


1. Introduction
The notion of level is of wide application in contemporary philosophy and biology.Footnote 1 Oppenheim and Putnam (Reference Oppenheim, Putnam, Feigl, Scriven and Maxwell1958) suppose the classical locus of the so-called layer-cake model of reality, according to which levels are structured in a stepwise compositional fashion, where theories formulated in terms of higher-level components (e.g., social structures) could potentially be reduced to theories formulated in terms of lower-level components (e.g., atoms or subatomic particles). Oppenheim and Putnam’s model was supposed to account for all the existing sciences, under the assumptions that the order of the sciences and how these could be theoretically reduced to one another reflected the ultimate order of nature. An underlying assumption in their model was that levels generated hierarchies, and these hierarchical ranks were essential for understanding nature.
Even while Oppenheim and Putnam’s model is rejected in contemporary science because of its many incongruences (see Potochnik and McGill Reference Potochnik and McGill2012), the concept of level still plays a key role in today’s philosophy, particularly in its application to biology. In this case, its application is narrower than in Oppenheim and Putnam’s, as we are focusing not on how nature as a whole is structured but rather on how the different biological entities (cells, multicellular organisms, colonial organisms, etc.) are hierarchically structured. In fact, the notion of level is pervasive in several fields of biology. For example, in physiology, it is customary to distinguish systems, organs, tissues, and cells; in ecology, one often distinguishes between ecosystems, regional ecosystems, local ecosystems, avatars, and organisms; in taxonomy, between kingdoms, phyla, classes, orders, families, genus, and species; and so forth. Levels can be established based on several criteria: scale levels (arranged according to scale properties such as size or tempo), causal levels (arranged according to the action of causality), or compositional levels (part–whole relations) (Craver Reference Craver2007, 171). In general, the different biological levels are established in hierarchies of compositional inclusion, where entities at a higher level are supposed to include, as components, only entities from the lower level, in a way that creates the appearance of a nested order (sec. 3).
The reason why we tend to order the biological world into levels partially rests on their heuristic value in understanding the complexity of nature. As Eldredge (Reference Eldredge1985), doing pioneering work on the notion of levels and on biological hierarchies, once put it: “[H]ierarchies actually deal with complexity by teasing it apart; it is as if hierarchies are more honest in their simple recognition that a system is complex than is an approach that seeks unity in characterizing the system in simple terms” (Eldredge Reference Eldredge1985, 11).
Similarly, Wimsatt characterizes levels “as local maxima of regularity and predictability in the phase space of alternative modes of organization of matter” (Reference Wimsatt2007, 209); Brooks et al. (Reference Brooks, DiFrisco and Wimsatt2021) open their Levels of Organization in the Biological Sciences by claiming that “levels demarcate quasi-independent channels of causal interactions and are sites of scale-specific dynamics and causal generalizations” (Brooks et al. Reference Brooks, DiFrisco and Wimsatt2021, 7, emphasis added). All these are ways of claiming that levels are structures of reality that both allow understanding its complexity and, I would claim, reflect the very structure of nature.
In this article, I focus on one of the uses of the notions of hierarchy and levels as they are applied in evolutionary biology, concretely in the debate on levels of selection, where it has been especially useful to tease apart biological complexity. I specifically argue that this view of levels of organization as levels of selection generates a nonnested evolutionary hierarchy, highlighting the necessity of rethinking our metaphysical assumptions on hierarchy construction, at least in evolutionary biology. Secondly, I consider a potential objection to my approach derived from a recent work by Eronen and Ramsey (Reference Eronen and Ramsey2025), where they claim that the notion of “levels” used in the debates on units of selection should be distinguished from the notion of “level” that is used in the debate on levels of organization. I will argue that Eronen and Ramsey (Reference Eronen and Ramsey2022) miss an important distinction between two types of units of selection: interactors and manifestors of adaptation (manifestors, in what follows) (Lloyd Reference Lloyd, Evelyn and Elisabeth1992, Reference Lloyd and Edward2024; Suárez and Lloyd Reference Suárez and Lloyd2023). If we take this distinction into account, then levels of selection correspond to levels of organization because manifestors are the result of organizational processes in their focal level.
2. The levels of evolutionary organization are nonnested
The nested hierarchy of the evolutionary hierarchy is generally accepted in biological research. For example, in his Unfinished Synthesis, Eldredge claims, “Many recent discussions of hierarchy in evolution seem almost to synonymize hierarchy with selection at different levels. A fundamental premise of this book is that the biological entities that participate in the evolutionary process are themselves hierarchically arranged in a nested fashion. Thus, ontology [i.e., how selection operates] virtually forces our evolutionary theory to be structured hierarchically” (Reference Eldredge1985, 104, emphasis added).
