



Introduction
Echinococcosis is a worldwide zoonotic disease caused by the larval stages of the cestode genus Echinococcus (Rudolphi, Reference Rudolphi1801). In sub-Saharan Africa, cystic echinococcosis (CE) caused by Echinococcus granulosus has been described from South Africa (Ortlepp, Reference Ortlepp1937; Verster, Reference Verster1965), although more comprehensive data are lacking (Aschenborn et al. Reference Aschenborn, Aschenborn, Beytell, Wachter, Melzheimer, Dumendiak, Rüffler, Mackenstedt, Kern, Romig and Wassermann2023). Several different life-cycle patterns occur. The most significant cycle involves humans as accidental or intermediate hosts, together with a variety of livestock species and domestic animals acting as definitive hosts. A sylvatic life cycle that uses a variety of wild ungulate species as herbivorous intermediate hosts also exists and was previously discovered in Africa (Romig and Wassermann, Reference Romig and Wassermann2024). In the past, wild canids, such as hunting dogs or African wild dogs (Lycaon pictus), jackals (Canis mesomelas and C. aureus), hyaenas (Crocuta crocuta) and sometimes the domestic dog (Canis lupus familiaris) were found to act as definitive hosts (Ortlepp, Reference Ortlepp1937; Verster, Reference Verster1965; Verster and Collins, Reference Verster and Collins1966; Young, Reference Young1975; Hüttner and Romig, Reference Hüttner and Romig2009). A recent study conducted in Namibia identified all 5 species of E. granulosus sensu lato in the country (Aschenborn et al., Reference Aschenborn, Aschenborn, Beytell, Wachter, Melzheimer, Dumendiak, Rüffler, Mackenstedt, Kern, Romig and Wassermann2023).
A sylvatic cycle involving the African lion and several intermediate host species, including the warthog (Phacochoerus africanus), bush pig (Potamochoerus larvatus), Burchell’s zebra (Equus burchelli), wildebeest (Connochaetes taurinus), buffalo (Syncerus caffer) and various antelope species, has also been reported from several African countries, these include South Africa, Namibia, Botswana, Kenya, Uganda, Tanzania and Zambia (Verster and Collins, Reference Verster and Collins1966; Thompson et al., Reference Thompson, Lymbery and Constantine1995; Hüttner et al., Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008; Halajian et al., Reference Halajian, Luus-powell, Roux, Nakao, Sasaki and Lavikainen2017; Zaffarano et al., Reference Zaffarano, De Klerk-lorist, Junker, Mitchell, Bhoora, Poglayen and Govender2021).
The E. granulosus sensu lato complex comprises several recognized species, including E. granulosus sensu stricto (sheep-associated), Echinococcus ortleppi, Echinococcus equinus, Echinococcus canadensis and Echinococcus felidis (Thompson et al., Reference Thompson, Lymbery and Constantine1995; World Health Organization and World Organisation for Animal Health, 2001, 2008; Thompson and McManus, Reference Thompson and McManus2002; Xiao et al., Reference Xiao, Qiu, Nakao, Li, Yang, Chen, Schantz, Craig and Ito2005; Hüttner et al., Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008; Nakao et al., Reference Nakao, Yanagida, Konyaev, Lavikainen, Odnokurtsev, Zaikov and Ito2013; Vuitton et al., Reference Vuitton, McManus, Rogan, Romig, Gottstein, Naidich, Tuxun, Wen and Menezes da Silva2020).
The molecular identification of Echinococcus species in South Africa has been limited to cysts obtained from human patients (including E. canadensis G6/7, E. granulosus sensu stricto and E. ortleppi), a cow (E. ortleppi), a white rhinoceros (Ceratotherium simum) (E. equinus), 3 hippopotamuses (E. felidis) and worms from 1 lion (E. felidis) (Hüttner et al., Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008; Mogoye et al., Reference Mogoye, Menezes, Wong, Stacey, von Delft, Wahlers and Frean2013; Halajian et al., Reference Halajian, Luus-powell, Roux, Nakao, Sasaki and Lavikainen2017; Zaffarano et al., Reference Zaffarano, De Klerk-lorist, Junker, Mitchell, Bhoora, Poglayen and Govender2021; Aschenborn et al., Reference Aschenborn, Aschenborn, Beytell, Wachter, Melzheimer, Dumendiak, Rüffler, Mackenstedt, Kern, Romig and Wassermann2023). Despite the extensive history of wildlife research, including studies on lions, a significant knowledge gap remains regarding lions as hosts of Echinococcus.
In sub-Saharan Africa, E. granulosus sensu stricto, E. ortleppi (cattle-associated), E. equinus (horse-associated), E. canadensis (notably camel-associated) and E. felidis have been reported, with E. felidis maintained predominantly in a wildlife (sylvatic) cycle involving lions and a range of wild ungulate intermediate hosts. Among these, E. ortleppi and E. equinus are well-established as distinct species, supported by clear genetic differentiation and host specificity (Thompson and McManus, Reference Thompson and McManus2002).
