Refine search
Actions for selected content:
10186 results in Genetics
A mathematical model of extranuclear genes and the genetic variability maintained in a finite population*
-
- Journal:
- Genetical Research / Volume 37 / Issue 3 / June 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 291-302
-
- Article
-
- You have access
- Export citation
-
A mathematical theory of population genetics accounting for the genes transmitted through mitochondria or chloroplasts has been studied. In the model it is assumed that a population consists of Nm males and Nf females, the genetic contribution from a male is β and that from a female 1 – β, and each cell line contains n effective copies of a gene in its cytoplasm. Assuming selective neutrality and an infinite alleles model, it is shown that the sum (H) of squares of allelic frequencies within an individual and the corresponding sum (Q) for the entire population are, at equilibrium, given by
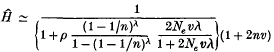
and
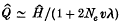
where ρ = 2β(1−β), Ne = {β2/Nm+(1−β)2 / Nf}−1, λ is the number of somatic cell divisions in one generation, and v is the mutation rate per cell division. If the genes are transmitted entirely through the female the formulae reduce to Ĥ ≃ 1/(1 + 2nv) and Q^ ≃ 1/{1 + (2Neλ+ 2n)υ}. Non-equilibrium behaviours of H and Q^ are also studied in the case of a panmictic population. These results are extended to geographically structured models, and applied to existing experimental data.
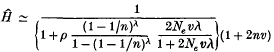
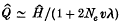
The negative correlation between somatic aneuploidy and growth in the oyster Crassostrea gigas and implications for the effects of induced polyploidization
-
- Journal:
- Genetical Research / Volume 68 / Issue 2 / October 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 109-116
-
- Article
-
- You have access
- Export citation
Abstracts of papers presented at the Mammalian Genetics Group meeting, held at the Royal Free Hospital Medical School on 26 and 27 November 1981
-
- Journal:
- Genetical Research / Volume 40 / Issue 1 / August 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 99-106
-
- Article
-
- You have access
- Export citation
GRH volume 19 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 19 / Issue 1 / February 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b5
-
- Article
-
- You have access
- Export citation
Thirteen genes (Cebpb, E2f1, Tcf4, Cyp24, Pck1, Acra4, Edn3, Kcnb1, Mc3r, Ntsr, Cd40, Plcg1 and Rcad) that probably lie in the distal imprinting region of mouse Chromosome 2 are not monoallelically expressed
-
- Journal:
- Genetical Research / Volume 65 / Issue 2 / April 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 83-93
-
- Article
-
- You have access
- Export citation
Genetic control of the kinetics of mouse spermatogenesis
-
- Journal:
- Genetical Research / Volume 22 / Issue 2 / October 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 155-167
-
- Article
-
- You have access
- Export citation
Mutants affecting amino acid cross-pathway control in Neurospora crassa
-
- Journal:
- Genetical Research / Volume 39 / Issue 2 / April 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 169-185
-
- Article
-
- You have access
- Export citation
Additive and non-additive genetic variance in odontometric traits in crosses of seven inbred lines of house mice
-
- Journal:
- Genetical Research / Volume 23 / Issue 2 / April 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 207-217
-
- Article
-
- You have access
- Export citation
Human Genetics: Problems and Approaches. Second edition. By F. Vogel and A. G. Motulsky. Berlin, Heidelberg, New York, Tokyo: Springer-Verlag1986, 807 pages. Hard cover DM 148, £52.25. ISBN 3 540 16411 1.