Echoing this, Tëmkin and Eldredge (Reference Tëmkin, Eldredge, Serrelli and Gontierr2015, 184) have recently claimed: “Biological evolutionary theory is ontologically committed to the existence of nested hierarchies in nature,” restating Eldredge’s commitment to the nestedness of the evolutionary hierarchy.Footnote 2
The assumption about nestedness is also present in most discussions about evolutionary transitions in individuality (Maynard-Smith and Szathmáry Reference Maynard Smith and Szathmáry1995). For instance, Michod’s (Reference Michod1999) model, according to which a transition entails a fitness transfer from the lower-level entities to the higher-level entities, involves an assumption about nestedness: If the lower level loses its direct visibility to natural selection because its fitness gets transferred to the higher level, then the higher level must contain, as components, all and only the lower-level units. Similar considerations apply to Godfrey-Smith’s (Reference Godfrey-Smith2009) model of Darwinian individuality. The evolution of a Darwinian individual at the higher level entails the de-Darwinization of the lower-level units, which become nested within the newly emerged Darwinian individual.
The assumption is also present in earlier works on units of selection. For instance, Lewontin’s (Reference Lewontin1970) founding article presupposed that the biological world was nested in a hierarchy that included molecules, organelles, cells, gametes, (multicellular) organisms, some groups of organisms, populations, and higher levels. Hull’s (Reference Hull1980) abstract approach to natural selection, distinguishing between interactors and replicators, generated a dual hierarchy of both interactors and replicators, as illustrated in Brandon’s (Reference Brandon and Henry1988) work. In all these cases, nested evolutionary hierarchies were presumed (for a helpful analysis, see Griesemer Reference Griesemer, Cartwright and Martin2005).
Similarly, the assumption is present in most philosophical accounts about levels, including Craver’s (Reference Craver2007) mechanistic model, in which mechanisms are composed of parts that can also be mechanisms, and this generates a nested rank. This equally applies to Wimsatt’s (Reference Wimsatt2007) model: “By level of organization, I mean here compositional levels—hierarchical divisions of stuff … organized by part-whole relations, in which wholes at one level function as parts at the next (and at all higher) levels” (Wimsatt Reference Wimsatt2007, 201, emphasis added).Footnote 3
