthiasis. The environmental sediment sample JABII04 also showed some trematode reads from Dicrocoeliidae and Paramphistomidae, findings discussed above, possibly due to vestiges of freshwater molluscs or the free-living phases of parasites.
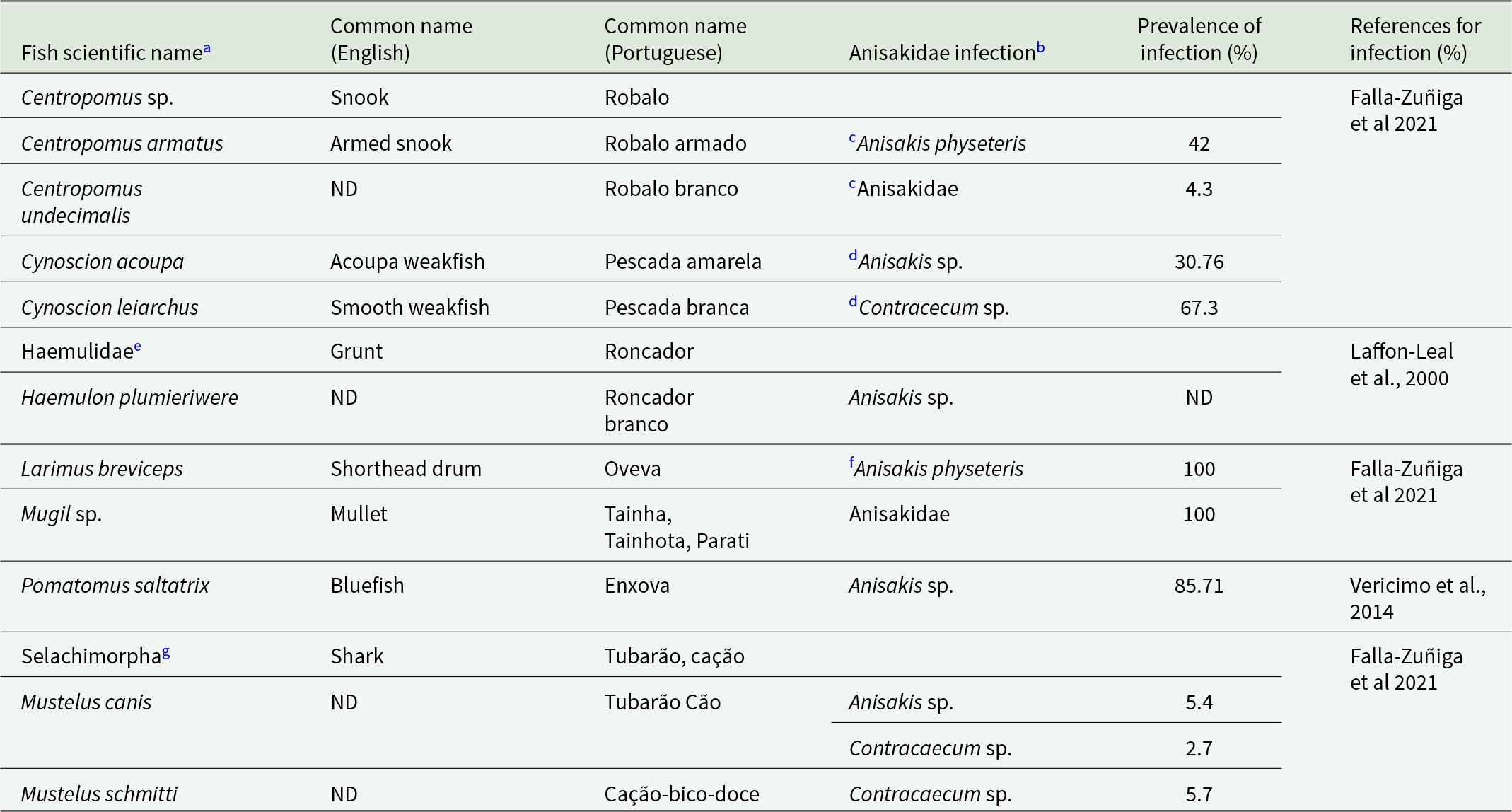
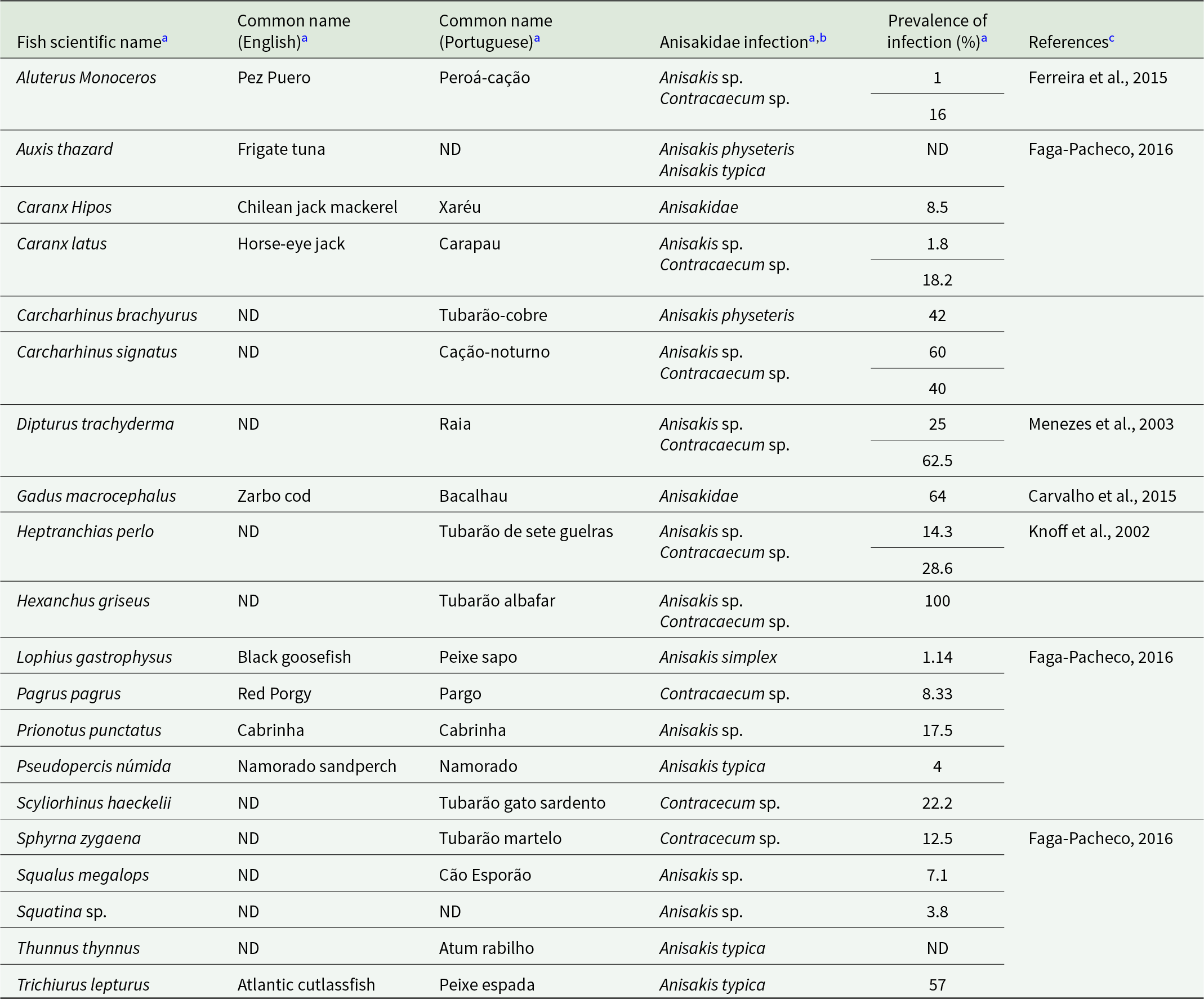
Anisakidae infection by A. simplex and C. osculatum was identified in two JABII individuals, 36A L2.05 E4 and 34 L2.05 E4 (Table 2). Anisakis simplex and Contracaecum osculatum are common parasites from marine crustaceans, cephalopods, and fish as intermediary hosts, marine mammals as definitive hosts, and accidentally infecting humans by consuming mainly fish. The primary food consumed at feasts at JABII was fish (Klokler, Reference Klokler2008), which led to the mass capture of fish for the banquet and the gathering of large specimens for deposition with the dead. Marine fish and cephalopods were significant in the sambaqui diet. We found that 10 common fish taxa at the JABII site have been documented as infected by A. simplex, C. osculatum, and/or Anisakidae, with prevalence of infection from 2.7 to 100% (Table 4). Most of the worldwide Anisakis species described have been recognized on the Brazilian coast (Di Azevedo et al., Reference Di Azevedo, Carvalho and Iñiguez2017; Di Azevedo and Iniguez, Reference Di Azevedo and Iñiguez2018). In addition, 20 fish taxa identified on the South Coast of Brazil were reported with Anisakidae, harbouring from 2.7 to 100% rate of infection (Table 5). These data strongly support the richest panorama of potential sources of Anisakidae infection of JABII people.
