Refine listing
Actions for selected content:
Contents
Research Article
Development, hatching and mortality of the eggs of Cimex lectularius L. (Hemiptera) in relation to climate, with observations on the effects of preconditioning to temperature
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 127-173
-
- Article
- Export citation
The longevity of the fasting bed-bug (C. lectularius L.) under experimental conditions and particularly in relation to the saturation deficiency law of water-loss
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 239-270
-
- Article
- Export citation
-
1. With unfed 1st instars the relation between mean length of life and saturation deficit at constant temperatures between 7 and 15° C. at relative humidities between 7 and 90% is hyperbolic. The relationship becomes more linear at higher temperatures.
At constant saturation deficits insects live longer at 15° C. than at lower temperatures. Longevity also decreases with rise of temperature above approximately 15° C.
2. In general, the longevity curves, except for those at temperatures below 15° C., bear a very similar relation to saturation deficit and to each other as the reciprocal curves for rate of water-loss at the different temperatures.
The influence of climatic factors on longevity at constant temperatures is discussed at length and it is concluded that over much of the temperature and humidity range survival time is limited by water-loss. At the higher humidities it is thought that either food, or perhaps an excessive accumulation of water within the insect, limits survival and causes a departure from the hyperbolic relation of longevity to saturation deficit.
3. The effects on longevity of a single meal are discussed. The principal effect of a blood meal is to increase the time of survival. But the factors which limit survival at different humidities appear to be the same as with unfed bugs, except at high humidities below about 15° C.
4.Mellanby's data on the rate of water-loss from fasting adult bed-bugs is analysed. It is found that the rate of water-loss is directly proportional to saturation deficit at constant temperatures between 8 and 37° C. and between 0 and 90% r.h. Although the rate may always be directly proportional to saturation deficit, the expression
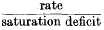 is not always constant.
is not always constant.Rate = K + b (saturation deficit), where K varies with temperature and b remains constant.
5. Longevity in relation to host blood is discussed. Rabbit blood appears to be slightly less favourable to survival than human blood.
6. If bugs are allowed to feed to repletion, longevity is not correlated with the size of the meal, nor with the weight of the unfed insect.
Virgin females live longer than mated ones, but no effects of mating on survival were noticed with males. Mated males tend to outlive mated females except at very low temperatures: virgin females live longer than unmated males.
7. The results of other workers and the possible causes of some discrepancies are discussed.
8. The maximum survival times of bugs are listed. Adults and 5th instars live longer than other stages. In a house which has remained empty for a long time it is probable that 5th instars and adults, particularly unmated female adults, would predominate in the population.
The longest observed survival was between 562 and 572 days.
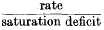 is not always constant.
is not always constant.The occurrence of Rickettsia-like bodies in the Reduviid bug Triatoma rubrofasciata and their transmission to laboratory animals
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 355-360
-
- Article
- Export citation
Notes on the preparation of Papers for publication in the Journal of Hygiene and in Parasitology1
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 1-62
-
- Article
- Export citation
Hexacotyle extensicauda n.sp., a monogenetic trematode from the gills of the tunny (Thunnus thynnus L.)
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 271-286
-
- Article
- Export citation
An investigation of the common fish louse, Argulus foliaceus (Linn.)
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 361-371
-
- Article
- Export citation
A new record of Plasmodium ovale (Stephens, 1922)
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 63-66
-
- Article
- Export citation
Some avian tapeworms from Antigua
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 174-197
-
- Article
- Export citation
Notes on the formation of the egg capsules in the monogenetic trematode, Hexacotyle extensicauda Dawes, 1940
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 287-295
-
- Article
- Export citation
Three closely related species of Aploparaksis Clerc, 1903
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 198-207
-
- Article
- Export citation
Studies on the germ cell cycle of the digenetic trematode Parorchis acanthus Nicoll: Part II. Structure of the miracidium and germinal development in the larval stages
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 372-391
-
- Article
- Export citation
Observations on the incidence of some nematode parasites of the common rabbit, Oryctolagus cuniculus
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 67-77
-
- Article
- Export citation
The effects of environmental conditions on the accessibility of third stage trichostrongyle larvae to grazing animals
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 208-225
-
- Article
- Export citation
Adelina schellacki n.sp., a coccidium from the intestine of the Indian centipede Cormocephalus dentipes poc.
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 392-396
-
- Article
- Export citation
Studies on populations of head-lice (Pediculus humanus capitis: Anoplura): III. Material from South India
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 296-302
-
- Article
- Export citation
Observations upon some common cestode parasites of the wild rabbit, Oryctolagus cuniculus
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 78-90
-
- Article
- Export citation
-
1. Between November 1936 and February 1937, ninety-six rabbits were examined, whilst between August 1937 and July 1938, 446 rabbits were examined for cestode parasites. The rabbits were obtained from the same area.
2. The rabbits were infected with the following cestodes to the extent given, for the two periods respectively:

3. Two species of the genus Hymenolepis are recorded for the first time from the rabbit, and they are briefly described.
4. The seasonal fluctuations in the incidence of the above cestodes are discussed.
5. The span of life of both Cittotaenia pectinata and C. denticulata is seen to be not greater than 10 months, and evidence is given to show that the intermediate host of C. denticulata is available between April and December, and that of C. pectinata from August to April.
6. Evidence of host and age resistance is demonstrated in Cysticercus pisiformis.

Some difficulties in determining the life cycle of Fasciola
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 397-398
-
- Article
- Export citation
On an Entamoeba occurring in English goats
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 226-237
-
- Article
- Export citation
Notes on some new species of parasitic mites Part 3
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 91-104
-
- Article
- Export citation
The biology of the body louse (Pediculus humanus corporis: Anoplura) under experimental conditions
-
- Published online by Cambridge University Press:
- 06 April 2009, pp. 303-312
-
- Article
- Export citation