This general view concerning the nested nature of levels of organization can be summarized by citing a passage from Eronen and Brooks (Reference Eronen, Brooks, Edward and Nodelman2023, sec. 1, emphasis added; see also Potochnik and McGill Reference Potochnik and McGill2012, 121): “This idea that levels of organization of organization form ‘nested’ compositional hierarchies where there are wholes at higher levels and their components at lower levels, and the components themselves can be further decomposed into parts, remains one of the core features of the notion of levels of organization up to this day.”Footnote 4
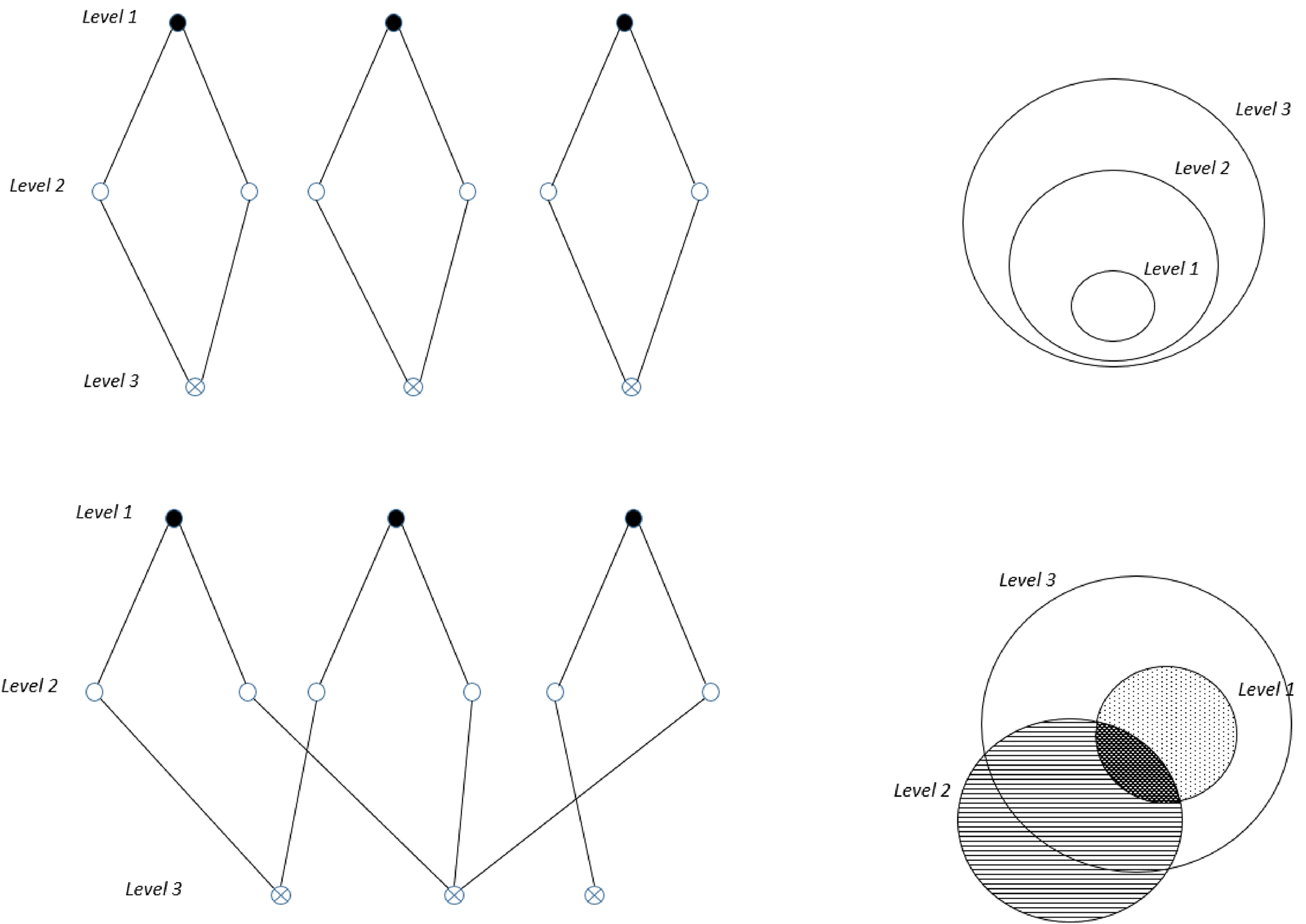
To argue that a hierarchy is nested means that the entities in the hierarchy are related based on inclusiveness. In a nested hierarchy, the level where a phenomenon occurs is the focal level, and entities below (less inclusive) and above (more inclusive) are the lower and higher levels, respectively. Semiformally, nestedness can be defined as follows: Take three levels, Ln, Ln*, and Ln**, and assume that Ln** is higher than Ln*, which is in turn higher than Ln. Then, when one generates a rank of these three levels, for example, a rank of sets and subsets, it is assumed that the rank constitutes a nested hierarchy if all the entities belonging to Ln belong to Ln*, and hence they also belong to Ln**. The nestedness assumption derives from the idea that levels include a relation of composition, and composition is ordered in a nested way. Figure 1 shows two ways of representing nested and nonnested hierarchies.
Upper, Two ways of representing nested hierarchies (graph; Venn diagram). Level 2 would be the focal level. Lower, Two ways of representing nonnested hierarchies (graph; Venn diagram).

Figure 1. Long description
Panel A: Graph representing nested hierarchies. Three diamond-shaped structures are shown, each with three levels labeled Level 1, Level 2, and Level 3. The top node is labeled Level 1, the middle nodes are labeled Level 2, and the bottom nodes are labeled Level 3. Panel B: Venn diagram representing nested hierarchies. Three concentric circles are shown, each labeled Level 1, Level 2, and Level 3, with Level 1 being the innermost circle and Level 3 being the outermost circle. Panel C: Graph representing nonnested hierarchies. Three diamond-shaped structures are shown, each with three levels labeled Level 1, Level 2, and Level 3. The top nodes are labeled Level 1, the middle nodes are labeled Level 2, and the bottom nodes are labeled Level 3, with some connections crossing between different diamonds. Panel D: Venn diagram representing nonnested hierarchies. Three overlapping circles are shown, each labeled Level 1, Level 2, and Level 3, with varying degrees of overlap between the circles.