Common fish taxa recovered from Jabuticabeira II archaeological site and reported with Anisakidae infection on the Atlantic coast

a Common fish in the JABII archaeological site from Klokler, Reference Klokler2008. Fish with no Anisakidae infection reported were omitted.
b The taxa considered were Anisakis spp., Contracaecum sp., and Anisakidae as cited. Other genera from Anisakidae were omitted.
c The data on infection is relative to the species Centropomus armatus and Centropomus undecimalis.
d The data on infection is relative to the genus Cynoscion.
e The fish taxon recovered from JABII is Haemulidae.
f The data on infection is relative to the species Larimus argenteus.
g Invalid taxon, the valid name is Euselachii, following ITIS (Integrated Taxonomic Information System).
ND: No Data.
Fish species identified on the South Coast of Brazil and reported with Anisakidae infection

a Data from Falla-Zuñiga et al., Reference Falla-Zuñiga, Castellanos-Garzón, Salazar and Pustovrh2021.
b The taxa considered were Anisakis spp., Contracaecum sp., and Anisakidae as cited. Other genera from Anisakidae were omitted.
c Data on fish taxa reported on the South Coast of Brazil.
ND: No Data.
Marine mammals, on the other hand, are notably rare at the JABII archaeological site, with only a single whale bone fragment recovered from a sedimentary profile (Klokler, Reference Klokler2008). This suggests that these animals were not actively hunted and were likely exploited only opportunistically, for example, through natural strandings. Despite their nutritional value, whales appear to have played a limited role in the diet or symbolic practices of local sambaqui groups. A similar pattern is observed for dolphins, although species such as the tucuxi dolphin, Sotalia guianensis (van Bénéden, 1864), and the bottlenose dolphin, Tursiops truncatus (Montagu, 1821), are the most common cetaceans in this coastal region (Daura-Jorge et al., Reference Daura-Jorge, Wedekin and Simões-Lopes2004). Even though records are infrequent, the presence of dolphins and whales provides additional supporting evidence for Anisakidae parasites in the JABII context.
The symptomatic manifestations of anisakiasis in the human body can be essentially divided into four clinical forms: gastric, intestinal, ectopic and allergic anisakiasis (Hochberg and Hamer, Reference Hochberg and Hamer2010; Nonković et al., Reference Nonković, Tešić, Šimat, Karabuva, Medić and Hrabar2025). In addition to these forms, there are asymptomatic forms in which the presence of parasite larvae does not affect the health of the host (Cong and Elsheikha, Reference Cong and Elsheikha2021). In the gastric form, or gastric anisakiasis, the parasite larvae penetrate the gastrointestinal mucosa, causing abdominal pain, nausea and fever (Hochberg and Hamer, Reference Hochberg and Hamer2010). In intestinal anisakiasis, there is abdominal pain, diarrhoea and intestinal obstruction by mucosal oedema (Hochberg and Hamer, Reference Hochberg and Hamer2010). The ectopic form occurs when the parasite larvae migrate to other visceral organs, such as the spleen, ovaries and liver. The allergic form involves specific Anisakis spp. proteins, allergenics and antibodies (Moneo et al., Reference Moneo, Carballeda-Sangiao and González-Muñoz2017; Nonković et al., Reference Nonković, Tešić, Šimat, Karabuva, Medić and Hrabar2025). Allergic form symptoms are rhinoconjunctivitis, asthma, anaphylaxis and dermatitis (Moneo et al., Reference Moneo, Carballeda-Sangiao and González-Muñoz2017).
