Refine search
Actions for selected content:
10186 results in Genetics
Differences in or near the responder region of complete and partial mouse t-haplotypes
-
- Journal:
- Genetical Research / Volume 50 / Issue 1 / August 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 29-34
-
- Article
-
- You have access
- Export citation
Genetic polymorphism and interspecific competitive ability in Drosophila
-
- Journal:
- Genetical Research / Volume 14 / Issue 2 / October 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 95-102
-
- Article
-
- You have access
- Export citation
GRH volume 3 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 3 / Issue 1 / February 1962
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Genetic differences between two substrains of the inbred 101 mouse strain
-
- Journal:
- Genetical Research / Volume 44 / Issue 3 / December 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 343-346
-
- Article
-
- You have access
- Export citation
Genetic analysis of actidione-resistant mutants in the Myxomycete Physarum polycephalum, Schw.
-
- Journal:
- Genetical Research / Volume 8 / Issue 1 / August 1966
- Published online by Cambridge University Press:
- 14 April 2009, pp. 101-110
-
- Article
-
- You have access
- Export citation
The genetic sensitivity to X-rays of mouse foetal gonads
-
- Journal:
- Genetical Research / Volume 1 / Issue 3 / November 1960
- Published online by Cambridge University Press:
- 14 April 2009, pp. 351-355
-
- Article
-
- You have access
- Export citation
The estimation of the number of mutationally silent loci in saturation-mapping experiments
-
- Journal:
- Genetical Research / Volume 35 / Issue 1 / February 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 33-44
-
- Article
-
- You have access
- Export citation
Plant Breeding Methodology. By Neal F. Jensen. Chichester, West Sussex UK: John Wiley & Sons. 1989. 676 pages £39.30. ISBN 0471 60190 X.
-
- Journal:
- Genetical Research / Volume 55 / Issue 1 / February 1990
- Published online by Cambridge University Press:
- 14 April 2009, p. 68
-
- Article
-
- You have access
- Export citation
Quantitative predictions of random segregation models of the ciliate macronucleus
-
- Journal:
- Genetical Research / Volume 27 / Issue 2 / April 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 227-238
-
- Article
-
- You have access
- Export citation
The fitness consequences of P element insertion in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 52 / Issue 1 / August 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 17-26
-
- Article
-
- You have access
- Export citation
GRH volume 62 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 62 / Issue 1 / August 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Models of long-term artificial selection in finite population with recurrent mutation
-
- Journal:
- Genetical Research / Volume 48 / Issue 2 / October 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 125-131
-
- Article
-
- You have access
- Export citation
-
The effects of mutation on mean and variance of response to selection for quantitative traits are investigated. The mutants are assumed to be unlinked, to be additive, and to have their effects symmetrically distributed about zero, with absolute values of effects having a gamma distribution. It is shown that the ratio
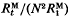 of expected cumulative response to generation t from mutants,
of expected cumulative response to generation t from mutants, 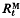 , and expected response over one generation from one generation of mutants,
, and expected response over one generation from one generation of mutants, 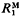 , is a function of t/N, where t is generations and N is effective population size. Similarly,
, is a function of t/N, where t is generations and N is effective population size. Similarly, 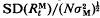 , is a function of t/N, where
, is a function of t/N, where 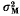 is the increment in genetic variance from one generation of mutants. The mean and standard deviation of response from mutations relative to that from initial variation in the population,
is the increment in genetic variance from one generation of mutants. The mean and standard deviation of response from mutations relative to that from initial variation in the population, 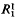 in the first generation, are functions of
in the first generation, are functions of 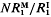 . Evaluation of these formulae for a range of parameters quantifies the important role that population size can play in response to long-term selection.
. Evaluation of these formulae for a range of parameters quantifies the important role that population size can play in response to long-term selection.
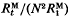 of expected cumulative response to generation t from mutants,
of expected cumulative response to generation t from mutants, 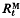 , and expected response over one generation from one generation of mutants,
, and expected response over one generation from one generation of mutants, 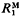 , is a function of t/N, where t is generations and N is effective population size. Similarly,
, is a function of t/N, where t is generations and N is effective population size. Similarly, 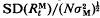 , is a function of t/N, where
, is a function of t/N, where 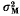 is the increment in genetic variance from one generation of mutants. The mean and standard deviation of response from mutations relative to that from initial variation in the population,
is the increment in genetic variance from one generation of mutants. The mean and standard deviation of response from mutations relative to that from initial variation in the population, 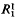 in the first generation, are functions of
in the first generation, are functions of 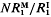 . Evaluation of these formulae for a range of parameters quantifies the important role that population size can play in response to long-term selection.
. Evaluation of these formulae for a range of parameters quantifies the important role that population size can play in response to long-term selection.Errata
-
- Journal:
- Genetical Research / Volume 50 / Issue 3 / December 1987
- Published online by Cambridge University Press:
- 14 April 2009, p. 251
-
- Article
-
- You have access
- Export citation
-
Genet. Res., Camb. (1987), 50, pp. 17–21
Heterogeneity of human haptoglobin α chains detected by two-dimensional gel electrophoresis
The running title for this article should have been Human plasma haptoglobin α chain heterogeneity
Genet. Res., Camb. (1987), 50, pp. 63–68
Additive variance and average effect with partial selfing
(1) The second line of Table 1 should read


GRH volume 39 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 39 / Issue 2 / April 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Spirochaete-mediated abnormal sex-ratio (SR) condition in Drosophila: A second virus associated with spirochaetes and its use in the study of the SR condition
-
- Journal:
- Genetical Research / Volume 18 / Issue 1 / August 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 45-56
-
- Article
-
- You have access
- Export citation
The parental origin of de novo X-autosome translocations in females with Duchenne muscular dystrophy revealed by M27ß methylation analysis
-
- Journal:
- Genetical Research / Volume 56 / Issue 2-3 / October 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 135-140
-
- Article
-
- You have access
- Export citation
Mechanisms of Molecular Evolution. Edited by N. Takahata and A. G. Clark. Japan Scientific Societies Press, Tokyo and Sinauer Associates, Inc., Sunderland, Mass. 1993. 250 pages. Price not available. ISBN 4 7622 6718 X/0 87893 825 7.
-
- Journal:
- Genetical Research / Volume 62 / Issue 3 / December 1993
- Published online by Cambridge University Press:
- 14 April 2009, p. 244
-
- Article
-
- You have access
- Export citation
GRH volume 48 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 48 / Issue 2 / October 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Genetic differences between populations of Drosophila melanogaster for a quantitative trait: I. Laboratory populations
-
- Journal:
- Genetical Research / Volume 22 / Issue 1 / August 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 51-68
-
- Article
-
- You have access
- Export citation
Clonal analysis of the lac operons from Klebsiella M5a1 and the Lac plasmid (pRE2) from Klebsiella V9A
-
- Journal:
- Genetical Research / Volume 54 / Issue 2 / October 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 85-91
-
- Article
-
- You have access
- Export citation