Although canids are regarded as the primary definitive hosts of Echinococcus species, E. felidis was first reported in lions in South Africa in 1937 (Ortlepp, Reference Ortlepp1937), demonstrating that felids can also serve as definitive hosts. The species was identified based on the distinctive rugosity of the rostellar hooks and its presence in a felid host (Hüttner and Romig, Reference Hüttner and Romig2009). This classification has been supported by various authors (Verster, Reference Verster1965; Hüttner et al., Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008; Thompson, Reference Thompson2008). Genetic characterization of lion-derived Echinococcus isolates is crucial for confirming their taxonomic relationship with other species and genotypes of Echinococcus (Thompson, Reference Thompson2008). A study by Hüttner et al. (Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008) compared taeniid eggs from a Ugandan lion’s feces with a preserved sample from a South African lion collected in the 1960s (Verster, Reference Verster1965), revealing genetic similarity, though the DNA was distinct from E. granulosus based on partial genome analysis. Mitochondrial genome-based phylogenetic analysis has since confirmed a sister relationship between E. felidis and E. granulosus sensu stricto (Nakao et al., Reference Nakao, Yanagida, Konyaev, Lavikainen, Odnokurtsev, Zaikov and Ito2013). A recent study provided the first molecular confirmation of E. felidis in Tanzania, identifying the African lion and the leopard as definitive hosts within the Serengeti ecosystem (Ndossi et al., Reference Ndossi, Mjingo, Zebedayo, Choe, Park, Dongmin, Eom and Bia2025).
Echinococcus equinus, formerly known as the horse strain of E. granulosus, is recognized as a distinct species (Saarma et al., Reference Saarma, Jõgisalu, Moks, Varcasia, Lavikainen, Oksanen, Simsek, Andresiuk, Denegri, Gonzalez, Ferrer, Garate, Rinaldi and Maravilla2009). It was first described morphologically in Great Britain (Williams and Sweatman, Reference Williams and Sweatman1963) and appears to use equids (such as horses, donkeys, mules and zebras) exclusively as intermediate hosts (Young, Reference Young1975; Thompson and McManus, Reference Thompson and McManus2002; Jenkins et al., Reference Jenkins, Romig and Thompson2005; Thompson, Reference Thompson2008; Saarma et al., Reference Saarma, Jõgisalu, Moks, Varcasia, Lavikainen, Oksanen, Simsek, Andresiuk, Denegri, Gonzalez, Ferrer, Garate, Rinaldi and Maravilla2009). Canids, including dogs and foxes, can serve as definitive hosts for E. equinus (Howkins et al., Reference Howkins, Heath and Smyth1965). Additionally, recent findings have reported infections of E. equinus in lions, Black-backed jackals and Burchell’s zebras in Namibia’s Etosha National Park (Wassermann et al., Reference Wassermann, Aschenborn, Aschenborn and Mackenstedt2015). The presence of E. equinus was confirmed for the first time in white rhinoceros (C. simum) in Kruger National Park (KNP) in 2021 (Zaffarano et al., Reference Zaffarano, De Klerk-lorist, Junker, Mitchell, Bhoora, Poglayen and Govender2021). The life-cycles of E. felidis and E. equinus appear to be entirely or predominantly sylvatic (Aschenborn et al., Reference Aschenborn, Aschenborn, Beytell, Wachter, Melzheimer, Dumendiak, Rüffler, Mackenstedt, Kern, Romig and Wassermann2023).
While extensive research has been conducted on wildlife dynamics and various aspects of Panthera leo ecology, our understanding of lions as potential hosts of Echinococcus spp. remains significantly limited. Despite a longstanding history of studies focused on the biology, behaviour and conservation of lions, the implications of these large carnivores as reservoir hosts for this genus of parasitic tapeworms have not been thoroughly explored. This gap is likely attributable, in part, to the logistical and ethical challenges associated with sampling apex predators in natural ecosystems. Consequently, a gap in the literature persists regarding the role of lions in the transmission and epidemiology of Echinococcus, warranting further investigation to elucidate the interactions between these felids and the parasite’s life cycle.
Materials and methods
Study area
KNP, situated in the northeast of Mpumalanga and Limpopo provinces of South Africa, encompasses nearly 2 million hectares of undisturbed wilderness. This region presents a remarkable mosaic of diverse ecosystems and rich wildlife. As one of Africa’s largest game reserves, it serves as a vital sanctuary for the iconic ‘Big Five’ species – lions, elephants, buffalo, rhinoceroses and leopards – while also supporting a diverse array of avian, reptilian and floral species. The KNP is home to 147 mammal species (Zaffarano et al., Reference Zaffarano, De Klerk-lorist, Junker, Mitchell, Bhoora, Poglayen and Govender2021).
