



Introduction
Despite recent studies investigating the helminth faunas of small mammals in South Africa (e.g. Froeschke et al., Reference Froeschke, Harf, Sommer and Matthee2010; Lutermann et al., Reference Lutermann, Medger and Junker2014; Julius et al., Reference Julius, Schwan and Chimimba2018; Spickett et al., Reference Spickett, Junker, Froeschke, Haukisalmi and Matthee2020; Raubenheimer et al., Reference Raubenheimer, Matthee, Stekolnikov, Wentzel, Swanepoel and Matthee2025; Schlemmer et al., Reference Schlemmer, Junker, Matthee and Matthee2026), their diversity remains poorly documented. During a recent survey of nematodes parasitizing small mammals in the Western Cape Province, South Africa (Schlemmer et al., Reference Schlemmer, Junker, Matthee and Matthee2026), several specimens from the stomach of a single female of Micaelamys namaquensis (A. Smith, 1834) (Muridae) from the Nama-Karoo biome could be allocated to the spirocercid genus Gastronodus Singh, 1934, based on the presence of a buccal capsule armed with 6 teeth and the presence of 3 pairs of pedunculate postcloacal papillae (Junker et al., Reference Junker, Lane, McRee, Foggin, van Dyk and Mutafchiev2013). The monotypic genus Gastronodus was erected for Gastronodus strasseni Singh, 1934 from Suncus murinus (Linnaeus, 1766) (= Crocidura coerulea Linnaeus, 1766) (Soricidae) in India (Singh, Reference Singh1934). Originally published as ‘Castronodus strassenii’, Singh (Reference Singh1948) corrected the misprint to ‘Gastronodus strasseni’ and amended and complemented the brief original description of the species. Based mainly on the presence of a buccal capsule armed with 6 strong teeth, Chabaud (Reference Chabaud1959) synonymized 3 genera, Gastronodus Singh, 1934, Skrjabinocercina Matschulsky, 1952 and Petrowospirura Matschulsky, 1952, with Cylicospirura Vevers, 1922. Subsequently, however, he recognized the following subgenera within this genus: Cylicospirura, Gastronodus and Skrjabinocercina (Chabaud, Reference Chabaud, Anderson, Chabaud and Willmott1975). Junker et al. (Reference Junker, Lane, McRee, Foggin, van Dyk and Mutafchiev2013) re-instated these as full genera, with species of Cylicospirura parasitizing mainly felids, canids, hyaenids and dasyurids in all realms except the Antarctic; Skrjabinocercina being monotypic and parasitizing rodents in Russia; and Gastronodus being monotypic (G. strasseni) and reported from S. murinus in India and Thailand.
While the newly sampled specimens from Micaelamys correspond to the genus Gastronodus as defined by Junker et al. (Reference Junker, Lane, McRee, Foggin, van Dyk and Mutafchiev2013), the present specimens differ from G. strasseni in several characters and are described below as a new species.
Molecular data on members of the Spirocercidae are scant with most papers addressing Spirocerca spp., likely because of the high pathogenicity associated with spirocercosis in dogs and other canines (Traversa et al., Reference Traversa, Costanzo, Iorio, Aroch and Lavy2007; Ferguson et al., Reference Ferguson, Woodberry, Gillin, Jackson, Sanders, Madigan, Bildfell and Kent2011; Poon et al., Reference Poon, Tam, Lau, Cheng, Yuen, Schuster and Woo2017; Gomez-Puerta et al., Reference Gomez-Puerta, Carrasco, Lopez-Urbina and Gonzalez2018; Rojas et al., Reference Rojas, Sanchis-Monsonís, Alić, Hodžić, Otranto, Yasur-Landau, Martínez-Carrasco and Baneth2018a,Reference Rojas, Sanchis-Monsonís, Alić, Hodžić, Otranto, Yasur-Landau, Martínez-Carrasco and Banethb; Chen et al., Reference Chen, Zeng, Gao, Zhang, Li and Li2023; Bisterfeld et al., Reference Bisterfeld, Raulf, Springer, Lang, Lierz, Strube and Siebert2024; Ge et al., Reference Ge, Zhang, Meng, Xu, Zhao, Lin, Li and Fu2025). We generated the first molecular data for the genus Gastronodus and, based on mitochondrial cytochrome oxidase 1 (cox1) sequences, examined its preliminary phylogenetic affinities with other members of the Spirocercidae.
Materials and methods
Sample collection and morphological examination
Helminths were fixed and stored in 70% ethanol and cleared in lactophenol for morphological examination. The body length of 6 males was measured while mounted in water, prior to removing their midsection for molecular characterization; the anterior and posterior parts were subsequently studied in lactophenol. Apical views were cut using a razor blade. Specimens were studied under a compound microscope (Olympus BX51) equipped with a drawing tube and digital camera (Olympus DP72). Photos and measurements were taken with the aid of digital imaging software (Olympus cellSens Dimension, version 1.4.1). Specimens used for SEM were dehydrated through a graded ethanol series, immersed in hexamethyldisilazane for 20 min, air-dried, coated with gold in an Emitech K500X sputter coater and examined using a Tescan LYRA 3 XMU FEG/SEMxFIB microscope at an accelerating voltage of 10 kV. All measurements are given in micrometres unless otherwise indicated; the range is followed by the mean in parentheses. Specimens were deposited in the National Collection of Animal Helminths, ARC-Onderstepoort Veterinary Institute, South Africa (NCAH) and in the Helminthological Collection of the Institute of Biodiversity and Ecosystem Research, Bulgarian Academy of Sciences (IBER-BAS).
DNA extraction, polymerase chain reaction (PCR) amplification and sequencing
Genomic DNA was extracted from 2 male specimens using the Qiagen DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) according to the manufacturer’s guidelines. The PCR followed standard cycling procedures, and the PCR reactions included negative controls. Each individual was amplified using a 25 μL total volume containing 3 μL DNA, 8.5 μL distilled water, 12.5 μL Taq DNA Polymerase Master Mix RED (Amplicon, Denmark), and 0.5 μL (1 μM) of each of the respective primers. The CO1intF forward- and CO1intR reverse primers by Casiraghi et al. (Reference Casiraghi, Anderson, Bandi, Bazzocchi and Genchi2001) were used to amplify the cox1 barcoding region. Five microliters of each PCR product, including the negative control, were visualized on a 1% agarose gel stained with ethidium bromide. Sanger sequencing was conducted on the ABI3730xl at the Central Analytical Facility of Stellenbosch University.