-
- Journal:
- Genetical Research / Volume 50 / Issue 2 / October 1987
- Published online by Cambridge University Press:
- 14 April 2009, p. 167
-
- Article
-
- You have access
- Export citation
GRH volume 16 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 16 / Issue 1 / August 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b3
-
- Article
-
- You have access
- Export citation
Selection for a threshold character in Drosophila: III. Genetic control of variability in plateaued populations*
-
- Journal:
- Genetical Research / Volume 15 / Issue 3 / June 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 285-300
-
- Article
-
- You have access
- Export citation
Cellular damage associated with multiple mating in E. coli
-
- Journal:
- Genetical Research / Volume 4 / Issue 3 / November 1963
- Published online by Cambridge University Press:
- 14 April 2009, pp. 463-469
-
- Article
-
- You have access
- Export citation
Parental age and recombination frequency in the house mouse
-
- Journal:
- Genetical Research / Volume 28 / Issue 3 / December 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 241-251
-
- Article
-
- You have access
- Export citation
The reduction in fixation probability caused by substitutions at linked loci
-
- Journal:
- Genetical Research / Volume 64 / Issue 3 / December 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 199-208
-
- Article
-
- You have access
- Export citation
-
The probability of fixation of a mutation with selective advantage s will be reduced by substitutions at other loci. The effect of a single substitution, with selective advantage
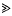 , can be approximated as a sudden reduction in the frequency of the favourable allele, by a fraction w = 1 −(s/S)r/s (where r is the recombination rate). An expression for the effect of a given sequence of such catastrophes is derived. This also applies to the ecological prxoblem of finding the probability that a small population will survive, despite occasional disasters. It is shown that if substitutions occur at a rate Δ, and are scattered randomly over a genetic map of length R, then an allele is unlikely to be fixed if its advantage is less than a critical value, Scrit = (π2/6)(2ΔS/(Rlog(S/s))). This threshold depends primarily on the variance in fitness per unit map length dueto substitutions, var(W)/R = 2ΔS/R. With no recombination, the fixation probability can be calculated for a finite population. If Δ > s, it is of the same order as for a neutral allele ( ≈ Δ/(2N(Δ−s))), whilst if
, can be approximated as a sudden reduction in the frequency of the favourable allele, by a fraction w = 1 −(s/S)r/s (where r is the recombination rate). An expression for the effect of a given sequence of such catastrophes is derived. This also applies to the ecological prxoblem of finding the probability that a small population will survive, despite occasional disasters. It is shown that if substitutions occur at a rate Δ, and are scattered randomly over a genetic map of length R, then an allele is unlikely to be fixed if its advantage is less than a critical value, Scrit = (π2/6)(2ΔS/(Rlog(S/s))). This threshold depends primarily on the variance in fitness per unit map length dueto substitutions, var(W)/R = 2ΔS/R. With no recombination, the fixation probability can be calculated for a finite population. If Δ > s, it is of the same order as for a neutral allele ( ≈ Δ/(2N(Δ−s))), whilst if 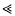 , fixation probability is much higher than for a neutral allele, but much lower than in the absence of hitch-hiking
, fixation probability is much higher than for a neutral allele, but much lower than in the absence of hitch-hiking 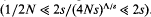 . These results suggest that hitch-hiking may substantially impede the accumulation of weakly favoured adaptations.
. These results suggest that hitch-hiking may substantially impede the accumulation of weakly favoured adaptations.
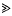 , can be approximated as a sudden reduction in the frequency of the favourable allele, by a fraction w = 1 −(s/S)
, can be approximated as a sudden reduction in the frequency of the favourable allele, by a fraction w = 1 −(s/S)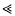 , fixation probability is much higher than for a neutral allele, but much lower than in the absence of hitch-hiking
, fixation probability is much higher than for a neutral allele, but much lower than in the absence of hitch-hiking 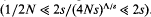 . These results suggest that hitch-hiking may substantially impede the accumulation of weakly favoured adaptations.
. These results suggest that hitch-hiking may substantially impede the accumulation of weakly favoured adaptations.In situ Hybridization. A Practical Approach. Edited by D.G. Wilkinson. IRL Press, Oxford. 1992. 163 pages. £18.50 Softback. ISBN 0 19 963327 4.
-
- Journal:
- Genetical Research / Volume 61 / Issue 3 / June 1993
- Published online by Cambridge University Press:
- 14 April 2009, p. 234
-
- Article
-
- You have access
- Export citation
Chromosomal change and rectangular evolution in North American cyprinid fishes*
-
- Journal:
- Genetical Research / Volume 35 / Issue 2 / April 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 157-164
-
- Article
-
- You have access
- Export citation
Multiple recombinational events within the 84 W locus of Ascorolus immersus
-
- Journal:
- Genetical Research / Volume 18 / Issue 2 / October 1971
- Published online by Cambridge University Press:
- 14 April 2009, pp. 199-214
-
- Article
-
- You have access
- Export citation
Selection against harmful mutations in large sexual and asexual populations
-
- Journal:
- Genetical Research / Volume 40 / Issue 3 / December 1982
- Published online by Cambridge University Press:
- 14 April 2009, pp. 325-332
-
- Article
-
- You have access
- Export citation
Drosophila genes cut and miniature are associated with the susceptibility to infection by Serratia marcescens
-
- Journal:
- Genetical Research / Volume 52 / Issue 1 / August 1988
- Published online by Cambridge University Press:
- 14 April 2009, pp. 51-56
-
- Article
-
- You have access
- Export citation
Genes. By B. Lewin. Chichester: John Wiley. 1984. xiii + 715 pages. Cloth £27.85. Paperback £13.95 ISBN 0 471093165
-
- Journal:
- Genetical Research / Volume 43 / Issue 2 / April 1984
- Published online by Cambridge University Press:
- 14 April 2009, pp. 221-222
-
- Article
-
- You have access
- Export citation