Sample origin
Based on the availability of post-mortem examinations, complete intestinal tracts from 4 lions were collected in 2012 from KNP. During necropsy, the small intestines were carefully removed and ligated at both ends to prevent leakage of contents. All procedures were conducted under appropriate biosafety conditions, including the use of personal protective equipment such as gloves, protective clothing and face shields, to minimize the risk of zoonotic transmission. The intestinal contents were handled with caution and transported in sealed, clearly labelled containers in accordance with established biosafety and biosecurity guidelines for parasitic infections. Each intestine was then dissected, the mucosal layer was scraped using a glass microscope slide and the scrapings were suspended in phosphate-buffered saline at pH 7.4 and incubated at 37 °C for 30 min. Following incubation, the samples were macroscopically and microscopically examined for the presence of Echinococcus parasites. Positive samples were subsequently divided into 3 groups and preserved in different solutions: (i) 95% ethanol, (ii) 20% ethanol and (iii) cold 2.5% glutaraldehyde (GA) solution. All samples were stored at 4 °C.
Sample processing for morphological examination
Adult parasites were stained and cleared by using Lacto Carmine – lacto-propiono-dimethylsulfoxide carmine) (Bonga, Reference Bonga1976), Van Cleave’s haematoxylin (method obtained from Prof R. M. Overstreet, 2003, Gulf Coast Research Laboratory, University of Southern Mississippi) and polyvinyl lactophenol clearing (Gray and Weiss, Reference Gray and Weiss1950). Samples were mounted, viewed and photographed after staining or clearing using a Nikon Eclipse Ni light microscope (100× magnification) and a Nikon Stereo SMZ745T microscope (7.5× zoom magnification) for a full morphological description. Samples preserved in 2.5% GA solution were cleaned and routinely processed for scanning electron microscope (SEM).
PCR analysis
For each lion sample that was evaluated microscopically positive for Echinococcus worms, 20 worms previously stored in 95% ethanol were randomly selected for further analysis. The worms were placed into a 1.5 mL Eppendorf tube and used for DNA extractions using the QiAamp®DNA Blood Mini kit (Qiagen, Netherlands) according to the manufacturer’s protocols. A fragment of the mitochondrial cytochrome c oxidase subunit I (cox1) gene was amplified using the published JB3/JB4.5 primer set (Bowles et al., Reference Bowles, Blair and McManus1992). PCR amplifications were performed in a final reaction volume of 50 µL containing 5 µL of template DNA, 1X reaction buffer, 200 mm of deoxynucleotide Triphosphates (dNTP), 0.4 µm of each primer and 0.5 units of Diamond Taq DNA polymerase (Roche). Thermal cycling comprised an initial denaturation at 95 °C for 5 min, followed by 35 cycles of 50 s at 94 °C, 50 s at 55 °C and 50 s at 72 °C, and a final extension step at 72 °C for 10 min. The amplicons were visualized by 1.5% agarose gel electrophoresis. Amplifications of the correct size (441 bp) were submitted for purification and sequencing at Inqaba Biotechnical Industries (Pty) Ltd (Pretoria, South Africa). For initial identification, each DNA sequence was used in a BLASTn search of the GenBank nucleotide database at the National Centre for Biotechnology Information (NCBI).
Results
In this study, the intestines of 4 lions, comprising 1 male and 3 females with estimated ages ranging from 3 to 10 years, were examined. These lions originated from the Greater KNP area. A significant number of Echinococcus worms (n ≥ 500) were detected in the small intestines of 2 lions (female ± 5 years of age; female ± 6 years of age), only few were present in 1 lion (n ≤ 20) (male ± 10 years of age) and the fourth lion’s (male ± 3 years of age) remains were too decomposed to provide viable data.
