Refine search
Actions for selected content:
11975 results in Plant sciences
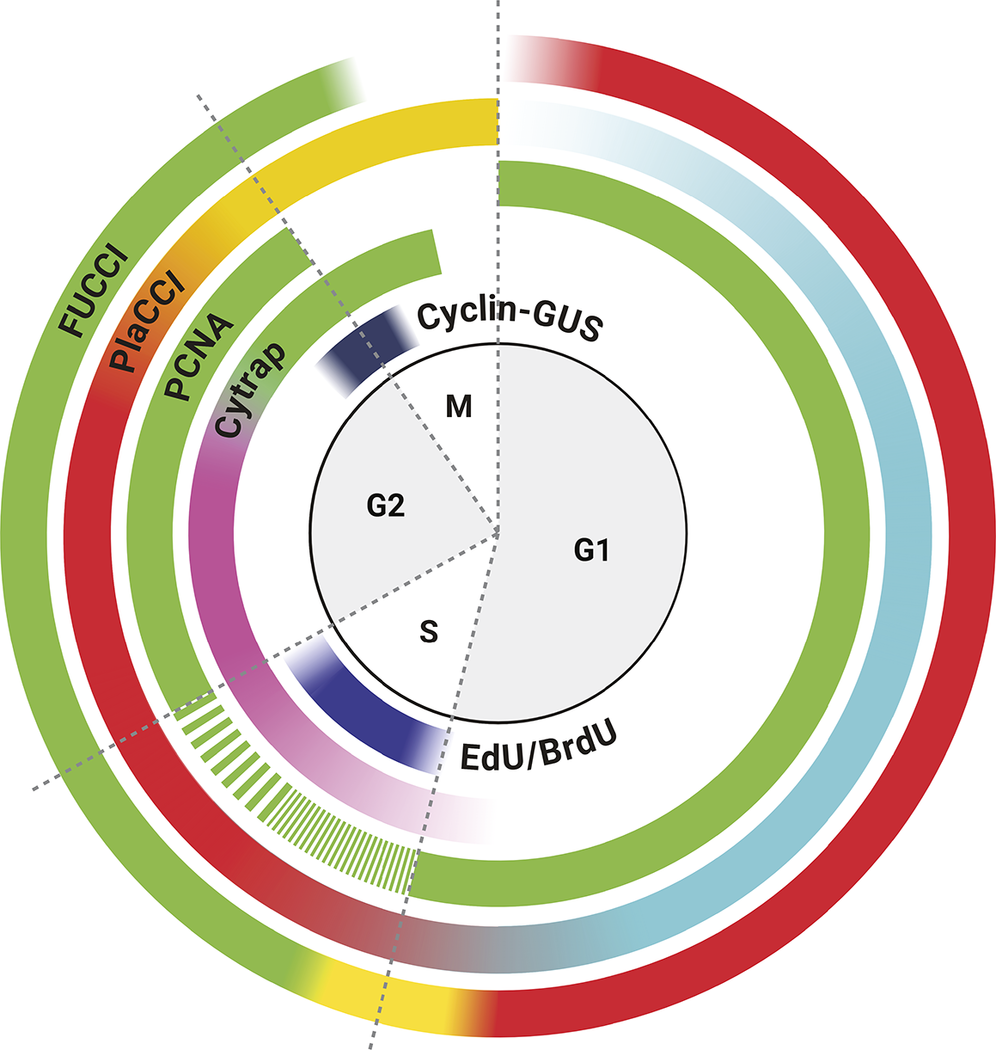
Molecular markers in cell cycle visualisation during development and stress conditions in Arabidopsis thaliana
- Part of
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 12 December 2024, e14
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
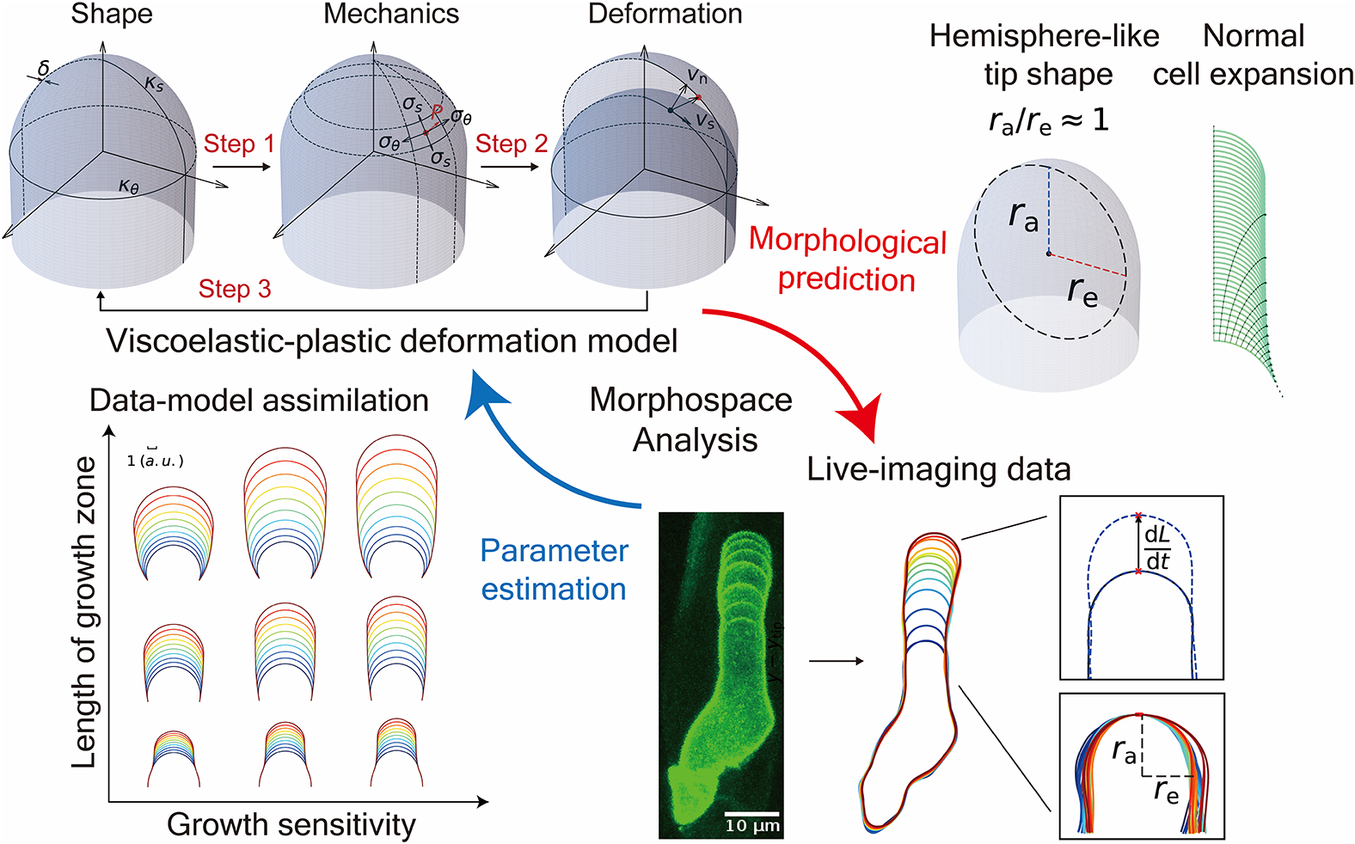
A viscoelastic–plastic deformation model of hemisphere-like tip growth in Arabidopsis zygotes
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 12 December 2024, e13
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
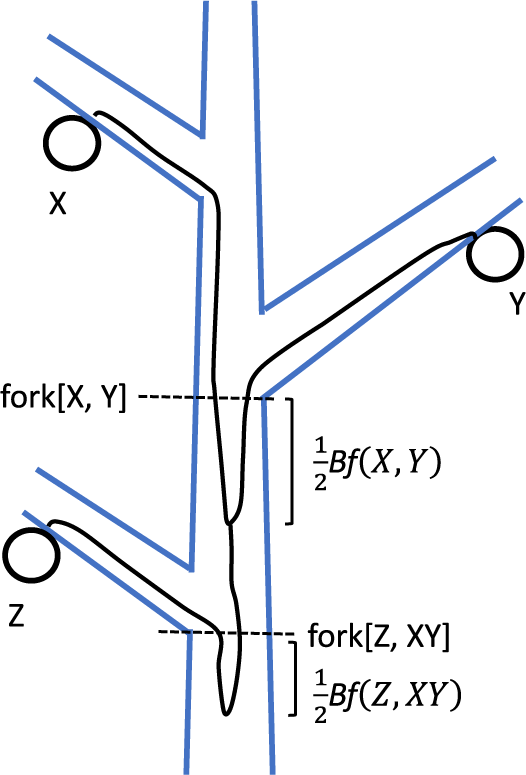
Genetic diversity within a tree and alternative indexes for different evolutionary effects
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 06 December 2024, e11
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Trees, living for centuries, accumulate somatic mutations in their growing trunks and branches, causing genetic divergence within a single tree. Stem cell lineages in a shoot apical meristem accumulate mutations independently and diverge from each other. In plants, somatic mutations can alter the genetic composition of reproductive organs and gametes, impacting future generations. To evaluate the genetic variation among a tree’s reproductive organs, we consider three indexes: mean pairwise phylogenetic distance (