Phylogenetic analysis
The Basic Local Alignment Search Tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was used to authenticate the sequences and to retrieve reference sequences of closely related species from GenBank. The newly generated nucleotide sequences were edited and aligned with the corresponding reference sequences using Geneious Prime 2025.1.3 (https://www.geneious.com). The GTR + I + G was selected as the best-fit model of nucleotide substitution by the Akaike information criterion and was used to specify the priors for the Bayesian analyses conducted in MrBayes version 3.2.6 (Ronquist et al., Reference Ronquist, Teslenko, van der Mark, Ayres, Darling, Höhna, Larget, Liu, Suchard and Huelsenbeck2012). A total of 5000 000 generations were used, with 2 independent runs of 4 Markov chains in parallel and trees and parameters were sampled every 1000 generations. The first 25% of samples were discarded as burn-in. Convergence was assessed using the default within-chain diagnostics generated by MrBayes and Tracer version 1.4.3 was used to visualize and edit the trees (Rambaut, Reference Rambaut2016).
Results
Order: Spirurida Chitwood, 1933
Superfamily: Spiruroidea Oerley, 1885
Family: Spirocercidae Chitwood and Wehr, 1932
Genus: Gastronodus Singh, 1934
Gastronodus karooensis Junker, Mutafchiev, Schlemmer, C. Matthee and S. Matthee n. sp
Type host: Micaelamys namaquensis (A. Smith, 1834) (Rodentia: Muridae); Namaqua rock mouse.
Type locality: A farm ca. 59 km north of Merweville, Western Cape Province, South Africa (28.ii.2024; 21.6675, −32.2778).
Site in host: Free in lumen of stomach.
Intensity and prevalence of infection: A single female of 20 hosts examined harboured 19 adult specimens.
Type material: NCAH/2026/001 (holotype male), NCAH/2026/002 (paratypes: 2 males, 3 females, all intact), NCAH/2026/003 (paratype: anterior and posterior extremities of 1 male), NCAH/2026/004 (paratype: anterior and posterior extremities of 1 male – voucher for GenBank Accession Number PZ139158). IBER-BAS N001.471 (paratypes: 1 male, 1 female, both intact; 1 female of which anterior extremity was used for SEM), IBER-BAS N001.472 (paratype: anterior and posterior extremities of 1 male), IBER-BAS N001.473 (paratype: anterior and posterior extremities of 1 male), IBER-BAS N001.474 (anterior and posterior extremities of 1 male), IBER-BAS N001.475 (paratypes mounted on SEM stubs: anterior and posterior extremities of 1 male – voucher for GenBank Accession Number PZ139157; anterior extremity of 1 female).