Light microscopy and scanning electron microscopy study
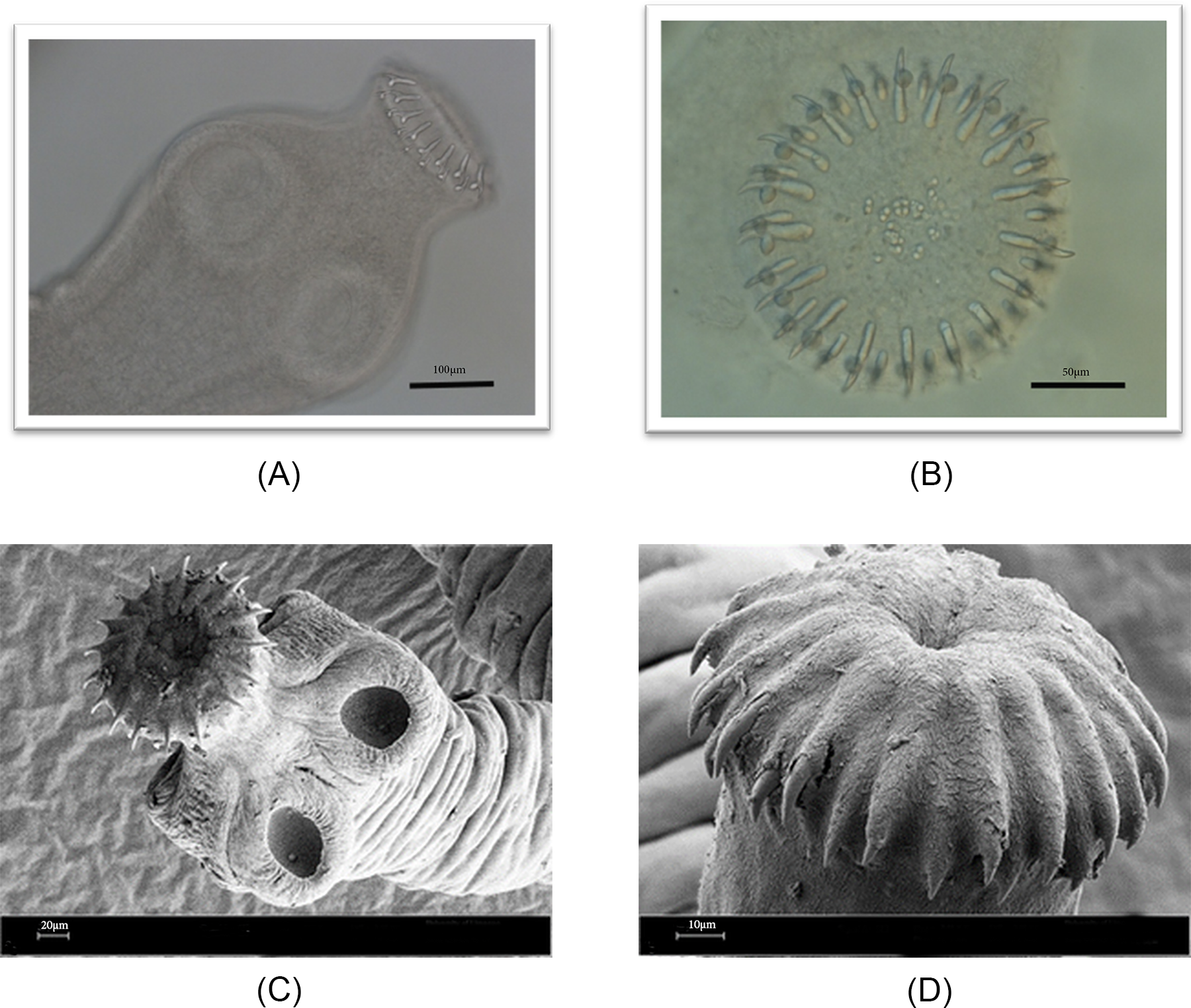
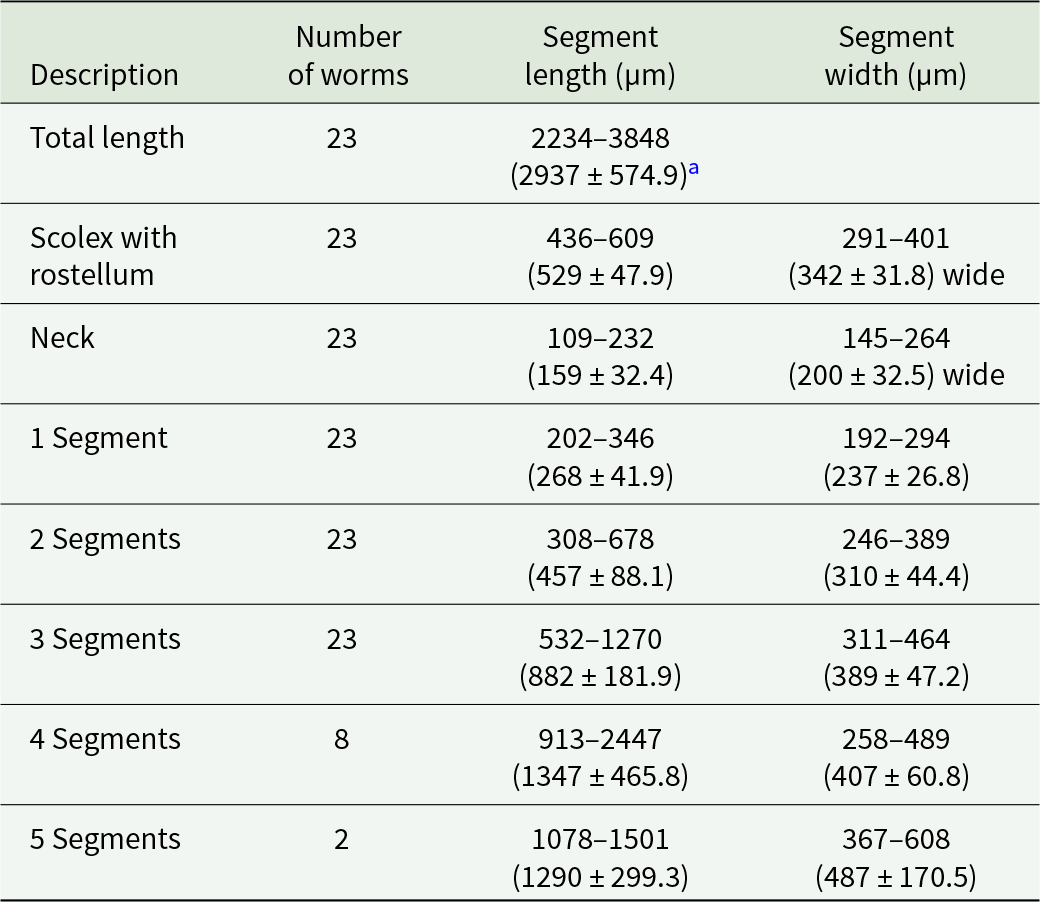
The adult form of Echinococcus spp from 2 with abundant worms was collected manually after post-mortem examination of the intestines, where they appeared as small, white worms. A total of 23 worms were investigated and measured as described in Table 1. The worms were characterized by a typical scolex at the anterior end, featuring 4 well-developed suckers and 2 distinct rows of rostellar hooks – 1 large and 1 small (Figure 1). The number of hooks on the rostellum ranged from 34 to 38. The hook arrangement was highly uniform, with 2 alternating rows of large and small hooks present on the rostellum (Figure 2).