If the assumption about the nested nature of the evolutionary hierarchy is so pervasive,Footnote 5 then one may think there is something correct in it. Unfortunately, the assumption is simply an ideal that does not correspond to the real structure of levels of evolutionary organization and, hence, the type of ideal that a pragmatist should reject. I have two reasons to assert this.
Firstly, the mechanism that produces levels of evolutionary organization does not require a nested biological hierarchy. To introduce this argument properly, I must first introduce a distinction between two meanings of levels of evolutionary organization or, also, levels of selection (Lloyd Reference Lloyd, Evelyn and Elisabeth1992, Reference Lloyd and Edward2024; Suárez and Lloyd Reference Suárez and Lloyd2023).Footnote 6 The term levels of selection can be ambiguously used to refer to two types of entities: interactors and manifestors. A unit of selection is an interactor when, in effect, it engages (as a cohesive whole) in fitness-affecting interactions with its environment in a way that reproduction is differential. For example, for all we know, and assuming that sexual selection is going on, humans are interactors for traits such as their eye colors because different variants of the trait within the population are expected to lead to differential reproduction of their bearers. On the other hand, a unit of selection is a manifestor when it is the result of selection reputedly acting at its focal level so that one or some of its traits are engineered (or to put it differently, are a product of transtemporal accumulation) to carry out the function(s) they perform. In this sense, manifestors are the result of optimization at their focal level, at least for one trait and often for more than one trait, as occurs with the type of entities Okasha (Reference Okasha2018) calls type 1 agents (a specific type of manifestor). For instance, humans are manifestors for organs such as the eye because the organ has been optimized through the constant action of natural selection to perform its function. Manifestors are thus products of selection, whereas interactors exist because there is a process of selection currently going on at that level.
Given their nature as products of selection, I consider manifestors to be the best candidates to play the role of levels of evolutionary organization. Thus, we must analyze the mechanism that produces manifestors to see whether nestedness is theoretically required to interpret their evolution. Manifestors are generated through the persistent action of selection on a trait (or traits) on a specific focal level. The mechanism often used to analyze how this process occurs is multilevel selection, or kin selection, but note that this approach requires the action of multilevel selection as well, is reducible to the latter, or even is epistemically parasitic on multilevel selection (see Lloyd Reference Lloyd and Edward2024). Although most multilevel selection models presume a nested hierarchy, in which particles are nested within collectives, this requirement is not necessary. As Okasha explains, “The underlying causal mechanism [of evolution by natural selection] does not require nesting; it could work equally well with overlapping groups of lower-level units” (Okasha Reference Okasha2006, 44). That is, the main process triggering evolution by natural selection (multilevel selection) is often theoretically conceived as if it acted on a nested hierarchy of levels of selection. But the presupposition is not needed for the theory to work and be applicable to the evolution of the biological world.
My argument could be contested by arguing that multilevel selection explains evolution acting on/generating interactors, but not generating manifestors, which are the units I claim generate levels of organization. This is correct. But note that multilevel selection can also generate manifestors, and if it describes the way natural selection acts, it must be the force generating manifestors too—whenever these are produced. There is a more serious objection to this argument, though. One could argue that my position is merely theoretical: Theoretically, multilevel selection could act on a nonnested world and even produce different levels of organization in a nonnested fashion. But the biological world is in fact nested: Empirically, different levels of evolutionary organization only arise in a nested fashion.
I, however, claim that this objection is incorrect because there is evidence of natural selection producing manifestors across a nonnested hierarchy, exactly in the nonnested way I think the evolutionary hierarchy is organized. Some holobionts constitute a canonical example of this (Suárez Reference Suárez2020). Holobionts are symbiotic assemblages that result from the association between an animal or plant host plus its symbiotic microbiome. Most holobionts contain parts that reproduce independently from one another; in most circumstances, this could prevent the evolution of a manifestor at the level of the holobiont, as many researchers have argued (Skillings Reference Skillings2016; Douglas and Werren Reference Douglas and Werren2016). Yet some evidence suggests that some holobionts are manifestors of adaptation in a very specific way, as shown for the case of vampire bats (Suárez Reference Suárez2020). In these cases, certain traits of the microbiome would be responsible for generating the nonnestedness of the evolutionary hierarchy. These traits will be such that they will depend on the microbiome for their phenotypic expression because the animal host (bat) cannot integrate them into their genome and transcribe them by itself. Yet the long-term persistence of these specific traits historically/evolutionarily depends on their phenotypic expression within the context of a vampire bat. Otherwise, these traits would likely be lost by the bacteria bearing them. The main prediction would be that these traits, which have also been called hologenomic traits (Suárez and Triviño Reference Suárez and Triviño2020), will engage in higher horizontal gene transfer within the context of the microbiome, will be easily acquired and expressed by several bacterial phyla, and will also be easily lost outside the host environment.