ZooBank registration: urn:lsid:zoobank.org:act:8FC704EF-F4A8-492D-9726-9F787DFCE3FE.
Etymology: The new species is named after the geographic region it was collected in.
Morphological description (Figs 1–3; Tables 1 and 2)
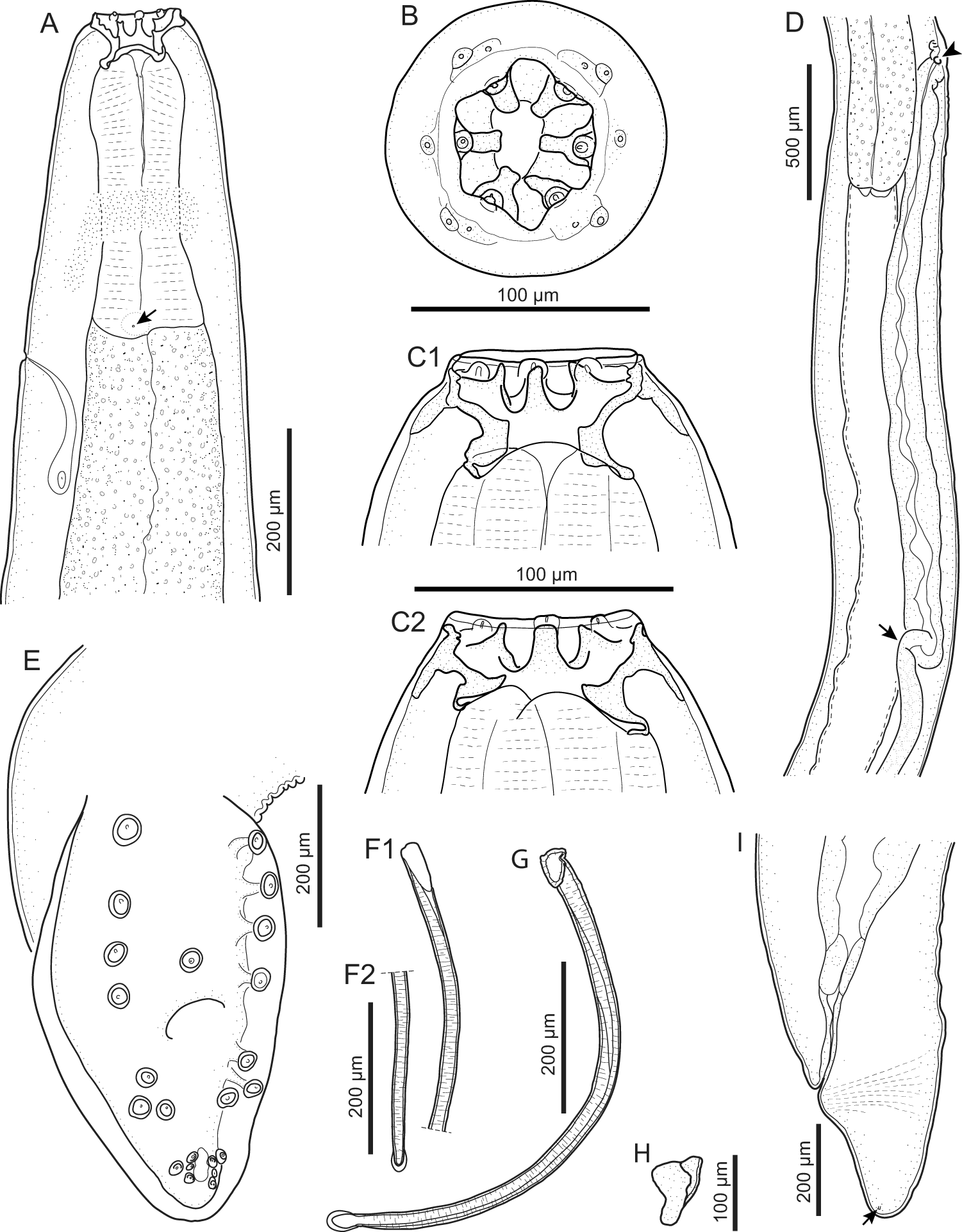
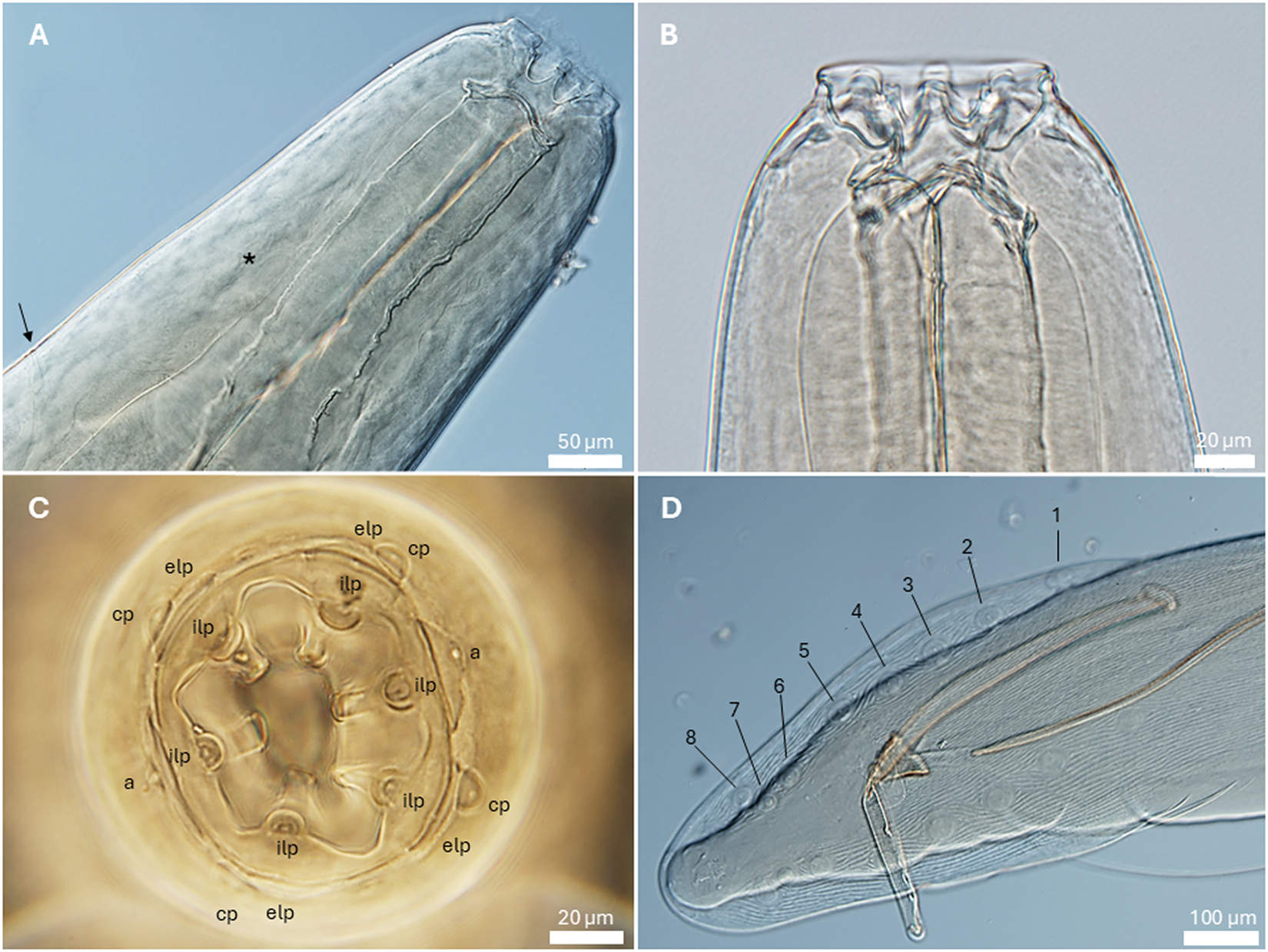
General. Medium-sized spirurid. Body cylindrical, anterior extremity bent dorsally, tapering at both ends; maximum body width at oesophago-intestinal junction. Cuticle with delicate transverse striations, spaced approximately 2 µm apart. Mouth opening hexagonal, elongated in dorsoventral direction, with rounded edges (Figures 1B and 3B). Head papillae composed of 6 prominent internal labial papillae in lateral and submedian positions, one at the base of each tooth, close to mouth opening, and an outer circle of 4 submedian external labial papillae associated with 4 prominent cephalic papillae (Figures 1B and 2C). Amphids posterior to lateral teeth. Buccal capsule short, heavily sclerotized, armed with 6 large teeth, 2 lateral and 4 submedian, arising from a short, concave base (Figure 2B); each tooth usually with 2 slightly pointed apical cusps with shallow dent between them (Figures 1B and C and 3B). Accessory teeth absent. Oesophagus distinctly divided into anterior muscular and posterior glandular part (Figures 1A and 2A). Muscular oesophagus short and stout, with pronounced dilatation close to its junction with buccal capsule; glandular oesophagus distinctly increasing in width just posterior to junction with muscular oesophagus, and subsequently more or less cylindrical. Nerve-ring surrounding muscular oesophagus in posterior half (Figures 1A and 2A); deirids symmetrical, minute, slightly bifurcated (Figure 3C), at level of junction of muscular and glandular oesophagus; excretory pore slightly posterior to deirids (Figures 1A and 2A).
Gastronodus karooensis n. sp. (A) Anterior end, female, lateral view, note the deirid (arrow); (B) Apical view, male; (C) Anterior extremity, male, lateral view, illustrating 2 different physiological states of the buccal capsule (C1 and C2); (D) Oesophago-intestinal junction and terminal part of female genital system, lateral view, note the vulva (arrowhead) and the end of the ovejector (arrow); (E) Posterior end, male, subventral view, longitudinal cuticular ridges not indicated; (F) Left spicule, proximal (F1) and distal end (F2); (G) Right spicule, lateral view; (H) Gubernaculum, ventrolateral view; (I) Posterior end, female, lateral view, note the phasmid (arrow).

