Refine search
Actions for selected content:
10186 results in Genetics
Inheritance of a meiosis I error expressed in mouse oocytes and modulated by a maternal factor
-
- Journal:
- Genetical Research / Volume 49 / Issue 3 / June 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 239-243
-
- Article
-
- You have access
- Export citation
Location of genes controlling the D group of LMW glutenin subunits on chromosome 1D of bread wheat
-
- Journal:
- Genetical Research / Volume 47 / Issue 3 / June 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 175-179
-
- Article
-
- You have access
- Export citation
Isolation and characterization of deficiencies exposing the rudimentary locus of Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 37 / Issue 2 / April 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 199-207
-
- Article
-
- You have access
- Export citation
The transposons Tn501(Hg) and Tn1721(Tc) are related
-
- Journal:
- Genetical Research / Volume 37 / Issue 3 / June 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 285-289
-
- Article
-
- You have access
- Export citation
The control of body size in mouse chimaeras
-
- Journal:
- Genetical Research / Volume 38 / Issue 1 / August 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 25-46
-
- Article
-
- You have access
- Export citation
GRH volume 4 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 4 / Issue 2 / July 1963
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Further evidence for the importance of parental source of the Xce allele in X chromosome inactivation
-
- Journal:
- Genetical Research / Volume 58 / Issue 1 / August 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 63-65
-
- Article
-
- You have access
- Export citation
Genetic assimilation of part of a mutant phenotype
-
- Journal:
- Genetical Research / Volume 26 / Issue 2 / October 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 149-162
-
- Article
-
- You have access
- Export citation
Fungal Differentiation, A Contemporary Synthesis (Mycology Series, Volume 4). Edited by J. E. Smith. Basel: Dekker. 656 pages. 1983. £84.70. ISBN 0 8247 1734 1.
-
- Journal:
- Genetical Research / Volume 43 / Issue 3 / June 1984
- Published online by Cambridge University Press:
- 14 April 2009, p. 350
-
- Article
-
- You have access
- Export citation
Differences in P element population dynamics between the sibling species Drosophila melanogaster and Drosophila simulans
-
- Journal:
- Genetical Research / Volume 63 / Issue 1 / February 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 27-38
-
- Article
-
- You have access
- Export citation
Genetic analysis of axonemal mutants in Paramecium tetraurelia defective in their response to calcium
-
- Journal:
- Genetical Research / Volume 43 / Issue 1 / February 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 11-20
-
- Article
-
- You have access
- Export citation
Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 29 / Issue 3 / June 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 295-297
-
- Article
-
- You have access
- Export citation
Genetics and Alzheimer's Disease. Edited by P. M. Sinet, Y. Lamour and Y. Christen pp. 179Springer-Verlag, Berlin, Heidelberg and New York1988. Price DM 108
-
- Journal:
- Genetical Research / Volume 54 / Issue 2 / October 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 161-162
-
- Article
-
- You have access
- Export citation
On the theory of artificial selection in finite populations*
-
- Journal:
- Genetical Research / Volume 13 / Issue 2 / April 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 143-163
-
- Article
-
- You have access
- Export citation
Relationship between chromosome content and nuclear diameter in early spermatids of Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 54 / Issue 3 / December 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 205-212
-
- Article
-
- You have access
- Export citation
Molecular Genetics of Parasitic Protozoa; Current Communications in Molecular Biology. Edited by Mervyn J. Turner and David Arnot. New York: Cold Spring Harbor Laboratory. 1988. 204 pages, Paper $25.00. ISBN 0 87969 313 4.
-
- Journal:
- Genetical Research / Volume 53 / Issue 1 / February 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 72-73
-
- Article
-
- You have access
- Export citation
The genetics of tasting in mice: III. Quinine
-
- Journal:
- Genetical Research / Volume 44 / Issue 2 / October 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 151-160
-
- Article
-
- You have access
- Export citation
Response to selection from new mutation and effective size of partially inbred populations. I. Theoretical results
-
- Journal:
- Genetical Research / Volume 66 / Issue 3 / December 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 213-225
-
- Article
-
- You have access
- Export citation
Prediction of long-term contributions and inbreeding in populations undergoing mass selection
-
- Journal:
- Genetical Research / Volume 62 / Issue 3 / December 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. 231-242
-
- Article
-
- You have access
- Export citation
-
For a population undergoing mass selection, derived from an unselected base population in generation zero, the expected long-term contribution to the population of an ancestor from generation 1 was shown to be equal to that expected during random selection multiplied by
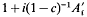 (where
(where 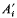 is one half the breeding value of the ancestor for the trait under selection standardized by the phenotypic standard deviation, i the intensity of selection, and
is one half the breeding value of the ancestor for the trait under selection standardized by the phenotypic standard deviation, i the intensity of selection, and 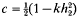 is the competitiveness which is defined by
is the competitiveness which is defined by 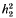 the heritability in generation 2 and k the variance reduction coefficient). It was shown that the rate of inbreeding (ΔF) could be partitioned into three components arising from expected contributions, sampling errors and sampling covariances respectively. Using this result ΔF was derived and shown to be dominated by terms that describe ΔF by variance of family size in a single generation plus a term that accounts for the expected proliferation of lines over generations from superior ancestors in generation 1. The basic prediction of ΔF was given by
the heritability in generation 2 and k the variance reduction coefficient). It was shown that the rate of inbreeding (ΔF) could be partitioned into three components arising from expected contributions, sampling errors and sampling covariances respectively. Using this result ΔF was derived and shown to be dominated by terms that describe ΔF by variance of family size in a single generation plus a term that accounts for the expected proliferation of lines over generations from superior ancestors in generation 1. The basic prediction of ΔF was given by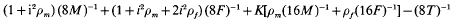
where M and F are the numbers of breeding males and females, T the number of offspring of each sex, ρm and ρt are correlations among half-sibs in generation 2 for males and females respectively, and K is a function of the intensity and competitiveness.
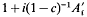 (where
(where 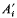 is one half the breeding value of the ancestor for the trait under selection standardized by the phenotypic standard deviation, i the intensity of selection, and
is one half the breeding value of the ancestor for the trait under selection standardized by the phenotypic standard deviation, i the intensity of selection, and 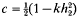 is the competitiveness which is defined by
is the competitiveness which is defined by 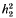 the heritability in generation 2 and k the variance reduction coefficient). It was shown that the rate of inbreeding (ΔF) could be partitioned into three components arising from expected contributions, sampling errors and sampling covariances respectively. Using this result ΔF was derived and shown to be dominated by terms that describe ΔF by variance of family size in a single generation plus a term that accounts for the expected proliferation of lines over generations from superior ancestors in generation 1. The basic prediction of ΔF was given by
the heritability in generation 2 and k the variance reduction coefficient). It was shown that the rate of inbreeding (ΔF) could be partitioned into three components arising from expected contributions, sampling errors and sampling covariances respectively. Using this result ΔF was derived and shown to be dominated by terms that describe ΔF by variance of family size in a single generation plus a term that accounts for the expected proliferation of lines over generations from superior ancestors in generation 1. The basic prediction of ΔF was given by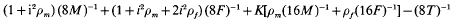
Physiological genetics of melanotic tumours in Drosophila melanogaster: VIII. The role of choline in the expression of the tumour gene tu bw and of its suppressor, su-tu
-
- Journal:
- Genetical Research / Volume 24 / Issue 2 / October 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 215-227
-
- Article
-
- You have access
- Export citation
