Refine search
Actions for selected content:
10186 results in Genetics
GRH volume 51 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 51 / Issue 2 / April 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Books received
-
- Journal:
- Genetical Research / Volume 19 / Issue 3 / June 1972
- Published online by Cambridge University Press:
- 14 April 2009, p. 320
-
- Article
-
- You have access
- Export citation
A test for the role of natural selection in the stabilization of transposable element copy number in a population of Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 49 / Issue 1 / February 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 31-41
-
- Article
-
- You have access
- Export citation
Cold Spring Harbor Symposia on Quantitative Biology. Volume L. Molecular Biology of Development. Cold Spring Harbor Laboratory. 1986. 920 pages. Cloth $140. ISBN 0 87969 050 X. Paper $70. ISBN 0 87969 051 8.
-
- Journal:
- Genetical Research / Volume 49 / Issue 2 / April 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 178-180
-
- Article
-
- You have access
- Export citation
Linkage relations and some pleiotropic effects of the dreher mutant of the house mouse
-
- Journal:
- Genetical Research / Volume 2 / Issue 1 / February 1961
- Published online by Cambridge University Press:
- 14 April 2009, pp. 92-95
-
- Article
-
- You have access
- Export citation
The probability of establishment of an advantageous mutant in a subdivided population
-
- Journal:
- Genetical Research / Volume 50 / Issue 1 / August 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 35-40
-
- Article
-
- You have access
- Export citation
Chromosomal location of a barley malt endopeptidase gene
-
- Journal:
- Genetical Research / Volume 59 / Issue 3 / June 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. 179-181
-
- Article
-
- You have access
- Export citation
Estimating gene flow in island populations
-
- Journal:
- Genetical Research / Volume 67 / Issue 2 / April 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 147-158
-
- Article
-
- You have access
- Export citation
Differences in allele frequencies of Aat between high- and mid-rocky shore populations of Littorina saxatilis (Olivi) suggest selection in this enzyme locus
-
- Journal:
- Genetical Research / Volume 54 / Issue 1 / August 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 7-12
-
- Article
-
- You have access
- Export citation
Natural selection as the process of accumulating genetic information in adaptive evolution*
-
- Journal:
- Genetical Research / Volume 2 / Issue 1 / February 1961
- Published online by Cambridge University Press:
- 14 April 2009, pp. 127-140
-
- Article
-
- You have access
- Export citation
-
1. In the course of evolution, complicated organisms have descended from much simpler ones. Since the instructions to form an organism are contained in the nucleus of its fertilized egg, this means that the genetic constitution has become correspondingly more complex in evolution. If we express this complexity in terms of its improbability, defining the amount of genetic information as the negative logarithm of its probability of occurrence by chance, we may say that genetic information is increased in the course of progressive evolution, guided by natural selection of random mutations.
2. It was demonstrated that the rate of accumulation of genetic information in adaptive evolution is directly proportional to the substitutional load, i.e. the decrease of Darwinian fitness brought about by substituting for one gene its allelic form which is more fitted to a new environment. The rate of accumulation of genetic information is given by
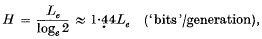
where Le is the substitutional load measured in ‘Malthusian parameters’.
3. Using Le = 0·199, a value obtained from the application of the ‘principle of minimum genetic load’ (cf. Kimura, 1960 b), we get
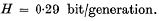
It was estimated that the total amount of genetic information accumulated since the beginning of the Cambrian epoch (500 million years) may be of the order of 108 bits, if evolution has proceeded at the standard rate.
Since the genetic information is transformed into phenotypic information in ontogeny, this figure (108 bits) must represent the amount of information which corresponds to the improved organization of higher animals as compared to their ancestors 500 million years back.
4. Problems involved in storage and transformation of genetic information thus acquired were discussed and it was pointed out that the redundancy of information in the form of repetition in linear sequence of nucleotide pairs within a gene may play an important role in the storage of genetic information.
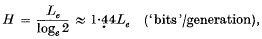
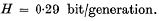
GRH volume 51 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 51 / Issue 2 / April 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
GRH volume 34 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 34 / Issue 1 / August 1979
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
GRH volume 47 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 47 / Issue 3 / June 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Genetic loads at a polymorphic locus which is maintained by frequency-dependent selection*
-
- Journal:
- Genetical Research / Volume 16 / Issue 2 / October 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 145-150
-
- Article
-
- You have access
- Export citation
A rapid and efficient replica plating technique
-
- Journal:
- Genetical Research / Volume 15 / Issue 3 / June 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 335-338
-
- Article
-
- You have access
- Export citation
Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 31 / Issue 3 / June 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. 309-311
-
- Article
-
- You have access
- Export citation
Genic basis of the mate-killer trait in Paramecium aurelia, stock 540
-
- Journal:
- Genetical Research / Volume 2 / Issue 1 / February 1961
- Published online by Cambridge University Press:
- 14 April 2009, pp. 82-91
-
- Article
-
- You have access
- Export citation
Selection at the alcohol dehydrogenase locus in Drosophila melanogaster imposed by environmental ethanol
-
- Journal:
- Genetical Research / Volume 26 / Issue 3 / December 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 265-274
-
- Article
-
- You have access
- Export citation
The maintenance of the genetic variability of polygenic characters by heterozygous advantage
-
- Journal:
- Genetical Research / Volume 22 / Issue 1 / August 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 9-12
-
- Article
-
- You have access
- Export citation
Banbury Report 14. Recombinant DNA: Applications to Human Disease. Edited by C. T. Caskey and R. L. White. Cold Spring Harbor Laboratory. 1983. About 375 pages. $55 U.S. ($66 outside U.S.) ISBN 0 87967 214 6.
-
- Journal:
- Genetical Research / Volume 43 / Issue 3 / June 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 346-347
-
- Article
-
- You have access
- Export citation