Gastronodus karooensis n. sp. (A) Anterior end, female, lateral view, note the nerve ring (asterisk) and excretory pore (arrow); (B) Anterior extremity, male, lateral view; (C) Male, apical view, a – amphid, cp – cephalic papilla, elp – external labial papilla, ilp – internal labial papilla; (D) Posterior end, male, ventral view, caudal papillae numbered according to Chabaud and Petter (Reference Chabaud and Petter1961).

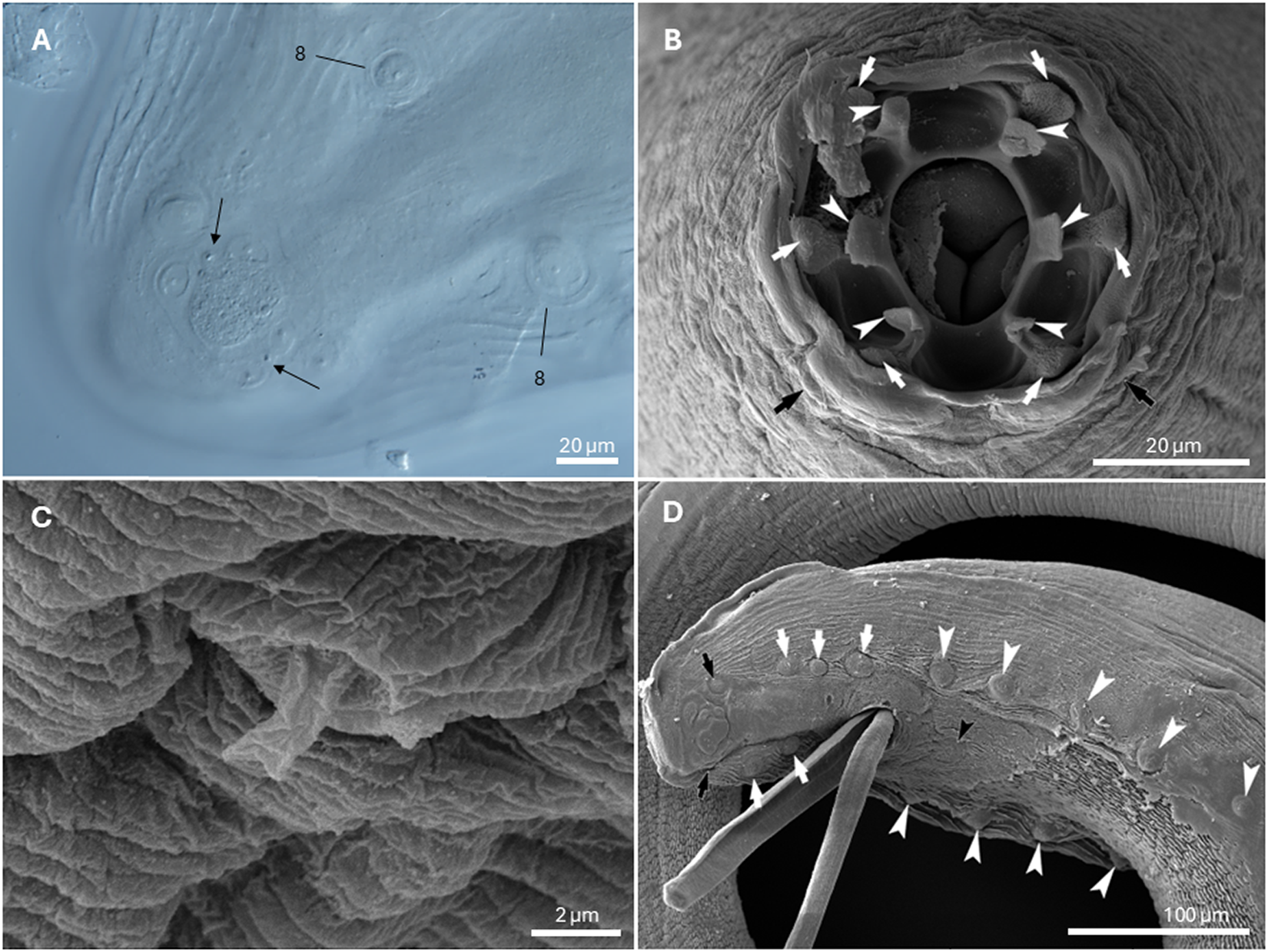
Gastronodus karooensis n. sp. (A) Posterior extremity, male, ventral view, note last pair of postcloacal papillae (8), arrows indicating phasmids; (B) Apical view, female, SEM, note teeth (white arrowheads), internal labial papillae (white arrows) and cephalic papillae (black arrows); (C) Deirid, female, SEM; (D) Male posterior end, SEM, sublateral view, note single ventral papilla (black arrow head), precloacal pairs of pedunculate papillae (white arrow heads), postcloacal pairs of pedunculate papillae (white arrows) and subterminal pair of pedunculate ventrolateral papillae (black arrows).

