Refine search
Actions for selected content:
11977 results in Plant sciences

CUC/auxin patterning of decanalised petal number in Cardamine hirsuta
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 01 July 2025, e29
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

An integrative process-based model of fruit growth as a function of carbon and water fluxes modulated by endogenous abscisic acid in blueberry fruit
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 30 June 2025, e19
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Chloride transport and homeostasis in plants
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 30 June 2025, e20
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Manganese handling in plants: Advances in the mechanistic and functional understanding of transport pathways
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 20 June 2025, e16
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Molecular tipping points in plant cell surface H+ homeostasis and signalling
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 18 June 2025, e28
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Four-dimensional phenotyping reveals MYOSIN XI-dependent establishment of branch morphology through upward- and stably-directed growth in Arabidopsis
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 10 June 2025, e15
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
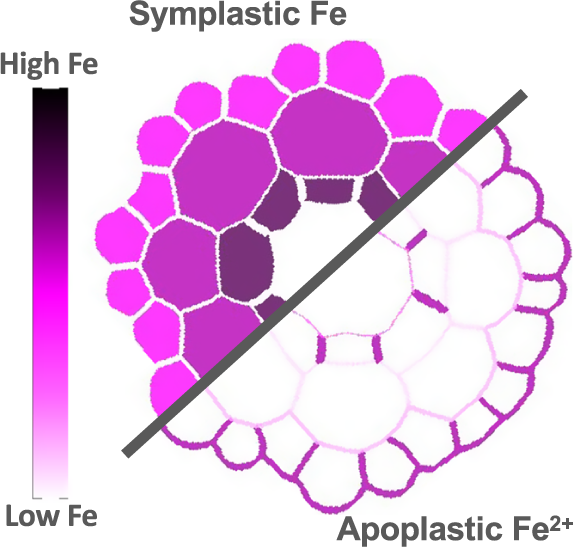
Untangling iron threads: A deep dive into plant intracellular pools
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 09 June 2025, e14
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Potassium homeostasis and signalling: from the whole plant to the subcellular level
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 08 May 2025, e13
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Wood formation of drought-resistant Eucalyptus cladocalyx under cyclical drought treatment
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 08 April 2025, e12
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

To flow or to grow? Impacts of tapping on sugar maple
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 07 April 2025, e11
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Changing paradigms for the micronutrient zinc, a known protein cofactor, as a signal relaying also cellular redox state
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 02 April 2025, e7
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Colour pattern studies: the SE (せ) method, a shape-centred approach to explore biodiversity and avoid aesthetic biases
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 02 April 2025, e10
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

A 3D morpho-space of sepal geometry reveals the importance of organ curvature
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 27 March 2025, e9
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Dynamics of homeostats: the basis of electrical, chemical, hydraulic, pH and calcium signaling in plants
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 21 March 2025, e8
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Synthetic gene circuits in plants: recent advances and challenges
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 27 February 2025, e6
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
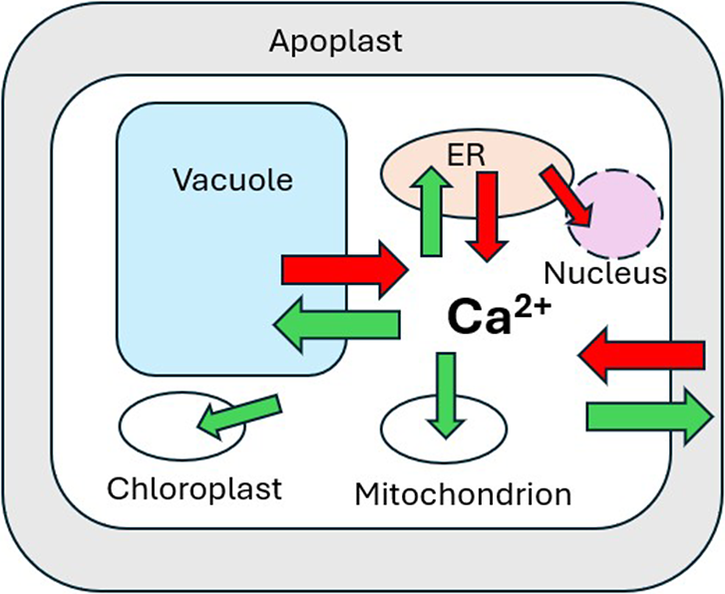
Cellular calcium homeostasis and regulation of its dynamic perturbation
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 14 February 2025, e5
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Why participatory plant research now?
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 13 February 2025, e4
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
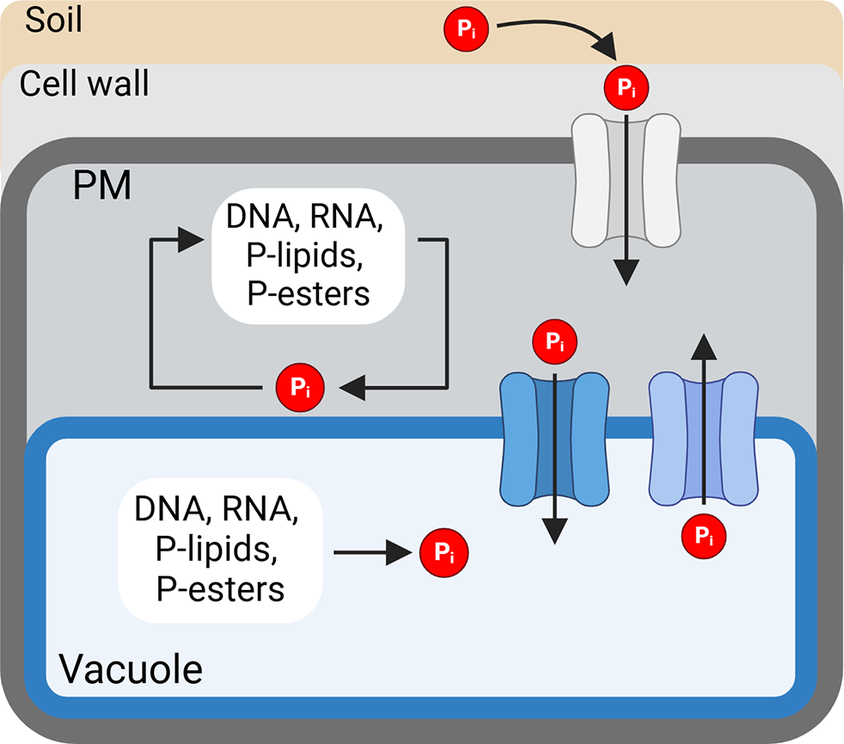
Mobilization and recycling of intracellular phosphorus in response to availability
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 30 January 2025, e3
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Microtubule flexibility, microtubule-based nucleation and ROP pattern co-alignment enhance protoxylem microtubule patterning
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 27 January 2025, e2
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

The Turing heritage for plant biology: all spots and stripes?
-
- Journal:
- Quantitative Plant Biology / Volume 6 / 2025
- Published online by Cambridge University Press:
- 13 January 2025, e1
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