(A) Light micrograph and (B) SEM micrograph of E. equinus recovered from P. leo.

Figure 1 Long description
This image shows an adult E. equinus specimen recovered from the intestine of a lion (P. leo) and examined using light microscopy. The tapeworm consists of a distinct scolex at the anterior end, followed by a short neck region and a series of proglottid segments. The scolex bears characteristic attachment structures, including visible suckers. The body exhibits the typical morphology of Echinococcus species, with a small number of progressively enlarging segments extending posteriorly. The terminal segment is noticeably larger than the immature anterior segments, reflecting maturation of the proglottids. The specimen is stained, allowing differentiation of internal structures and segment boundaries. A scale bar representing 600 µm is provided for size reference.
Light and SEM micrographs of E. equinus showing (A) the scolex with suckers and hooks under light microscopy, (B) the rostellum with a double row of hooks, (C) the scolex with protruded rostellum, extended hooks and suckers under SEM and (D) the rostellum bearing hooks.

Figure 2 Long description
This light micrograph shows the anterior attachment organ (scolex) of an adult E. equinus. Two of the 4 muscular suckers are visible as circular structures on the lateral surfaces of the scolex. At the apex, the armed rostellum protrudes slightly and bears a series of hooks arranged around its margin. The hooks appear as curved, refractile structures projecting from the rostellum and are used by the parasite to anchor to the intestinal mucosa of the definitive host. The image demonstrates the characteristic morphology of an armed cestode scolex. A scale bar of 100 µm is included.
Measurements of 23 E. equinus adults

Table 1 Long description
Measurements are reported for adult Echinococcus equinus body regions and sequential segments, with sample sizes per row. For 23 worms, total body length spans 2234 to 3848 micrometres, with an average of 2937 and a standard deviation of 574.9. The scolex with rostellum measures 436 to 609 micrometres long and 291 to 401 micrometres wide, while the neck is shorter at 109 to 232 micrometres long and 145 to 264 micrometres wide. Segment length increases with segment number: segment 1 is 202 to 346 micrometres, segment 2 is 308 to 678, and segment 3 is 532 to 1270, each based on 23 worms. Segment widths also rise from segment 1 at 192 to 294 micrometres to segment 3 at 311 to 464 micrometres. Later segments are based on fewer worms and show wider ranges: segment 4 is measured in 8 worms at 913 to 2447 micrometres long and 258 to 489 micrometres wide, and segment 5 is measured in 2 worms at 1078 to 1501 micrometres long and 367 to 608 micrometres wide. Because the number of worms decreases for segments 4 and 5, those values are less representative and should be compared cautiously to earlier segments.
a All measurements are in micrometres as minimum to maximum values, followed by the average and standard deviation in parentheses.
The large hooks measured 43.35–50.81 µm, averaging 47.08 µm, while the small hooks ranged from 27.95 to 35.97 µm, averaging 31.96 µm. The rostellar hooks exhibited a smooth outline, with a pronounced groove along the inner blade surface on both sides (Figure 3A). The hooks had long handles, some of which tilted upward at the tips; this feature was variable, with other hooks exhibiting a straighter profile toward the tips. The guard portion of the hooks had a consistently rounded appearance across both large and small hooks (Figure 3A). Although sharp claw-like hooks attached to the rostellum were mostly observed (Figure 3B), some SEM images also revealed that the hooks could be retracted into a membranous sheath within the rostellum (Figure 3C).