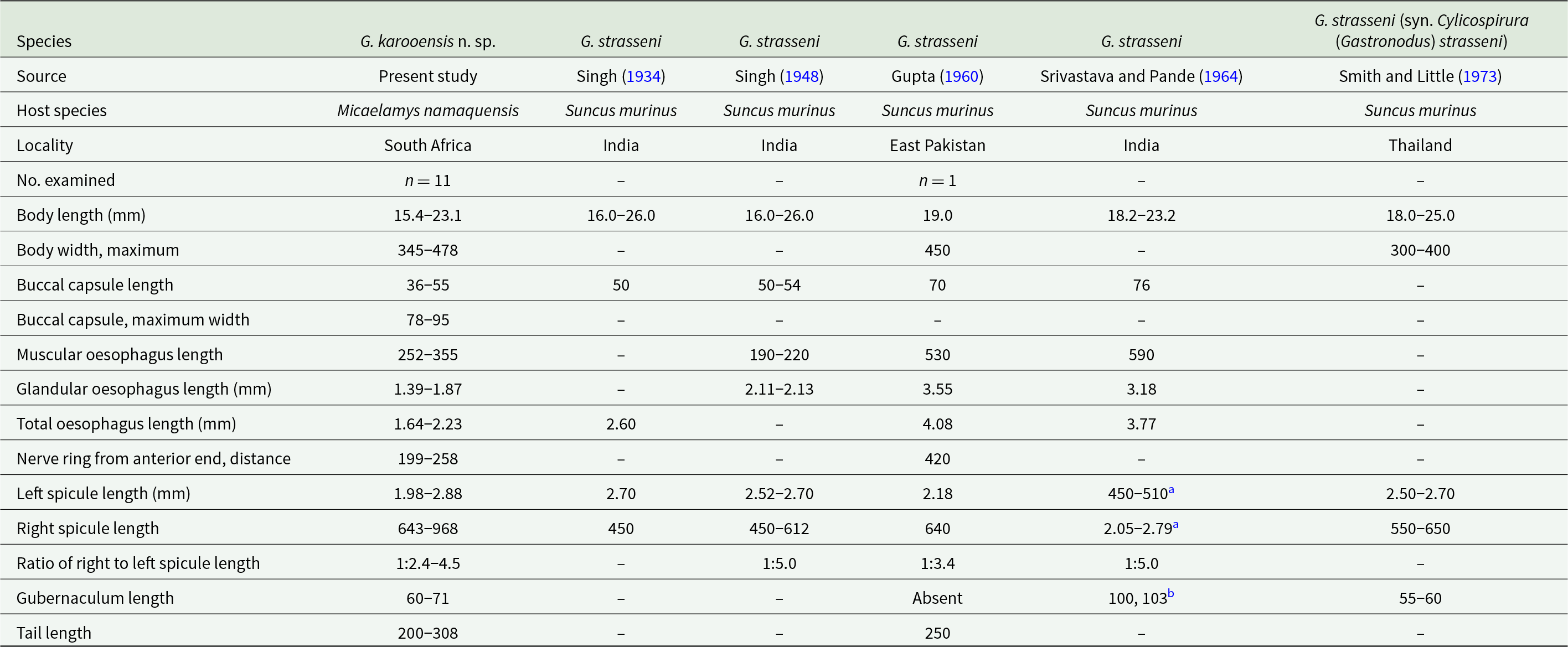
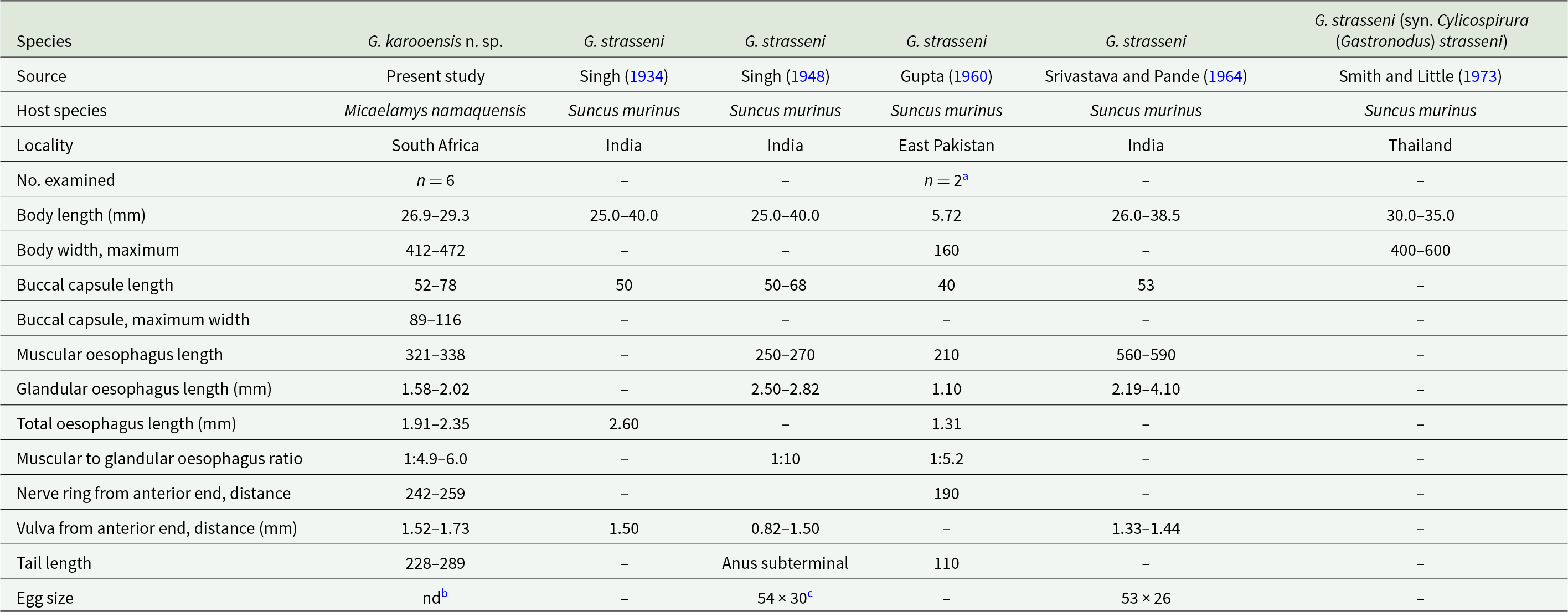
Morphometric data of males of Gastronodus karooensis n. sp. and Gastronodus strasseni as described by various authors

Measurements are in micrometres unless otherwise stated.
a Measurements given as per the original reference; however, the authors likely confused the right and left spicule.
b Srivastava and Pande (Reference Srivastava and Pande1964) refer to the length of the 2 arms of the V-shaped gubernaculum.
Morphometric data of females of Gastronodus karooensis n. sp. and Gastronodus strasseni as described by various authors

Measurements are in micrometres unless otherwise stated.
a According to Gupta (Reference Gupta1960), description is based on 2 immature females.
b Eggs not embryonated yet.
c Eggs embryonated.
