Refine search
Actions for selected content:
10186 results in Genetics
The effects on litter size of crossing lines of mice inbred without selection
-
- Journal:
- Genetical Research / Volume 1 / Issue 2 / July 1960
- Published online by Cambridge University Press:
- 14 April 2009, pp. 239-252
-
- Article
-
- You have access
- Export citation
Long-term selection for a quantitative character in large replicate populations of Drosophila melanogaster: 1. Response to selection
-
- Journal:
- Genetical Research / Volume 35 / Issue 1 / February 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-17
-
- Article
-
- You have access
- Export citation
-
The response to long-term selection for increased abdominal bristle number was studied in six replicate lines of Drosophila melanogaster derived from the sc Canberra outbred strain. Each line was continued for 86–89 generations with 50 pairs of parents selected at an intensity of 20%, and subsequently for 32–35 generations without selection. Response continued for at least 75 generations and average total response was in excess of 36 additive genetic standard deviations of the base population (σA) or 51 times the response in the first generation. The pattern of longterm response was diverse and unpredictable typically with one or more accelerated responses in later generations. At termination of the selection, most of the replicate lines were extremely unstable with high phenotypic variability, and lost much of their genetic gains rapidly upon relaxation of selection.
The variation in response among replicates rose in the early phase of selection to level off at approximately 7·6
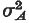 around generation 25. As some lines plateaued, it increased further to a level higher than would be accommodated by most genetic models. The replicate variation was even higher after many generations of relaxed selection. The genetic diversity among replicates, as revealed in total response, the individuality of response patterns and variation of the sex-dimorphism ratio, suggests that abdominal bristle number is influenced potentially by a large number of genes, but a smaller subset of them was responsible for selection response in any one line.
around generation 25. As some lines plateaued, it increased further to a level higher than would be accommodated by most genetic models. The replicate variation was even higher after many generations of relaxed selection. The genetic diversity among replicates, as revealed in total response, the individuality of response patterns and variation of the sex-dimorphism ratio, suggests that abdominal bristle number is influenced potentially by a large number of genes, but a smaller subset of them was responsible for selection response in any one line.
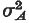 around generation 25. As some lines plateaued, it increased further to a level higher than would be accommodated by most genetic models. The replicate variation was even higher after many generations of relaxed selection. The genetic diversity among replicates, as revealed in total response, the individuality of response patterns and variation of the sex-dimorphism ratio, suggests that abdominal bristle number is influenced potentially by a large number of genes, but a smaller subset of them was responsible for selection response in any one line.
around generation 25. As some lines plateaued, it increased further to a level higher than would be accommodated by most genetic models. The replicate variation was even higher after many generations of relaxed selection. The genetic diversity among replicates, as revealed in total response, the individuality of response patterns and variation of the sex-dimorphism ratio, suggests that abdominal bristle number is influenced potentially by a large number of genes, but a smaller subset of them was responsible for selection response in any one line.Dynamics of gametic disequilibria between loci linked to chromosome inversions: the recombination-redistributing effect of inversions
-
- Journal:
- Genetical Research / Volume 67 / Issue 1 / February 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 67-76
-
- Article
-
- You have access
- Export citation
Studies on genetic organization in higher organisms: III. Confirmation of the single cistron-allele complementation model of organization of the maroon-like region of Drosophila melanogaster*
-
- Journal:
- Genetical Research / Volume 15 / Issue 3 / June 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 351-355
-
- Article
-
- You have access
- Export citation
GRH volume 43 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 43 / Issue 2 / April 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
GRH volume 37 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 37 / Issue 2 / April 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
The highly polarized recombination pattern within the methA gene of Aspergillus nidulans Recombination within the methA cistron of A. nidulans
-
- Journal:
- Genetical Research / Volume 17 / Issue 2 / April 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 125-131
-
- Article
-
- You have access
- Export citation
The role of growth hormone in lines of mice divergently selected on body weight
-
- Journal:
- Genetical Research / Volume 61 / Issue 2 / April 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. 101-106
-
- Article
-
- You have access
- Export citation
GRH volume 39 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 39 / Issue 1 / February 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Reduction of wild-type X chromosomes with the Ybb− chromosome of Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 43 / Issue 1 / February 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 93-98
-
- Article
-
- You have access
- Export citation
The estimation of generation interval in experimental populations of Drosophila
-
- Journal:
- Genetical Research / Volume 3 / Issue 3 / November 1962
- Published online by Cambridge University Press:
- 14 April 2009, pp. 388-404
-
- Article
-
- You have access
- Export citation
On Competition by C. T. de Wit. First published in 1960 and now reprinted as Evolutionary Monographs, vol. 7 by University of Chicago Press, 915 East 57th St, Chicago, IL 60637, USA. 1986. 82 pages. $10.00 (institutions); $8.00 (individuals).
-
- Journal:
- Genetical Research / Volume 50 / Issue 1 / August 1987
- Published online by Cambridge University Press:
- 14 April 2009, p. 82
-
- Article
-
- You have access
- Export citation
GRH volume 14 issue 3 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 14 / Issue 3 / December 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b7
-
- Article
-
- You have access
- Export citation
An SD (Segregation Distribution) – MR (Male Recombination) chromosome isolated from a natural population of Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 43 / Issue 2 / April 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 149-158
-
- Article
-
- You have access
- Export citation
Mammals from Pouches and Eggs: Genetics, Breeding and Evolution of Marsupials and Monotremes. Edited by Jennifer A. Marshall Graves, Rory M. Hope and Desmond W. Cooper. CSIRO Publications, PO Box 89, 314 Gilbert Street, East Melbourne, Victoria 3002, Australia. 1990. 337 pages. Hardback US $50.00. ISBN 0 643 05020 5.
-
- Journal:
- Genetical Research / Volume 58 / Issue 2 / October 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 179-180
-
- Article
-
- You have access
- Export citation
A simple method for increasing the response to artificial selection
-
- Journal:
- Genetical Research / Volume 44 / Issue 3 / December 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 347-349
-
- Article
-
- You have access
- Export citation
The genetic implications of u.v. light exposure and liquid-holding post-treatment in the yeast Saccharomyces cerevisiae
-
- Journal:
- Genetical Research / Volume 19 / Issue 1 / February 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-16
-
- Article
-
- You have access
- Export citation
Genetic structure of a natural population of Coriolus versicolor (L. ex Fr.) Quél.
-
- Journal:
- Genetical Research / Volume 32 / Issue 1 / August 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. 55-65
-
- Article
-
- You have access
- Export citation
Genetics of Neuropsychiatric Diseases. Edited by L. Wetterberg. London: Macmillan Press. 1989. 363 pages. £50.00. ISBN 0 935859 65 9.
-
- Journal:
- Genetical Research / Volume 54 / Issue 3 / December 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 242-243
-
- Article
-
- You have access
- Export citation
The genetics of tasting in mice VII. Glycine revisited, and the chromosomal location of Sac and Soa
-
- Journal:
- Genetical Research / Volume 66 / Issue 2 / October 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 167-174
-
- Article
-
- You have access
- Export citation