Light and SEM micrographs of E. equinus showing (A) large and small rostellar hooks under light microscopy, (B) a double row of large and small hooks under SEM and (C) a scolex with retracted hooks.

Figure 3 Long description
This light micrograph provides a detailed view of the rostellar armature of E. equinus. The hooks are arranged in a characteristic double row consisting of larger and smaller hooks alternating around the circumference of the rostellum. Individual hooks are visible as curved, blade-like structures with broadened bases and pointed tips directed outward. The larger hooks are more robust and elongated, while the smaller hooks are shorter but maintain a similar shape. The arrangement and size variation of the hooks are important diagnostic features used in the morphological identification of E. equinus. The image highlights the symmetry of the hook arrangement and the distinction between the 2 hook size classes. A scale bar of 50 µm is included.
Scanning electron microscopy of the specimens’ tegument revealed that the outer membrane of the cestode tegument projected toward the host as fine, finger-like structures known as microtriches, which covered the entire surface of the worm, including the suckers. Two morphologically distinct types of microtriches were identified. The first type, located on the scolex, was thinner, more filamentous and contained electron-dense material (Figure 4A and B). The second type, observed on the strobila, was thicker and blade-like (Figure 4C). On the strobila, these microtriches were densely packed and flattened, forming a network that encapsulated the bulbous sensory endings.
SEM micrographs of E. equinus showing the tegumental surface structures: (A, B) sucker regions bearing microtriches and (C) bulbous sensory endings with blade-like microtriches.

Figure 4 Long description
This scanning electron micrograph shows the surface of a sucker from the scolex of E. equinus. The sucker appears as a large, circular depression surrounded by raised muscular margins. The entire attachment surface is densely covered with microscopic hair-like projections known as microtriches, which give the surface a velvety texture. These structures increase the surface area of the tegument and contribute to nutrient absorption and host attachment. The image demonstrates the complex architecture of the sucker and the dense distribution of surface projections associated with attachment to the host intestinal mucosa. Scale bar = 2 µm.
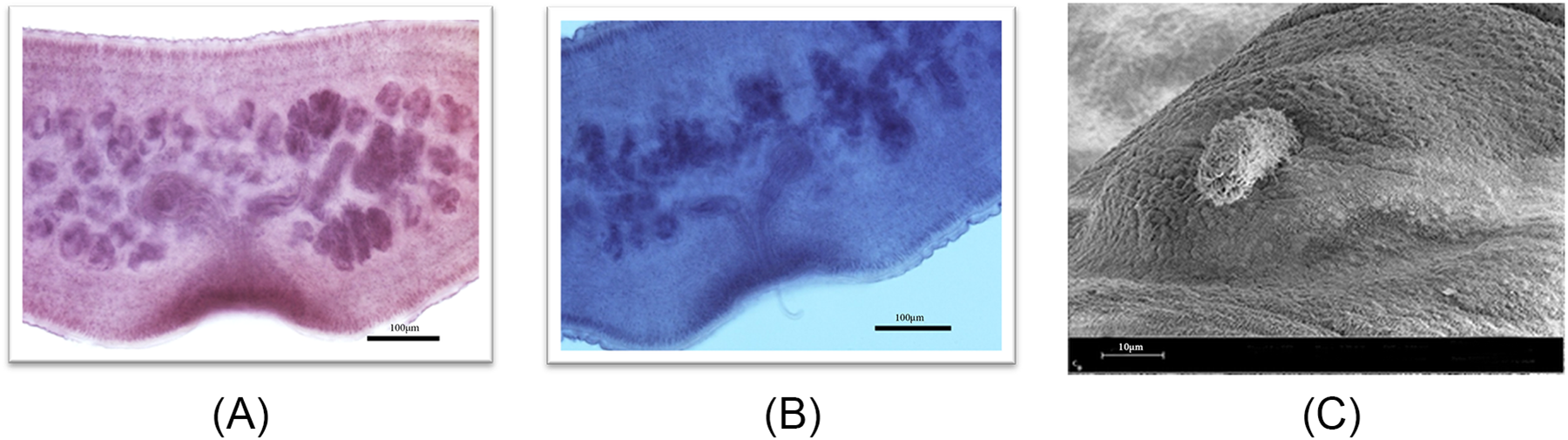
The number of testes observed ranged from 32 to 43, with some positioned posterior to the vitelline gland within the segment. The cirrus sac exhibited a shape that varied from round to slightly oval and was oriented more parallel to the genital pore opening, situated closer to the centre of the segment. The cirrus tube displayed an anterior bend, which was acute in most specimens but less pronounced in others, due to variations in relaxation prior to segment fixation. In mature segments, the genital pore opening was located near the middle of the segment, while in gravid segments, it was more posteriorly positioned. The 2 lobes of the ovary were generally spread horizontally and demonstrated a distinctly irregular shape (Figure 5A). The male copulatory organ, represented by the cirrus, was identified in some light micrographs (Figure 5B) and SEM images (Figure 5C).