Males (based on 11 males; except where otherwise indicated). Body 15.4–23.1 (18.5) mm long. Maximum body width 345–478 (412). Buccal capsule 36–55 (46) long, including cylindrical base 15–30 (22) long, measured at lowest point on muscular oesophagus, and 43–63 (51) wide; maximum width of buccal capsule 78–95 (84). Nerve-ring, deirids and excretory pore at 199–258 (225), 333–457 (397; n = 20) and 397–497 (453) from anterior extremity, respectively. Total length of oesophagus 1638–2227 (1863); i.e. 8.7–12.5% (10.2%) of body length. Muscular oesophagus 252–355 (302) long; 93–110 (100) wide at dilatation and 79–110 (91) wide at level of nerve-ring; glandular oesophagus 1386–1872 (1561) long and 210–283 (246) wide; ratio of muscular to glandular oesophagus length 1:4.4–5.7 (1:5.2). Reflexion of testis at 2068–3795 (3046; n = 10) from anterior extremity. Tail coiled ventrally, 200–308 (256) long. Caudal alae narrow, 495–738 (612; n = 9) long. Area rugosa composed of distinct, fine, parallel, longitudinal cuticular ridges (Figures 2D and 3D); absent in area surrounding cloaca and subterminal papillae; extending 1888 and 2312 anteriorly from tip of tail in 2 specimens measured, 17.4 mm and 23.1 mm long, respectively. Cloaca slit-like, without conspicuous cuticular lips. Caudal papillae as follows: 1 single, large, sessile, median papilla anterior to cloaca present (Figures 1E and 3D); precloacal ventrolateral pedunculate papillae, large, 4 or 5 pairs (in one specimen 5 papillae on right and 4 on left side); at mid-tail usually 3 pairs of large pedunculate papillae present, forming a distinct group (in one specimen only 2 pairs of these were present); near tail tip 3 or 4 pairs of papillae, most anterior pair larger, pedunculate and situated slightly lateral (Figure 3A and D), last 2 or 3 pairs smaller and sessile (Figures 1E and 3A). Phasmids situated between first and second pair of sessile papillae (Figure 3A). Spicules unequal and dissimilar; left spicule 1975–2883 (2461) long and slender, tapering to narrow, rounded hyaline distal tip (Figure 1F); right spicule 643–968 (729) long, robust with broad, rounded, hyaline distal tip (Figures 1G and 2D); spicular ratio 1:2.4–4.5 (1:3.4). Gubernaculum present, triangular in dorsoventral view, 60–71 (66) long (Figure 1H).
Females (based on 6 females, with eggs not embryonated yet; except where otherwise indicated). Body 26.9–29.3 (27.8; n = 5) mm long. Maximum body width at oesophago-intestinal junction 412–472 (447; n = 5); width at vulva and anus 373–440 (414; n = 4) and 224–255 (237; n = 5), respectively. Buccal capsule 52–78 (59) long, including base 27–37 (30) long, measured at lowest point on muscular oesophagus, and 50–68 (57) wide; maximum width of buccal capsule 89–116 (102). Nerve-ring, deirids and excretory pore at 242–259 (252; n = 5), 298–456 (370; n = 11) and 413–504 (441; n = 5) from anterior extremity, respectively. Total length of oesophagus 1905–2353 (2172; n = 5); i.e. 6.9–8.7% (7.8%; n = 5) of body length. Muscular oesophagus 321–338 (329; n = 5) long; 93–139 (117) wide at dilatation and 83–114 (100; n = 5) wide at level of nerve-ring; glandular oesophagus 1584–2015 (1843; n = 5) long and 189–267 (234; n = 5) wide; ratio of muscular to glandular oesophagus length 1:4.9–6.0 (1:5.6; n = 5). Uteri didelphic, opisthodelphic. Vulva without salient lips, at 1522–1734 (1628; n = 4) from anterior extremity, 298–912 (569; n = 4) anterior to oesophago-intestinal junction (Figure 1D), i.e. at 5.6–6.0% (5.8%; n = 4) of body length; vagina simple, directed posteriorly, 1686–2312 (1911; n = 4) long; ovejector convoluted, 1342 long in one female 27.6 mm long. Immature eggs in uterus not yet embryonated, thin-shelled, rounded. Tail conical, slightly bent ventrally, 228–289 (251) long, with rounded tip (Figure 1I); tail length to width at anus ratio 1:0.9–1.1 (1.0; n = 5). Phasmids subterminal.
Molecular characterization
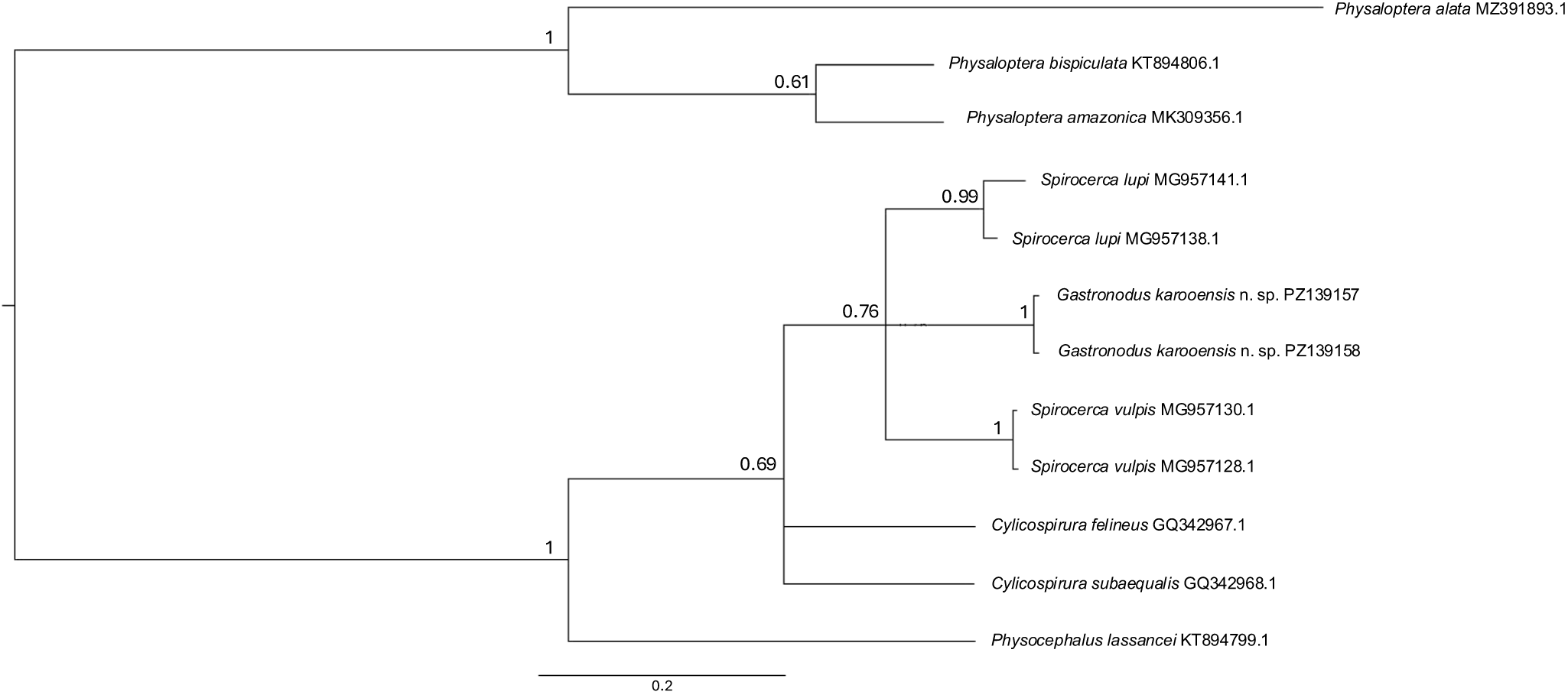
Partial mitochondrial cox1 DNA sequences (386 bp in length) were generated from 2 males that produced identical sequences (GenBank Accession Numbers PZ139157 and PZ139158). The Bayesian inference tree based on the cox1 analysis of G. karooensis n. sp. and closely related species from GenBank (Figure 4) provides support (not significant) for a monophyletic Spirocercinae, comprising Gastronodus, Spirocerca and Cylicospirura. The 3 genera grouped in a weakly supported sister clade to Physocephalus lassancei Travassos, 1921, a member of the spirocercid subfamily Ascaropsinae. Interestingly, within this clade, G. karooensis n. sp. seems more closely related to Spirocerca spp. than to species of Cylicospirura, a genus of which it had formerly been considered a subgenus. Our phylogenetic analysis, positioning the new species in a group with S. lupi and S. vulpis, further suggests that Spirocerca may not be monophyletic
Bayesian inference tree based on the cox1 analysis of Gastronodus karooensis n. sp. and closely related species from GenBank (accession numbers following species names). Physaloptera species were chosen as the outgroup. Posterior probability values are indicated on the branches.