These traits, which constitute the basis for holobionts to be manifestors, are the ones generating a nonnested evolutionary hierarchy. They would entail the existence of a manifestor at the level of the bacterial phyla of the microbiome, at the level of the host, and finally, at the level of the biological composite formed by the host plus these specific microbiome traits. Note that in this hierarchical organization, the levels of manifestors will be nonnested because only parts of a lower level (the hologenomic traits) will belong to the higher level, but the lower level as a whole would not belong to the higher level, despite its necessity for the expression of lower-level traits. Holobionts thus constitute an empirical example of the nonnestedness of the biological hierarchy.
3. Levels of selection and levels of organization
The account I have presented is prone to one objection, though. In a recent article, Eronen and Ramsey (Reference Eronen and Ramsey2025, 503) ask about the notion of “levels” underlying the debate on units of selection. Their main claim is that there are at least two concepts of hierarchy and level that are usually conflated in the debates on units of selection: a hierarchy of levels of organization (e.g., molecules, cells, organisms, populations, species) and a hierarchy of levels of fitness-affecting interactions (Eronen and Ramsey’s preferred deflationary account of levels of selection), where entities are entangled together because they engage in fitness-affecting interactions with one another. Eronen and Ramsey believe that both hierarchies require the existence of particles (lower level) and collectives or groups of a certain sort (higher level) in a nested relationship.
Eronen and Ramsey believe that these two conceptions of “level” must be distinguished for two reasons. First, ontologically, they claim that “although the parts and collectives in levels of organization can engage in fitness-affecting interactions, they do not have to” (Eronen and Ramsey Reference Eronen and Ramsey2022); additionally, they claim that their criterion for defining levels of selection needs to be liberal enough to accommodate many different processes, and in this sense, it will not have a neat correspondence to levels of organization as these are traditionally defined. Second, epistemically, they argue that each notion of level plays a different role in the debates on organization, the former (levels of selection) being important for analyzing how specific examples of multilevel selection work and the latter for asking more general questions (e.g., the role of multilevel selection in the emergence of new levels of organization, or the relationship between both concepts).
I think Eronen and Ramsey (Reference Eronen and Ramsey2025) are pointing to something partially correct concerning levels of selection, but they fail to use the conceptual tools that would allow them to characterize the problem correctly. Particularly, they fail to distinguish between the two meanings of level of selection I introduced earlier: interactor and manifestor. Distinguishing these two concepts illuminates where their argument fails and why at least a specific meaning of level of selection (manifestor) has a neat correspondence with traditional levels of organization.
If by level of selection, Eronen and Ramsey (Reference Eronen and Ramsey2025) mean interactors, then they are correct: Taking fitness-affecting interactions, which is the defining feature of interactors, as the criterion to determine levels of organization is way too liberal. In fact, a common mistake in part of the recent literature on units is to think that because interactors need to form “a cohesive whole,” they must already possess evolved properties for being visible to natural selection (Suárez and Lloyd Reference Suárez and Lloyd2023, 44). But interactors are simply empirically and “statistically” cohesive; that is, they are cohesive enough so that it is possible to tell apart cases when natural selection is acting on them from cases when the effects perceived at their level are merely a cross-product of selection acting at a different level. In other words, it is easy for interactors to appear. They simply require the existence of a trait at their focal level that leads to differential reproduction either to their own level or to a lower level. That’s why species, almost every holobiont, and groups are often interactors. Interactors lead to product-of-selection adaptations, but they do not necessarily lead to the evolution of engineering adaptationsFootnote 7 at their focal level because they do not necessarily reproduce themselves in a way that is consistent enough to generate those traits. In this sense, as Eronen and Ramsey (Reference Eronen and Ramsey2025) correctly claim, interactors cannot equate traditional levels of organization, even though they also form their own biological hierarchy.
