Refine search
Actions for selected content:
10186 results in Genetics
Maternal effects on body weight in mice selected for large and small size
-
- Journal:
- Genetical Research / Volume 32 / Issue 3 / November 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. 295-302
-
- Article
-
- You have access
- Export citation
GRH volume 60 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 60 / Issue 3 / December 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Inversion polymorphism in a two-locus genetic system
-
- Journal:
- Genetical Research / Volume 23 / Issue 3 / June 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 259-280
-
- Article
-
- You have access
- Export citation
Changes in genetic constitution and sterol composition during growth of nystatin-resistant heterokaryons of Neurospora crassa
-
- Journal:
- Genetical Research / Volume 42 / Issue 1 / August 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 91-103
-
- Article
-
- You have access
- Export citation
GRH volume 9 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 9 / Issue 2 / April 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f5
-
- Article
-
- You have access
- Export citation
GRH volume 39 issue 3 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 39 / Issue 3 / June 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
Population genetics of extranuclear genomes under the neutral mutation hypothesis*
-
- Journal:
- Genetical Research / Volume 42 / Issue 3 / December 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 235-255
-
- Article
-
- You have access
- Export citation
Two-way within-family and mass selection for 8-week body weight in different mouse populations
-
- Journal:
- Genetical Research / Volume 43 / Issue 2 / April 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 191-200
-
- Article
-
- You have access
- Export citation
Adaptation to environmental heterogeneity in populations of Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 49 / Issue 1 / February 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-10
-
- Article
-
- You have access
- Export citation
An approach to the problem of whether clustering of functionally related genes occurs in higher organisms*
-
- Journal:
- Genetical Research / Volume 9 / Issue 2 / April 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 141-147
-
- Article
-
- You have access
- Export citation
Detrimental genes with partial selfing and effects on a neutral locus*
-
- Journal:
- Genetical Research / Volume 23 / Issue 2 / April 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 191-200
-
- Article
-
- You have access
- Export citation
Plant Infectious Agents. By Hugh D. Robertson, Stephen H. Howell, Milton Zaitlin and Russel L. Malmberg. Cold Spring Harbor Laboratory, P.O. Box 100, Cold Spring Harbor, New York 11724. 200 pp. (approx.). Paperback $23 ($27·60 outside U.S.) ISBN 0 87969 159 X.
-
- Journal:
- Genetical Research / Volume 43 / Issue 1 / February 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 101-102
-
- Article
-
- You have access
- Export citation
Infection into paramecia of metagons derived from other mate-killer paramecia
-
- Journal:
- Genetical Research / Volume 5 / Issue 1 / February 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 85-106
-
- Article
-
- You have access
- Export citation
Polymorphism of plasma esterases in flounder and plaice
-
- Journal:
- Genetical Research / Volume 11 / Issue 2 / April 1968
- Published online by Cambridge University Press:
- 14 April 2009, pp. 179-182
-
- Article
-
- You have access
- Export citation
Map and function of gad mutations in Physarum polycephalum*
-
- Journal:
- Genetical Research / Volume 42 / Issue 1 / August 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 41-57
-
- Article
-
- You have access
- Export citation
Unusual behaviour of linkage disequilibrium in two-locus gene conversion models
-
- Journal:
- Genetical Research / Volume 51 / Issue 1 / February 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 55-58
-
- Article
-
- You have access
- Export citation
-
The amount of linkage of disequilibrium maintained in a two-locus infinite population model by gene conversion and recombination is examined. Intrachromosomal conversion (conversion between different loci on the same chromosome) generates positive linkage disequilibrium. Specifically,
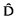 = p(1 − p) [1 − r/(γ + r − γr)], where p is the frequency of allele A at both loci, r is the recombination rate between loci and γ is the per-gamete conversion rate. Somewhat unexpectedly, interchromosomal conversion (conversion between loci on different chromosomes) also generates positive disequilibrium, albeit very small. More interestingly, the behaviour of this disequilibrium as a function of recombination is unusual. If β is the interchromosomal conversion rate between a pair of loci, then = p(1 − p) [rβ/(β+r − βr)]. increases with increasing recombination, being zero for the case of complete linkage (r = 0), and maximized at r = 1/2. This unusual behaviour can be accounted for by the generation of excess coupling gametes when an interchromosomal conversion event is followed by recombination.
= p(1 − p) [1 − r/(γ + r − γr)], where p is the frequency of allele A at both loci, r is the recombination rate between loci and γ is the per-gamete conversion rate. Somewhat unexpectedly, interchromosomal conversion (conversion between loci on different chromosomes) also generates positive disequilibrium, albeit very small. More interestingly, the behaviour of this disequilibrium as a function of recombination is unusual. If β is the interchromosomal conversion rate between a pair of loci, then = p(1 − p) [rβ/(β+r − βr)]. increases with increasing recombination, being zero for the case of complete linkage (r = 0), and maximized at r = 1/2. This unusual behaviour can be accounted for by the generation of excess coupling gametes when an interchromosomal conversion event is followed by recombination.
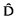 = p(1 − p) [1 − r/(γ + r − γr)], where p is the frequency of allele A at both loci, r is the recombination rate between loci and γ is the per-gamete conversion rate. Somewhat unexpectedly, interchromosomal conversion (conversion between loci on different chromosomes) also generates positive disequilibrium, albeit very small. More interestingly, the behaviour of this disequilibrium as a function of recombination is unusual. If β is the interchromosomal conversion rate between a pair of loci, then
= p(1 − p) [1 − r/(γ + r − γr)], where p is the frequency of allele A at both loci, r is the recombination rate between loci and γ is the per-gamete conversion rate. Somewhat unexpectedly, interchromosomal conversion (conversion between loci on different chromosomes) also generates positive disequilibrium, albeit very small. More interestingly, the behaviour of this disequilibrium as a function of recombination is unusual. If β is the interchromosomal conversion rate between a pair of loci, then Teratocarcinomas and Embryonic Stem Cells: A Practical Approach. Edited by E. J. Robertson. Oxford: IRL Press, Practical Approach Series. 1987. xii + 268 pages. Subject index. Softbound £17.00/$US 32.00 (hardcover £26.00/$US 47.00).
-
- Journal:
- Genetical Research / Volume 52 / Issue 2 / October 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 158-159
-
- Article
-
- You have access
- Export citation
Mapping the adrenal lipid depletion gene of the AKR/J mouse strain
-
- Journal:
- Genetical Research / Volume 26 / Issue 3 / December 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 307-312
-
- Article
-
- You have access
- Export citation
A structural gene (Tcp-1) within the mouse t complex is separable from effects on tail length and lethality but may be associated with effects on spermatogenesis
-
- Journal:
- Genetical Research / Volume 38 / Issue 2 / October 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 115-123
-
- Article
-
- You have access
- Export citation
Conditional induction of λ prophage in exrA mutants of Escherichia coli
-
- Journal:
- Genetical Research / Volume 17 / Issue 2 / April 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 161-163
-
- Article
-
- You have access
- Export citation