.
Discussion
The subfamily Spirocercinae encompasses spirocercid nematodes with poorly developed lips and a buccal cavity without rugous or annual thickenings but often armed with teeth (Chabaud, Reference Chabaud, Anderson, Chabaud and Willmott1975). In having a hexagonal oral aperture and a short, heavily sclerotized buccal capsule armed with 6 teeth, small deirids situated near the nerve ring, as well as the presence of 3 pairs of pedunculate postcloacal papillae, a gubernaculum and unequal, dissimilar spicules in the males, the present specimens conform to the diagnosis of the subfamily and in particular to that of the genus Gastronodus as amended by Junker et al. (Reference Junker, Lane, McRee, Foggin, van Dyk and Mutafchiev2013). The presence of 3 pairs of pedunculate postcloacal papillae distinguishes our material from the closely related spirocercine genera Cylicospirura and Skrjabinocercina, which possess 2 pairs of pedunculate postcloacal papillae (Junker et al., Reference Junker, Lane, McRee, Foggin, van Dyk and Mutafchiev2013, Reference Junker, de Klerk-lorist, Foggin, Martin and Mutafchiev2024). In addition, the vulva is situated posteriorly, near the anus in females of Skrjabinocercina (Matschulsky, Reference Matschulsky1952), but on the level of the glandular oesophagus in the current material.
A buccal cavity armed with 6 teeth has been described for 3 further genera within the Spirocercinae. Of these, Spiralatus Chabaud, Brygoo and Durette, 1963 and Paraspiralatus Gibbons, Nicholls, Bailey and Samour, 2004 parasitize avian hosts and are characterized by large deirids in a markedly anterior position (Chabaud, Reference Chabaud, Anderson, Chabaud and Willmott1975; Gibbons, Reference Gibbons2010), whereas the deirids are small and situated near the junction between the muscular and glandular oesophagus in the present specimens. Spirocerca Raillet and Henry, 1911 was originally described as having 6 buccal teeth (Raillet and Henry, Reference Raillet and Henry1911). However, Yorke and Maplestone (Reference Yorke and Maplestone1926) elucidated that the teeth of the type species of the genus, Spirocerca sanguinolenta (Rudolphi, 1819), a junior synonym of Spirocerca lupi (Rudolphi, 1809), are ‘small subsidiary papillae situated in the wall of the vestibule just within the mouth’ and that the buccal capsule is unarmed (see also Chabaud, Reference Chabaud, Anderson, Chabaud and Willmott1975). Spirocerca vulpis Rojas et al., 2018, described as a parasite of the red fox, Vulpes vulpes L., in Europe by Rojas et al. (Reference Rojas, Sanchis-Monsonís, Alić, Hodžić, Otranto, Yasur-Landau, Martínez-Carrasco and Baneth2018a), is armed with 6 teeth, thus not conforming to the current diagnosis of Spirocerca. Our sample can be easily distinguished from S. vulpis, by its considerably shorter body (males 15.4–23.1 mm vs 33.9–48.9 mm; females 26.9–29.3 mm vs 41.7–89.5 mm), shorter total length of the oesophagus (males 1.6–2.2 vs 3.8–4.25 mm; females 1.9–2.4 vs 6.1–7.2 mm), vulva situated closer to the anterior end of the body (1.5–1.7 mm vs 8.0–11.5 mm) and males with 3 pairs vs 2 pairs of large pedunculate papillae at mid-tail.
According to Singh (Reference Singh1948), Gastronodus possesses a pair of large sessile papillae and 2 pairs of minute papillae near the tip of the tail. Singh (Reference Singh1948) also mentioned the presence of a gubernaculum, referred to as ‘an accessory piece which is lightly chitinized and forms a V-shaped structure’.
The males of G. karooensis n. sp. have 4–5 pairs of pedunculate precloacal papillae whereas those of G. strasseni, as described by Singh (Reference Singh1934, Reference Singh1948), possess 5–7 pairs of pedunculate precloacal papillae, including the papillae listed as adanal. The males from South Africa have a considerably longer right spicule (643–968) compared to that of G. strasseni (450–612 long), and the left spicule has a round hyalin distal end, whereas Singh (Reference Singh1948) described the distal end of the left spicule as pointed. Singh (Reference Singh1948) states that the anus of the females of G. strasseni is subterminal and their posterior extremity carries 2 large papillae projecting beyond the tip of the tail (figure 6). In contrast, the female tail of the material studied by us is conical, has inconspicuous subterminal phasmids and structures projecting beyond the tail are absent.