Manifestors are also a level of selection and, I claim, the one that most neatly corresponds to traditional levels of organization, at least as these are seen in biology.Footnote 8 This is because manifestors, because of their own origin as the result of a persistent selection process acting at their focal level, exemplify the typical features of levels of organization as these are conceived in biology. Firstly, manifestors correspond to real structures of the world that can be identified through a robust analysis (Wimsatt Reference Wimsatt2007), their existence is established based on part–whole relationships (i.e., their existence is compositional), and their existence at one focal level imposes constraints on their lower-level component parts (Moreno and Mossio Reference Moreno and Mossio2015). For example, because humans are manifestors for the eye organ, the existence of the organ imposes constraints on how cone and rod cells should be disposed and interact. Or when manifestors become type 1 agents, they reduce the internal conflict between their parts, to a point at which they lose their own autonomy. This latter notion of constraint is important because it is the key element of the organizational account of individuality elaborated by Moreno and collaborators. An advantage of the concept of manifestor is that it allows explaining where these constraints come from (i.e., their origins), what norms they impose in their component parts, and how they operate. Additionally, the type of constraints imposed by the manifestor on the lower-level parts is not a necessary characteristic of the interactor, as Eronen and Ramsey (Reference Eronen and Ramsey2025) also notice. However, it seems to be a defining feature of levels of organization because it allows creating a certain sense of “order” at the focal level, which seems a reasonable criterion for referring to a level as a level of “organization.”
Secondly, all the usual lists of levels of organization include units that are classical manifestors. For instance, Eronen and Ramsey (Reference Eronen and Ramsey2025) provide a list that includes molecules, cells, individuals, populations, and species. But all these have been shown to be manifestors, in certain cases (for a summary, see Lloyd Reference Lloyd and Edward2024). Other lists may include colonial organisms (e.g., some species of ants, termites, bees), holobionts, or some other types of groups. My claim is that the elements of these lists are typically included because they have been shown to be, at least in certain circumstances, manifestors as the concept has been defined for the debates on units of selection.
Thirdly, most of the properties that have been used to define biological individuality, which could ultimately be taken as conditions to elaborate a hierarchy of levels of organization, correspond to features that have been selected for at the specific level where there is a manifestor. This has been shown in Clarke’s (Reference Clarke2013) work on the concept of individuality, where she argues that different properties (division of labor, germ–soma separation, immunological recognition, etc.) correspond to either a demarcating or a policing mechanism, preventing selection from a lower level and hence ensuring that the focal level acts as a manifestor. Thus, a hierarchy of manifestors would be ontologically prior or even equivalent to any other hierarchy of levels of organization.Footnote 9
If my arguments about the relationship between the manifestor and the debate on levels of organization are correct, they show, contra Eronen and Ramsey’s approach, that the debates on levels of selection provide an adequate platform to evaluate the notion of level of organization. Hence, my conclusion about the nonnestedness of the biological hierarchy is still supported by the evidence.
4. Conclusion
In this article, I have carved out one of the metaphysical characteristics of the evolutionary hierarchy (as a hierarchy of levels of selection), arguing that it must be a hierarchy of nonnested levels. My argument has been based on two observations: firstly, that theoretically, there are no grounds to assume that the evolutionary hierarchy needs to be nested, as the mechanism used to study it (multilevel selection) does not require such assumption; secondly, that there are some good empirical examples of levels of evolutionary of organization existing at cross-levels, hence empirically supporting the existence of a nonnested hierarchy. Secondly, I have considered a potential objection by Eronen and Ramsey’s (Reference Eronen and Ramsey2025), who claim that the notion of “level” at play in the debates on levels of selection is different from the notion of “level” that appears in levels of organization. I argued that their position is mistaken because it rests on failing to appropriately distinguish between interactors and manifestors of adaptations as two meanings of the term level of selection. Once this distinction is cleared up, it becomes patent that the hierarchy of interactors does not correspond to the hierarchy of levels of organization, whereas the hierarchy of manifestors (which is the one where I grounded my arguments for the nonnestedness of the biological hierarchy) does.
Acknowledgments
This article was presented at the 2024 Biennial Meeting of the Philosophy of Science Association, as part of the symposium “Pragmatist or Merely Pragmatic? Using Pragmatism in Biological Practice.” I thank all my co-panelists, as well as the participants, for their comments. I also thank Gui Sanches de Oliveira for reading and commenting on a previous draft. Finally, I formally thank the Ministry of Science, Innovativeness and Universities, project PID2022-137993NA-I00, “An Exploration of the Philosophy of Lineages in the Light of Contemporary Biology” for financial support.
Funding Statement
Ministry of Science, Innovativeness and Universities, project PID2022-137993NA-I00, “An Exploration of the Philosophy of Lineages in the Light of Contemporary Biology.”
Declarations
None to declare.

 Open access
Open access