Refine search
Actions for selected content:
10186 results in Genetics
Errata
-
- Journal:
- Genetical Research / Volume 10 / Issue 1 / August 1967
- Published online by Cambridge University Press:
- 14 April 2009, p. 119
-
- Article
-
- You have access
- Export citation
-
Recombination within the Y locus in Ascobolus immersus
By A. Kruszewska and W. Gajewski
Vol. 9, No. 2, p. 159 (1967)
In the legend to Fig. 1, p. 165, for ‘(in %)’ on lines 4 and 5 substitute ‘(× 103)’.
Recombination and incompatibility in Schizophyllum
By G. Simchen
Vol. 9, No. 2, p. 195 (1967)
On p. 207, line 9, for ‘½(nα + 1)’ read
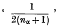 .
.
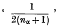 .
.Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 63 / Issue 3 / June 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 237-239
-
- Article
-
- You have access
- Export citation
Effective population size/adult population size ratios in wildlife: a review
-
- Journal:
- Genetical Research / Volume 66 / Issue 2 / October 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 95-107
-
- Article
-
- You have access
- Export citation
Genetic relationships between inbred strains of rats. An analysis based on genetic markers at 28 biochemical loci
-
- Journal:
- Genetical Research / Volume 44 / Issue 3 / December 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 271-281
-
- Article
-
- You have access
- Export citation
Some sampling properties of selectively neutral alleles: Effects of variability of mutation rates
-
- Journal:
- Genetical Research / Volume 34 / Issue 3 / December 1979
- Published online by Cambridge University Press:
- 14 April 2009, pp. 253-267
-
- Article
-
- You have access
- Export citation
Y-chromosomal DNA polymorphism in mouse inbred strains
-
- Journal:
- Genetical Research / Volume 50 / Issue 1 / August 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 69-72
-
- Article
-
- You have access
- Export citation
The liver/erythrocyte pyruvate kinase gene complex [Pk-1] in the mouse: regulatory gene mutations
-
- Journal:
- Genetical Research / Volume 58 / Issue 3 / December 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 233-241
-
- Article
-
- You have access
- Export citation
Genetic models for the inheritance of the silver colour mutation of foxes
-
- Journal:
- Genetical Research / Volume 64 / Issue 1 / August 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 11-18
-
- Article
-
- You have access
- Export citation
Mutagen specificity in conversion pattern in Sordaria brevicollis
-
- Journal:
- Genetical Research / Volume 29 / Issue 1 / February 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 65-81
-
- Article
-
- You have access
- Export citation
A t-haplotype (tw75) overlapping two complementation groups
-
- Journal:
- Genetical Research / Volume 33 / Issue 3 / June 1979
- Published online by Cambridge University Press:
- 14 April 2009, pp. 279-285
-
- Article
-
- You have access
- Export citation
Genetical studies on growth and form in the fowl 1. Phenotypic variation in the relative growth pattern of shank length and body-weight
-
- Journal:
- Genetical Research / Volume 4 / Issue 2 / July 1963
- Published online by Cambridge University Press:
- 14 April 2009, pp. 167-192
-
- Article
-
- You have access
- Export citation
The quantitative relations between variation in red eye pigment and related pteridine compounds in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 15 / Issue 1 / February 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 65-86
-
- Article
-
- You have access
- Export citation
Origin of rare Ha-ras alleles: relationship of VTR length to a 5′ polymorphic Xho I site
-
- Journal:
- Genetical Research / Volume 54 / Issue 2 / October 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 149-153
-
- Article
-
- You have access
- Export citation
Zigzag: a genetic defect of the horizontal canals in the mouse
-
- Journal:
- Genetical Research / Volume 1 / Issue 2 / July 1960
- Published online by Cambridge University Press:
- 14 April 2009, pp. 189-195
-
- Article
-
- You have access
- Export citation
Studies of aromatic biosynthetic and catabolic enzymes in Ustilago maydis and in mutants of U. violacea
-
- Journal:
- Genetical Research / Volume 19 / Issue 3 / June 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 261-270
-
- Article
-
- You have access
- Export citation
Rates of spontaneous mitotic recombination in Saccharomyces cerevisiae
-
- Journal:
- Genetical Research / Volume 18 / Issue 2 / October 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 147-151
-
- Article
-
- You have access
- Export citation
Gene Manipulation in Fungi. Edited by J. W. Bennett and Linda L. Lasure. Orlando: Academic Press. 558 Pages. Cloth £65, Paper £35. ISBN 0 12 088640 5.
-
- Journal:
- Genetical Research / Volume 47 / Issue 3 / June 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 232-233
-
- Article
-
- You have access
- Export citation
Consequences of double cross-over detections on the functional interpretation of segment ‘29’ in Podospora anserina
-
- Journal:
- Genetical Research / Volume 19 / Issue 3 / June 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 313-319
-
- Article
-
- You have access
- Export citation
The pattern of the parasexual cycle in Aspergillus amstelodami
-
- Journal:
- Genetical Research / Volume 5 / Issue 1 / February 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 162-163
-
- Article
-
- You have access
- Export citation
Transformation in Escherichia coli: studies on the nature of donor DNA after uptake and integration
-
- Journal:
- Genetical Research / Volume 35 / Issue 3 / June 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 279-289
-
- Article
-
- You have access
- Export citation