Gupta (Reference Gupta1960) and Srivastava and Pande (Reference Srivastava and Pande1964) reported Gastronodus strasseni (as Castronodus strassenii) from the type host in East Pakistan and India, respectively. The authors were apparently unaware of the work by Singh (Reference Singh1948) and provided unsound morphological comparisons of their material with the description of G. strasseni as published by Singh (Reference Singh1934). The material studied by Srivastava and Pande (Reference Srivastava and Pande1964) morphologically resembles that of G. strasseni; however, the sample studied by Gupta (Reference Gupta1960) differs from G. strasseni in the presence of 2 median teeth larger than the remaining teeth, the lack of a gubernaculum and a considerably smaller ratio between the left and right spicule (1:3.5) and could, in fact, belong to a different species. Gastronodus strasseni was also reported from S. murinus in Thailand by Smith and Little (Reference Smith and Little1973).
Gupta and Trivedi (Reference Gupta and Trivedi1985) described ‘Castronodus muelleri’ as a new species from S. murinus in India. However, based on males possessing 5 precloacal, 1 adcloacal and 6 postcloacal pairs of pedunculate papillae, a narrow conical tail in females and females laying unembryonated eggs, Junker et al. (Reference Junker, Lane, McRee, Foggin, van Dyk and Mutafchiev2013) considered this a species incertae sedis. The presence of a median spinous tail process in the males of the specimens described by Gupta and Trivedi (Reference Gupta and Trivedi1985) further distinguishes them from the present specimens.
Based on the above morphological differences between previous descriptions of G. strasseni and our current material, we conclude that the present specimens represent a new species for which we suggest the name G. karooensis. This is the first report of a member of this genus from the Afrotropics, and its presence in the murid M. namaquensis constitutes a new host record, having previously been restricted to reports from a soricid host in Indomalaysia (Singh, Reference Singh1934, Reference Singh1948; Gupta, Reference Gupta1960; Srivastava and Pande, Reference Srivastava and Pande1964; Smith and Little, Reference Smith and Little1973).
While Spirocerca spp., Cylicospirura spp. and G. strasseni have typically been found in granulomata in the stomach and oesophagus (Baylis, Reference Baylis1927; Singh, Reference Singh1934, Reference Singh1948; Gupta, Reference Gupta1960; Srivastava and Pande, Reference Srivastava and Pande1964; Smith and Little, Reference Smith and Little1973; Pence et al., Reference Pence, Samoil and Stone1978; Waid and Pence, Reference Waid and Pence1988; Junker et al., Reference Junker, Lane, McRee, Foggin, van Dyk and Mutafchiev2013; Rojas et al., Reference Rojas, Dvir, Farkas, Sarma, Borthakur, Jabbar, Markovics, Otranto and Baneth2018b), our finding of G. karooensis n. sp. occurring free in the stomach of its host is not unheard of in the Spirocercinae. Junker et al. (Reference Junker, de Klerk-lorist, Foggin, Martin and Mutafchiev2024) found Cylicospirura phiri Junker and Mutafchiev, 2024 free in the cranial part of the oesophagus as well as in open granulomata in the stomach of Crocuta crocuta (Erxleben, 1777). However, the stomach of the single infected female of M. namaquenis was in poor condition and it is possible that open granulomata could no longer be identified as such. More samples would be needed to establish if G. karooensis n. sp. typically occurs free in the lumen of the stomach or is associated with granulomatous lesions.
In addition to the 20 hosts examined at the type locality, a further 162 M. namaquensis as well as numerous Rhabdomys spp. (n = 237) and Otomys unisulcatus (n = 157) were examined at various localities in the Western Cape Province, South Africa, as part of the same study (Schlemmer et al., Reference Schlemmer, Junker, Matthee and Matthee2026). None of these harboured representatives of the new species. Similarly, a previous study, including 13 rodent species collected at 26 localities across South Africa (Spickett et al., Reference Spickett, Junker, Froeschke, Haukisalmi and Matthee2020), did not yield any Gastronodus specimens, suggesting that G. karooensis n. sp. is not a common or widespread parasite of African rodents. This might be supported by the fact that only unembryonated eggs were observed in the females’ uteri. Its preferred host might be a species of shrew as seen in its congener G. strasseni. The helminth fauna of shrews in South Africa remains poorly studied; however, in 30 shrews collected at various localities in South Africa as part of the study of Spickett et al. (Reference Spickett, Junker, Froeschke, Haukisalmi and Matthee2020), Gastronodus was absent (unpublished data).
Our phylogenetic analysis provides preliminary evidence that G. karooensis n. sp. most likely belongs to the Spirocercidae, specifically the subfamily Spirocercinae, and revealed some affiliation with the genus Spirocerca. The fact that S. lupi and S. vulpis did not form a monophyletic group, but grouped with G. karooensis n. sp., is noteworthy, especially given the fact that S. vulpis does not conform to the current diagnosis of Spirocerca. However, posterior probability support for the nodes was non-significant, most likely due to the short fragment sequenced. Sampling of a wider taxonomic range within the Spirocercinae, and the addition of more molecular markers, will be needed to ultimately resolve the phylogenetic relations within this subfamily.
Data availability statement
All data generated or analysed during the study are included in this published article. The datasets used and/or analyses are available from the corresponding author upon request.
Acknowledgements
We sincerely thank the farm owners (Kobus and Helena Marais) for granting us permission to conduct fieldwork on their farm. We are grateful to J Kipling and A Little for their support in the field.
Author contributions
ES: Sample collection, morphological, molecular and phylogenetic investigation and analyses, writing – original draft, writing – review and editing. KJ: Morphological investigation and analysis, resources, supervision, visualization (light microscopy, original line drawings), writing – original draft, writing – review and editing. YM: Morphological investigation and analysis, resources, visualization (scanning electron microscopy, digital inking), writing – original draft, writing – review and editing. CAM: Sample collection, molecular and phylogenetic investigation and analysis, resources, supervision, writing – review and editing. SM: Conceptualization, project administration, sample collection, resources, supervision, writing – review and editing. All authors read and approved the final manuscript.
Financial support
KJ received funding from the Department of Agriculture, South Africa (project number A150). CM received funding from the National Research Foundation (NRF) in South Africa (Grant reference numbers: GUN 129276 and FBIS22100761503). Any opinion, finding and conclusion or recommendation expressed in this material is that of the authors and the NRF does not accept any liability in this regard.
Competing interests
The authors declare there are no conflicts of interest.
Ethical standards
The study was approved by Animal Care and Use Ethics Committee of Stellenbosch University (reference numbers: ACU-2022-17062, ACU-2024-30885). A Section 20 permit from Department of Agriculture, Land Reform and Rural Development (no:12/11/1/7/5 (b) (6091MG), as well as collection permit from Cape Nature (Western Cape Provincial Nature Conservation authority) CN44-87-27373 and CN44-87-33283.

 Open access
Open access