In the context of the JABII population, the molecular detection of Anisakidae provides evidence of parasite–host interaction but does not allow direct inference regarding clinical manifestation. Importantly, we contextualize the presence of Anisakidae in the JABII population, which is already known to have experienced multiple parasitic infections. Previous studies have documented paleopathological lesions consistent with treponematoses (Filippini, Reference Filippini2012; Filippini et al., Reference Filippini, Pezo-Franco and Eggers2019) and intestinal infection by Ascaris sp. (Iniguez et al., Reference Iñiguez, Hoon, Bianucci, Hoon and Bianucci2021), including multi-parasitism reported in the present study. In addition, paleopathological signals, such as cribra orbitalia and porotic hyperostosis, commonly interpreted as markers of chronic iron-deficiency anaemia and often associated with prolonged infectious stress, have been described at JABII (Filippini et al., Reference Filippini, Pezo-Franco and Eggers2019; Di Giusto, Reference Di Giusto2023). Further evidence of systemic physiological stress includes periostitis affecting both subadults and adults, inferred as indicative of recurrent or endemic infections within the population (Storto et al., Reference Storto, Eggers and Lahr1999; Okumura and Eggers, Reference Okumura and Eggers2005). Despite Okumura and Eggers (Reference Okumura and Eggers2005) observing a low frequency of dental caries at JABII, it is associated with specific pathophysiological conditions affecting the oral cavity, likely reflecting dietary patterns and microbial dynamics distinct from those observed in agricultural populations. The L3 FS7 individual analysed in this study has previously described as having a fragile skeletal condition and evidence of pathological alterations. Although the remains are poorly preserved, earlier paleopathological analyses reported lesions compatible with chronic infectious processes, particularly affecting the right femur (Filippini, Reference Filippini2012, Reference Filippini, Pezo-Franco and Eggers2019). Taken together, these data suggest a context of chronic infectious stress in the JABII population, in which anisakiasis, even if asymptomatic, may have contributed to co-infection dynamics and compromised health. In the case of Ascaris sp., humans are definitive hosts, whereas humans are accidental hosts for Anisakis sp. Therefore, the parasite load becomes relevant for the maintenance of the parasite in the JABII population, as well as the expression of symptoms.
In the study, we performed a Blastn alignment for computational analysis, based on the best hits for identity and cover alignment rates, as a way of obtaining more diversity data in taxa. Alternatively, in the future, the Kraken 2 algorithm (Wood et al., Reference Wood, Lu and Langmead2019), based on the Latest Common Ancestor (LCA) and usually applied in palaeogenomicss, will be evaluated to reduce runtime and confirm positive taxonomic classification (Ravishankar et al., Reference Ravishankar, Perez, Davidson, Roca-Rada, Lan and Llamas2025). Another limitation of the study was the impossibility of applying bioinformatics analysis to aDNA data authentication, by using, for example, the MapDamage2 program (Jónsson et al., Reference Jósson, Ginolhac, Schubert, Johnson and Orlando2013). The software identifies aDNA-specific damage patterns in a dataset obtained by the HTS shotgun method, based on whole DNA alignment against a reference genome, and the HTS data in the present study are from metabarcoding amplicon sequencing. Certainly, the shotgun method will be essential in the further investigations into the paleoecological and health scenarios of JAB II, both to expand the data on the archaeological context and to authenticate aDNA.
In conclusion, the present palaeogenomics study revealed an increase in frequency and diversity of helminth presence in JABII people. We report here, for the first time, Anisakidae fish parasite infection, the confirmation of ascariasis spreading, and pseudo-parasitism by plathelminths, which configured a more complete and richer paleoparasitological panorama of JABII people. The study exposed intestinal parasites that reflect both their close relationship with the marine and estuarine environmental settings and the health challenges faced in daily life. The novel data completely agree with the subsistence of JABII based on a diet of fish and molluscs, their way of life as fisher-gatherers, the crowding and the sedentary lifestyle of long-standing settlements, and the faunal surroundings in the Southern coastal region of Brazil.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182026101863.
Data availability statement
All data used within this study are presented within the main text.
Acknowledgements
We would like to thank Sabine Eggers (Naturhistorisches Mu-seum, Wien, Anthropologische Abteilung, Burgring, Vienna, Austria) for their valuable collaboration, for her always humble availability. We also thank the FIOCRUZ bioinformatic platform (RPT04A/FIOCRUZ) and the Genomics Unit of INCA/RJ for their valuable technical assistance during the study.
Author’s contribution
AMI conceptualized the study. LB and CF completed laboratory analysis. JA and MP completed computational analysis and data interpretation. JA, LB and AMI wrote the initial manuscript. JA, LB, MP, CF and AMI contributed to data interpretation, manuscript preparation, and revision.
Financial support
This project was supported by a grant of fellowships from the Coordination for the Improvement of Higher Education Personnel (CAPES) (http://www.capes.gov.br/) (JA, grant no. 88887.002115/2024-00) and (LB, grant no. 88887.978546/2024-00); National Council for Scientific and Technological Development (CNPq) (http://www.cnpq.br/) (AMI, grant no. 315634/2021‐9) and Carlos Chagas Filho Foundation for Research Support of the State of Rio de Janeiro (FAPERJ)] (http://www.faperj.br/) (AMI, award numbers FAPERJ/CNE E-26/201.069/2021 and E-26/210.131/2023). The founders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript.
Competing interests
None.
Ethical standards
Not applicable.



 Open access
Open access