(A) Light micrograph of a mature proglottid; (B) mature proglottid with cirrus; and (C) SEM micrograph of the cirrus organ.

Figure 5 Long description
This light micrograph shows a mature proglottid of E. equinus. The segment is rectangular in shape and represents one of the sexually mature reproductive units of the tapeworm. Internal reproductive structures are visible within the proglottid, including components of the male and female reproductive systems characteristic of hermaphroditic cestodes. The segment is wider than long and displays well-defined boundaries separating it from adjacent proglottids. The reproductive organs occupy a substantial portion of the segment, reflecting their role in gamete production and fertilization. The image illustrates the organization of the mature reproductive stage before the development of a gravid uterus. A scale bar of 200 µm is included.
PCR analysis
DNA from 3 samples was successfully amplified with primers JB3/JB4.5, yielding the expected 441 bp fragment of the mitochondrial cox1 gene. A neighbour-joining tree was constructed using 390 bp of the cox1 gene, depicting the relationships of the investigated samples with previously published Echinococcus genotypes (tree not shown). Two of the samples were grouped with the cox1 sequence for isolate G4–horse-dog strain (NCBI accession number AF346403, Le et al., Reference Le, Pearson, Blair, Dai, Zhang and McManus2002). The third sample was grouped with a representative sequence for E. felidis (NCBI accession number EF558356, Hüttner et al., Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008). The 2 samples identified as E. equinus were selected for further microscopic analysis because adult worm samples were available.
Discussion
This study contributes to expanding knowledge of the role of lions as potential definitive hosts of Echinococcus species, particularly E. equinus and E. felidis, in the Greater KNP, South Africa. The presence of these parasites in lions has significant implications for understanding the epidemiology and transmission dynamics of echinococcosis in African wildlife ecosystems, given the limited recent data available (Aschenborn et al., Reference Aschenborn, Aschenborn, Beytell, Wachter, Melzheimer, Dumendiak, Rüffler, Mackenstedt, Kern, Romig and Wassermann2023).
The detection of E. felidis in 1 lion sample is consistent with earlier reports of this parasite in South African lions (Ortlepp, Reference Ortlepp1937; Hüttner et al., Reference Hüttner, Nakao, Wassermann, Siefert, Boomker, Dinkel, Sako, Mackenstedt, Romig and Ito2008). Echinococcus felidis, also known as the ‘lion strain’ of E. granulosus, has been identified based on genetic characteristics. The genetic analysis in this study, which grouped the E. felidis sample with previously published sequences, reinforces the taxonomic distinction of this parasite from E. granulosus sensu stricto. The finding of E. felidis in lions from the Greater KNP highlights the potential role of these predators in maintaining the sylvatic cycle of echinococcosis in southern Africa.
The identification of E. equinus in 2 lion samples aligns with previous reports of this parasite in other wild carnivores, such as black-backed jackals and Burchell’s zebras in Namibia’s Etosha National Park (Wassermann et al., Reference Wassermann, Aschenborn, Aschenborn and Mackenstedt2015). A recent study also reports a first incident of E. equinus in a Southern white rhinoceros from KNP (Zaffarano et al., Reference Zaffarano, De Klerk-lorist, Junker, Mitchell, Bhoora, Poglayen and Govender2021). A comprehensive morphological study was performed on random specimens from the intestines of 2 lions in the KNP, from the same batch prepared for genetic characterization, to determine the status of this species. Similarities between the present E. equinus description and the studies done by Williams and Sweatman (Reference Williams and Sweatman1963) and Kumaratilake et al. (Reference Kumaratilake, Thompson and Eckert1986a) include the total body length, number of segments, length of the large and small hooks, the number of testes and their position, the position of the genial pore, shape and position of the cirrus and form of the ovary and seminal receptacle. The morphological characteristics observed in the current study, including the rostellar hooks and segment structure, are consistent with the descriptions of E. equinus provided by Saarma et al. (Reference Saarma, Jõgisalu, Moks, Varcasia, Lavikainen, Oksanen, Simsek, Andresiuk, Denegri, Gonzalez, Ferrer, Garate, Rinaldi and Maravilla2009) and Thompson and McManus (Reference Thompson and McManus2002). The major difference between the E. equinus specimens compared in this study and the 2 morphological studies by Williams and Sweatman (Reference Williams and Sweatman1963) and Kumaratilake et al. (Reference Kumaratilake, Thompson and Eckert1986a) is that the host in this study is lions, whereas in the other studies it is horses or other equids. The finding of E. equinus in lions suggests that these large carnivores may play a more significant role in the life cycle of this parasite than previously recognized, potentially contributing to its transmission within wildlife populations.
