Refine search
Actions for selected content:
10186 results in Genetics
Herbicide-tolerant tobacco mutants selected in situ and recovered via regeneration from cell culture
-
- Journal:
- Genetical Research / Volume 32 / Issue 1 / August 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. 85-89
-
- Article
-
- You have access
- Export citation
GRH volume 48 issue 3 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 48 / Issue 3 / December 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b7
-
- Article
-
- You have access
- Export citation
Compatibility and stability of diploids in Coprinus lagopus
-
- Journal:
- Genetical Research / Volume 8 / Issue 1 / August 1966
- Published online by Cambridge University Press:
- 14 April 2009, pp. 61-72
-
- Article
-
- You have access
- Export citation
Biochemistry and Molecular Biology of Plant- Pathogen Interactions. Edited by C. J. Smith. Clarendon Press, Oxford. 1991. 291 pages. £45.00. ISBN 0 19 857734 6.
-
- Journal:
- Genetical Research / Volume 59 / Issue 2 / April 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. 149-150
-
- Article
-
- You have access
- Export citation
Somatic reversion of P transposable element insertion mutations in the singed locus of Drosophila melanogaster requiring specific P insertions and a trans- acting factor
-
- Journal:
- Genetical Research / Volume 54 / Issue 2 / October 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 101-112
-
- Article
-
- You have access
- Export citation
Novel pleiotropic effect of rifampicin resistance mutation in a Micromonospora sp
-
- Journal:
- Genetical Research / Volume 40 / Issue 1 / August 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 33-40
-
- Article
-
- You have access
- Export citation
Selection for recombination in a polygenic model
-
- Journal:
- Genetical Research / Volume 35 / Issue 3 / June 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 269-277
-
- Article
-
- You have access
- Export citation
Some additional results on the maintenance of kappa particles in Paramecium aurelia (stock 51) after loss of the gene K
-
- Journal:
- Genetical Research / Volume 9 / Issue 3 / June 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 369-373
-
- Article
-
- You have access
- Export citation
Dosage dependent modifiers of white alleles in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 62 / Issue 1 / August 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. 15-22
-
- Article
-
- You have access
- Export citation
Dynamics of polygenic variability under stabilizing selection, recombination, and drift
-
- Journal:
- Genetical Research / Volume 65 / Issue 1 / February 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 63-74
-
- Article
-
- You have access
- Export citation
Variations in resistance to three antibiotics among some single-step mutants to Chloramphenicol resistance in a strain of Escherichia coli K 12
-
- Journal:
- Genetical Research / Volume 6 / Issue 2 / July 1965
- Published online by Cambridge University Press:
- 14 April 2009, pp. 310-315
-
- Article
-
- You have access
- Export citation
A recombination test to classify mutants of Bacillus subtilis of identical phenotype
-
- Journal:
- Genetical Research / Volume 27 / Issue 1 / February 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 47-58
-
- Article
-
- You have access
- Export citation
The incidence of chromosomally unbalanced gametes in T(14; 15)6 Ca heterozygote mice
-
- Journal:
- Genetical Research / Volume 27 / Issue 1 / February 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 77-84
-
- Article
-
- You have access
- Export citation
A buff spore colour mutant in Sordaria brevicollis showing high-frequency conversion: 2. Loss of the high-frequency conversion
-
- Journal:
- Genetical Research / Volume 41 / Issue 2 / April 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 155-163
-
- Article
-
- You have access
- Export citation
The effects of inbreeding on the components of litter size in mice
-
- Journal:
- Genetical Research / Volume 10 / Issue 1 / August 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 73-80
-
- Article
-
- You have access
- Export citation
Genetics of plasma transferrins in the mouse
-
- Journal:
- Genetical Research / Volume 1 / Issue 3 / November 1960
- Published online by Cambridge University Press:
- 14 April 2009, pp. 431-438
-
- Article
-
- You have access
- Export citation
Books Received
-
- Journal:
- Genetical Research / Volume 59 / Issue 1 / February 1992
- Published online by Cambridge University Press:
- 14 April 2009, p. 71
-
- Article
-
- You have access
- Export citation
Hybrid dysgenesis in Drosophila melanogaster: partial sterility associated with embryo lethality in the P-M system
-
- Journal:
- Genetical Research / Volume 44 / Issue 1 / August 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 11-28
-
- Article
-
- You have access
- Export citation
Theoretical study on genetic variation in multigene families*
-
- Journal:
- Genetical Research / Volume 31 / Issue 1 / February 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. 13-28
-
- Article
-
- You have access
- Export citation
-
Genetic variation contained in a multigene family was theoretically investigated from the standpoint of population genetics. Unequal crossover is assumed to be responsible for the coincidental evolution of mutant genes in a chromosome. When the allowed latitude of the duplicated or deleted number of gene units at unequal crossover is 10 ˜ 15% of the total gene number in a chromosome, the arrangement of gene lineage in a chromosome is shown to be roughly random. The equilibrium properties of genetic variation or the probability of identity of two genes within a family (clonality) were studied under mutation, unequal crossover, interchromosomal crossover and sampling of gametes. The clonality of a multigene family within a chromosome is shown to be approximately
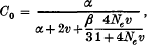
in which α = 2k/n2 with k = effective number of cycles of unequal crossover and with n = number of gene units in a family, v is the mutation rate per gene unit, β is the rate of interchromosomal crossover per family and Ne is the effective size of the population, all measured by the rate per generation. The clonality of a gene family between two different chromosomes becomes approximately C1 = C0/(l + 4Neν). Some models of natural selection which lowers the clonality or increases genetic variation in a multigene family were investigated. It was shown that natural selection may be quite effective in increasing genetic variation in a gene family.
GRH volume 57 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 57 / Issue 2 / April 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
