Refine search
Actions for selected content:
10186 results in Genetics
Erratum
-
- Journal:
- Genetical Research / Volume 40 / Issue 1 / August 1982
- Published online by Cambridge University Press:
- 14 April 2009, p. 107
-
- Article
-
- You have access
- Export citation
-
The interpretation of gene conversion data from ordered eight-spored asci
By J. R. S. Fincham, W. G. Hill and E. C. R. Reeve
Genetical Research, Vol. 35, No. 2, 1980
On p. 181 the phrases ‘wild to mutant’ and ‘mutant to wild’ in lines 8–9 in the third paragraph should be interchanged: p and r are correction frequencies from mutant to wild and q and s are correction frequencies from wild to mutant, as is apparent in the Table immediately following.
On p. 188 the calculated ratios p/q and r/s may appear to be at variance with the values given in Table 2. This is due to the latter having been rounded to two significant figures. The more accurate values for the + × h3 cross are as follows: p = 0·043, q = 0·161, r = 0·157, s = 0·426 (firstsolution) and p′ = 0·026, q′ = 0·493, r′ = 0·243 and s′ = 0·128 (second solution).
On p. 191 (foot of page) some primes are misplaced. The correct formulae for r, q and s (as given on p. 182) are:
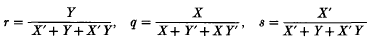
On p. 194, line 7, G = 2B + C should read G = 2B + D.
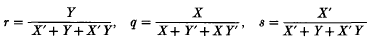
Gametic selection and the selection component analysis
-
- Journal:
- Genetical Research / Volume 37 / Issue 3 / June 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 275-284
-
- Article
-
- You have access
- Export citation
Chloride channel 2 gene (Clc2) maps to chromosome 16 of the mouse, extending a region of conserved synteny with human chromosome 3q
-
- Journal:
- Genetical Research / Volume 66 / Issue 2 / October 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 175-178
-
- Article
-
- You have access
- Export citation
The initiation and development of the hair follicle population in tabby mice
-
- Journal:
- Genetical Research / Volume 10 / Issue 2 / October 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 161-171
-
- Article
-
- You have access
- Export citation
GRH volume 12 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 12 / Issue 3 / December 1968
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f5
-
- Article
-
- You have access
- Export citation
Egg albumen polymorphisms in the fowl: The ovalbumin locus
-
- Journal:
- Genetical Research / Volume 5 / Issue 2 / July 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 257-268
-
- Article
-
- You have access
- Export citation
A cya deletion mutant of Escherichia coli develops thermotolerance but does not exhibit a heat-shock response
-
- Journal:
- Genetical Research / Volume 55 / Issue 1 / February 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-6
-
- Article
-
- You have access
- Export citation
GRH volume 10 issue 3 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 10 / Issue 3 / December 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b7
-
- Article
-
- You have access
- Export citation
Antigenic polymorphism in a wild population of Paramecium aurelia
-
- Journal:
- Genetical Research / Volume 1 / Issue 1 / February 1960
- Published online by Cambridge University Press:
- 14 April 2009, pp. 62-68
-
- Article
-
- You have access
- Export citation
On sojourn times at particular gene frequencies
-
- Journal:
- Genetical Research / Volume 25 / Issue 2 / April 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 89-94
-
- Article
-
- You have access
- Export citation
Inactivation of whole chromosomes in mammals and coccids: some comparisons*
-
- Journal:
- Genetical Research / Volume 18 / Issue 3 / December 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 265-276
-
- Article
-
- You have access
- Export citation
Sex ratio distortion in D. simulans after treatment with quinacrine
-
- Journal:
- Genetical Research / Volume 49 / Issue 1 / February 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 19-23
-
- Article
-
- You have access
- Export citation
Complex Organismal Functions: Integration and Evolution in Vertebrates. Edited by D. B. Wake and G. Roth. Chichester, UK: John Wiley and Sons. 1989. 451 pages. Cloth (£57.50). ISBN 0 471 92375 3.
-
- Journal:
- Genetical Research / Volume 55 / Issue 3 / June 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 223-224
-
- Article
-
- You have access
- Export citation
Gene Structure and Expression. By J. D. Hawkins. Cambridge University Press. 1985. 173 pages. £20.000 hardback, £7.95 paperback. ISBN 0 521 25824 3
-
- Journal:
- Genetical Research / Volume 46 / Issue 2 / October 1985
- Published online by Cambridge University Press:
- 14 April 2009, p. 237
-
- Article
-
- You have access
- Export citation
A correction to the exact test based on the Ewens sampling distribution
-
- Journal:
- Genetical Research / Volume 68 / Issue 3 / December 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 259-260
-
- Article
-
- You have access
- Export citation
Response to selection for increased pupa weight in Tribolium castaneum as related to population structure*
-
- Journal:
- Genetical Research / Volume 30 / Issue 3 / December 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 237-246
-
- Article
-
- You have access
- Export citation
Production of auxotrophic mutants in ferns
-
- Journal:
- Genetical Research / Volume 14 / Issue 3 / December 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 337-339
-
- Article
-
- You have access
- Export citation
The effects of inhibition of protein synthesis on the phenotype of Abnormal-abdomen*
-
- Journal:
- Genetical Research / Volume 21 / Issue 3 / June 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 229-238
-
- Article
-
- You have access
- Export citation
The nature of the purebred-crossbred genetic covariance*
-
- Journal:
- Genetical Research / Volume 18 / Issue 1 / August 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-7
-
- Article
-
- You have access
- Export citation
Exploiting New Technologies in Animal Breeding: Genetic Developments. Edited by C. Smith, J. W. B. King, and J. C. MacKay. Oxford University Press, 1986. 216 pages. £30. ISBN 0 19 854 209 7.
-
- Journal:
- Genetical Research / Volume 51 / Issue 1 / February 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 77-79
-
- Article
-
- You have access
- Export citation
