Refine search
Actions for selected content:
10186 results in Genetics
Genetics of lycopene cyclization and substrate transfer in β-carotene biosynthesis in Phycomyces
-
- Journal:
- Genetical Research / Volume 36 / Issue 3 / December 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 299-309
-
- Article
-
- You have access
- Export citation
Recombination and the evolution of satellite DNA
-
- Journal:
- Genetical Research / Volume 47 / Issue 3 / June 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 167-174
-
- Article
-
- You have access
- Export citation
Glucose-6-phosphate dehydrogenase expression in heterozygous kangaroo embryos and extra-embryonic membranes
-
- Journal:
- Genetical Research / Volume 45 / Issue 2 / April 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 205-208
-
- Article
-
- You have access
- Export citation
Isolation and properties of mutants of the FP2 sex factor of Pseudomonas aeruginosa
-
- Journal:
- Genetical Research / Volume 22 / Issue 1 / August 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 13-24
-
- Article
-
- You have access
- Export citation
International Industrial Biotechnology. Cambridge University Press. Vol. 8. 1988. 6 issues. £55 for Institutions, £32 for Individuals. ISSN 0269 7815.
-
- Journal:
- Genetical Research / Volume 52 / Issue 1 / August 1988
- Published online by Cambridge University Press:
- 14 April 2009, p. 78
-
- Article
-
- You have access
- Export citation
Selection for conditional lethals: A general negative selective system for Schizophyllum commune
-
- Journal:
- Genetical Research / Volume 29 / Issue 1 / February 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 93-96
-
- Article
-
- You have access
- Export citation
Molecular Biology of the Photosynthetic Apparatus. Edited by K. E. Steinback, S. Bonitz, C. J. Arntzen and L. Bogorad. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory. $75. ISBN 0 87969 188 3.
-
- Journal:
- Genetical Research / Volume 48 / Issue 1 / August 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 58-59
-
- Article
-
- You have access
- Export citation
Spontaneous mutation for a quantitative trait in Drosophila melanogaster. I. Response to artificial selection
-
- Journal:
- Genetical Research / Volume 61 / Issue 2 / April 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. 107-116
-
- Article
-
- You have access
- Export citation
Man Made Life, A Genetic Engineering Primer, By Jeremy Cherfas. Oxford: Basil Blackwell (1982). 270 pages, price £5.50.
-
- Journal:
- Genetical Research / Volume 41 / Issue 1 / February 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 106-107
-
- Article
-
- You have access
- Export citation
Heterogeneity in the Sxr (sex-reversal) locus of the mouse as revealed by synthetic
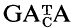 probes
probes
-
- Journal:
- Genetical Research / Volume 51 / Issue 3 / June 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 239-246
-
- Article
-
- You have access
- Export citation
-
The sex-reversal mutation, Sxr and a variant form, Sxr′ have been established on the inbred C57BL/6Mcl background by repeated backcrossing to form the CB and CB′ strains, respectively. DNAs of normal XY, XX Sxr and XX Sxr′ as well as XY Sxr and XY Sxr′ carrier male mice have been digested with the restriction enzymes Hae III and Hinf I and electrophoresed. The DNAs show many common but also differing hybridization bands with synthetic
oligonucleotide probes. In XY Sxr (and XY Sxr′) carrier males, the hybridization patterns of normal XY and those of XX Sxr (and XX Sxr′) males are simply superimposed. Individual differing bands can be categorized by their differential hybridization behaviour to the (GATA)4, (GACA)4, (GATA)2 GACA (GATA)2 and (GATA)3 (GACA)2 probes. In general, the hybridization patterns are regularly inherited. In addition to the predominant pattern in each strain, one additional XX Sxr and one additional XX Sxr′ hybridization pattern was observed: the additional pattern in the CB strain was transmitted (via variant XY Sxr carriers) while the secondary XX Sxr′ pattern in the CB′ strain could only be observed once. Thus ‘DNA finger printing’ with oligonucleotide probes can successfully be used to discriminate the DNAs of normal Y chromosomes, XX Sxr and XX Sxr′ variants as well as XY Sxr and XY Sxr′ carrier mice. Implications of the comparatively high unequal recombination rate involving the murine Y chromosome are discussed, as well as possible mechanisms.
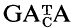 probes
probes
The functions of the phage T4 immunity and spackle genes in genetic exclusion
-
- Journal:
- Genetical Research / Volume 52 / Issue 2 / October 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 81-90
-
- Article
-
- You have access
- Export citation
Mitochondrial Genes. Edited by Piotr Slonimski, Piet Borst and Giuseppe Attardi. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1982.
-
- Journal:
- Genetical Research / Volume 42 / Issue 2 / October 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 229-230
-
- Article
-
- You have access
- Export citation
Deletion analysis of male sterility effects of t–haplotypes in the mouse
-
- Journal:
- Genetical Research / Volume 56 / Issue 2-3 / October 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 179-183
-
- Article
-
- You have access
- Export citation
GRH volume 56 issue 2-3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 56 / Issue 2-3 / October 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Segregation variance after hybridization of isolated populations
-
- Journal:
- Genetical Research / Volume 64 / Issue 1 / August 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 51-56
-
- Article
-
- You have access
- Export citation
GRH volume 4 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 4 / Issue 1 / February 1963
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f6
-
- Article
-
- You have access
- Export citation
GRH volume 17 issue 3 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 17 / Issue 3 / June 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b5
-
- Article
-
- You have access
- Export citation
Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 16 / Issue 3 / December 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 369-371
-
- Article
-
- You have access
- Export citation
GRH volume 61 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 61 / Issue 2 / April 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Observations on a set of radiation-induced dominant T-like mutations in the mouse*
-
- Journal:
- Genetical Research / Volume 26 / Issue 1 / August 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 95-108
-
- Article
-
- You have access
- Export citation