The discovery of both large and small hooks on the rostellum, with the large hooks measuring 47.08 µm on average, is consistent with previous studies and confirms the morphological distinction of E. equinus from other Echinococcus species (Kumaratilake et al., Reference Kumaratilake, Thompson and Eckert1986b). The presence of a well-developed cirrus sac and the number of testes observed further corroborate the identification of E. equinus. The presence of 2 morphologically distinct types of microtriches on the tegument, as revealed by SEM, provides insights into the adaptation to its felid host. Microtriches are specialized tegumental projections that increase the surface area available for nutrient absorption and facilitate attachment and interaction with the host intestinal environment in cestodes (Chervy, Reference Chervy2009). The detailed morphological and SEM analyses provide a comprehensive description of this parasite’s structure, which can be useful for future comparative studies.
The identification of E. equinus and E. felidis in lions underscores the importance of understanding the role of wildlife in the epidemiology of echinococcosis. Lions, as predators, may serve as reservoir hosts for these parasites, facilitating their transmission to other susceptible species, including wild ungulates, potentially domestic animals and humans. Lions (P. leo) are opportunistic yet selective predators whose prey choice reflects both availability and behavioural decision-making during the hunting sequence. Studies from KNP show that medium-sized ungulates dominate lion diets, with species such as zebra, wildebeest and kudu accounting for a substantial proportion of kills, with zebra alone contributing approximately 15–16% of recorded prey items across long-term datasets (Maruping-Mzileni et al., Reference Maruping-Mzileni, Funston and Ferreira2017). Similarly, behavioural studies reveal that lions preferentially prey on species such as plains zebra (E. burchellii), not only because of abundance but also because of higher encounter rates and active selection during hunting. Lions tend to forage in habitats where preferred prey is more likely to occur and are more likely to initiate hunts on these species once encountered. However, prey selection is dynamic and influenced by environmental factors such as rainfall and prey condition; for example, during droughts, lions may shift toward larger or more vulnerable prey such as buffalo. Overall, zebras represent a common and consistent component of lion diets across African savanna systems, reflecting both their ecological availability and their suitability as preferred prey (Hayward et al., Reference Hayward, Hayward, Tambling and Kerley2011).
While extensive research has been conducted on wildlife dynamics and various aspects of P. leo ecology, our understanding of lions as potential hosts of Echinococcus spp. remains significantly limited. This gap is likely attributable, in part, to sampling challenges, including the opportunistic nature of post-mortem access, limited availability of suitable carcasses and restrictions within protected areas. In addition, the zoonotic risk posed by Echinococcus spp. necessitates stringent biosafety procedures, further constraining field and laboratory investigations. Prioritization of livestock and human health over wildlife reservoirs in many African research settings may also contribute to the lack of data. Consequently, a gap in the literature persists regarding the role of lions in the transmission and epidemiology of Echinococcus, warranting further investigation to elucidate the interactions between these felids and the parasite’s life cycle.
Conclusion
This study provides new insights into the role of P. leo as a definitive host of E. equinus and E. felidis in the Greater KNP. However, the relatively small sample size limits the ability to accurately determine the true positivity rate of lions for Echinococcus spp. in this ecosystem. Further studies with larger sample sizes are required to better estimate infection prevalence, although such efforts remain challenging due to the logistical, ethical and biosafety constraints associated with sampling apex predators in natural settings, as previously discussed.
The morphological and genetic analyses confirm the presence of these parasites in lions, with potential implications for wildlife health and the transmission dynamics of echinococcosis in southern Africa. The molecular and morphological findings and data generated in this study and in future studies in the KNP and surrounding conservation areas will contribute to knowledge and a better understanding of Echinococcus spp. (especially E. felidis and E. equinus) in southern Africa as a whole.
Acknowledgements
We gratefully acknowledge the assistance of the field and technical staff of the State Veterinary Office, Kruger National Park, with sample collections. Dr C. Baker and the Electron Microscope Unit at Sefako Makgatho Health Sciences University are acknowledged for technical assistance. During the preparation of this article, the author used ChatGPT (OpenAI, the latest version available in 2026) and Grammarly (the latest version available in 2026) to assist with language editing, improving clarity and readability. ChatGPT was further used to create the graphical abstract.
This article is partially based on the author’s thesis entitled ‘Genetic and morphological characterization of Echinococcus spp. isolated from lions in the Kruger National Park South Africa’ towards the degree of Magister Technologiae: Veterinary Technology in the Department of Biomedical Sciences, Tshwane University of Technology, South Africa, with supervisors C. Boshoff and P. King, received in 2017, it is available here: https://hdl.handle.net/20.500.14519/2385.
Author contributions
SSJ conducted laboratory work, including morphological analyses and data collection. PK supervised morphological analyses. CB conceived the study, oversaw sample collection, genetic analyses and contributed to manuscript preparation. All authors approved the final manuscript.
Financial support
This project was funded by the Department of Biomedical Sciences, TUT.
Competing interests
The authors declare there are no conflicts of interest.
Ethical standards
Samples were obtained from ad hoc post-mortem examinations conducted in accordance with SANPARK wildlife management protocols. No animals were euthanized specifically for this study.




 Open access
Open access