Refine search
Actions for selected content:
10186 results in Genetics
The phasmid as a tool for plasmid genetics: II. Isolation of point mutations that affect replication of a ColE1-related plasmid
-
- Journal:
- Genetical Research / Volume 40 / Issue 3 / December 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 233-247
-
- Article
-
- You have access
- Export citation
Mitotic recombination in the paba 1 cistron of Aspergillus nidulans
-
- Journal:
- Genetical Research / Volume 5 / Issue 2 / July 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 316-327
-
- Article
-
- You have access
- Export citation
Persistence of a lethal t haplotype in a laboratory stock of outbred mice
-
- Journal:
- Genetical Research / Volume 43 / Issue 1 / February 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 21-25
-
- Article
-
- You have access
- Export citation
GRH volume 54 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 54 / Issue 3 / December 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
GRH volume 3 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 3 / Issue 1 / February 1962
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f7
-
- Article
-
- You have access
- Export citation
Antisuppressor mutations in Aspergillus nidulans: cold-resistant revertants of suppressor suaC109
-
- Journal:
- Genetical Research / Volume 49 / Issue 3 / June 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 191-200
-
- Article
-
- You have access
- Export citation
GRH volume 38 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 38 / Issue 2 / October 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
GRH volume 29 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 29 / Issue 2 / April 1977
- Published online by Cambridge University Press:
- 14 April 2009, p. b1
-
- Article
-
- You have access
- Export citation
Acriflavin resistant rII deletions of bacteriophage T4
-
- Journal:
- Genetical Research / Volume 13 / Issue 3 / June 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 329-331
-
- Article
-
- You have access
- Export citation
GRH volume 39 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 39 / Issue 2 / April 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Books Received
-
- Journal:
- Genetical Research / Volume 62 / Issue 2 / October 1993
- Published online by Cambridge University Press:
- 14 April 2009, p. 167
-
- Article
-
- You have access
- Export citation
A genetic analysis of non-disjunction and mitotic recombination in Neurospora crassa
-
- Journal:
- Genetical Research / Volume 16 / Issue 1 / August 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 29-35
-
- Article
-
- You have access
- Export citation
The age of an allele in a finite population*
-
- Journal:
- Genetical Research / Volume 23 / Issue 2 / April 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 137-143
-
- Article
-
- You have access
- Export citation
Genetic characterization of rec-1, a mutant of Ustilago maydis defective in repair and recombination
-
- Journal:
- Genetical Research / Volume 27 / Issue 3 / June 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 413-453
-
- Article
-
- You have access
- Export citation
Dynamics of unconditionally deleterious mutations: Gaussian approximation and soft selection
-
- Journal:
- Genetical Research / Volume 65 / Issue 2 / April 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 113-121
-
- Article
-
- You have access
- Export citation
-
This paper studies the influence of two opposite forces, unidirectional unconditionally deleterious mutations and directional selection against them, on an amphimictic population. Mutant alleles are assumed to be equally deleterious and rare, so that homozygous mutations can be ignored. Thus, a genotype is completely described by its value with respect to a quantitative trait x, the number of mutations it carries, while a population is described by its distribution p(x) with mean M[p] and variance V[p] = σ2[p]. When mutations are only slightly deleterious, so that M » 1, before selection p(x) is close to Gaussian with any mode of selection. I assume that selection is soft in the sense that the fitness of a genotype depends on the difference between its value of x and M, in units of σ. This leads to a simple system of equations connecting the values of M and V in successive generations. This system has a unique and stable equilibrium,
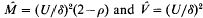 where U is the genomic deleterious mutation rate, δ is the selection differential for x in units of σ, and p is the ratio of variances of p(x) after and before selection. Both δ and ρ are parameters of the mode of soft selection, and do not depend on M or V. In an equilibrium population, the selection coefficient against a mutant allele is ŝ = δ2[U(2–ρ)]−1. The mutation load can be tolerable only if the genome degradation rate υ = U/σ is below 2. Other features of mutation-selection equilibrium are also discussed.
where U is the genomic deleterious mutation rate, δ is the selection differential for x in units of σ, and p is the ratio of variances of p(x) after and before selection. Both δ and ρ are parameters of the mode of soft selection, and do not depend on M or V. In an equilibrium population, the selection coefficient against a mutant allele is ŝ = δ2[U(2–ρ)]−1. The mutation load can be tolerable only if the genome degradation rate υ = U/σ is below 2. Other features of mutation-selection equilibrium are also discussed.
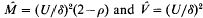 where U is the genomic deleterious mutation rate, δ is the selection differential for x in units of σ, and p is the ratio of variances of p(x) after and before selection. Both δ and ρ are parameters of the mode of soft selection, and do not depend on M or V. In an equilibrium population, the selection coefficient against a mutant allele is ŝ = δ
where U is the genomic deleterious mutation rate, δ is the selection differential for x in units of σ, and p is the ratio of variances of p(x) after and before selection. Both δ and ρ are parameters of the mode of soft selection, and do not depend on M or V. In an equilibrium population, the selection coefficient against a mutant allele is ŝ = δA comparison of genetic variability at X-linked and autosomal loci in kangaroos, man and Drosophila
-
- Journal:
- Genetical Research / Volume 33 / Issue 3 / June 1979
- Published online by Cambridge University Press:
- 14 April 2009, pp. 243-252
-
- Article
-
- You have access
- Export citation
Editorial - New Directions
-
- Journal:
- Genetical Research / Volume 36 / Issue 3 / December 1980
- Published online by Cambridge University Press:
- 14 April 2009, p. i
-
- Article
-
- You have access
- Export citation
Reconstruction of allopolyploid Brassicas through non-homologous recombination: introgression of resistance to pod shatter in Brassica napus
-
- Journal:
- Genetical Research / Volume 56 / Issue 1 / August 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-2
-
- Article
-
- You have access
- Export citation
Microbial Development. Edited by Richard Losick and Lucy Shapiro. New York: Cold Spring Harbor Laboratory. 1984. 303 pages. Paperback $28.00 ($33.60 outside U.S.), hardback $52.00 ($62.40 outside U.S.). ISBN 0 87969 173 5.
-
- Journal:
- Genetical Research / Volume 45 / Issue 3 / June 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 346-347
-
- Article
-
- You have access
- Export citation
Genetic variation in the dietary sucrose modulation of enzyme activities in Drosophila melanogaster*
-
- Journal:
- Genetical Research / Volume 43 / Issue 3 / June 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 307-321
-
- Article
-
- You have access
- Export citation