 $\overline{D}$), phylogenetic diversity (
$\overline{D}$), phylogenetic diversity (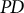 $PD$; sum of branch lengths in molecular phylogeny) and parent-offspring phylogenetic distance (
$PD$; sum of branch lengths in molecular phylogeny) and parent-offspring phylogenetic distance (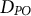 ${D}_{PO}$). The tissue architecture of trees facilitated the accumulation of somatic mutations, which have various evolutionary effects, including enhancing fitness under strong sib competition and intense host-pathogen interactions, efficiently eliminating deleterious mutations through epistasis and increasing genetic variance in the population. Choosing appropriate indexes for the genetic diversity of somatic mutations depends on the specific aspect of evolutionary influence being assessed.
${D}_{PO}$). The tissue architecture of trees facilitated the accumulation of somatic mutations, which have various evolutionary effects, including enhancing fitness under strong sib competition and intense host-pathogen interactions, efficiently eliminating deleterious mutations through epistasis and increasing genetic variance in the population. Choosing appropriate indexes for the genetic diversity of somatic mutations depends on the specific aspect of evolutionary influence being assessed.

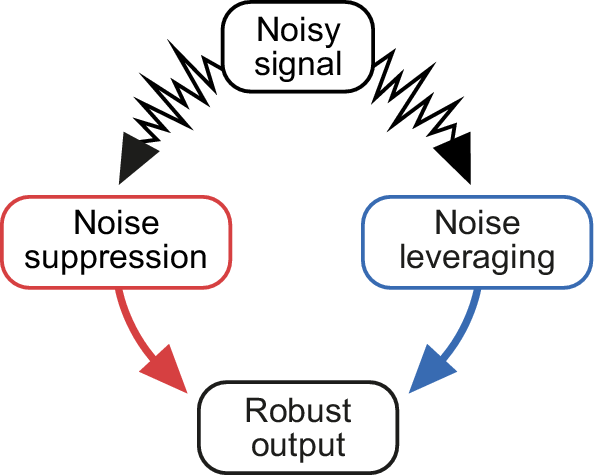
Getting it right: suppression and leveraging of noise in robust decision-making
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 27 November 2024, e10
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Quantitative analysis of the root posture of Arabidopsis thaliana mutants with wavy roots
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 25 November 2024, e9
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Contributors
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp vii-viii
-
- Chapter
- Export citation
10 - Salinity
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 194-208
-
- Chapter
- Export citation
7 - Light as a Major Driver of Algal Physiology and Evolution
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 115-135
-
- Chapter
- Export citation
Acknowledgments
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp xi-xii
-
- Chapter
- Export citation
4 - The Appearance of Eukaryotic Microalgae
- from Part I - Origins and Consequences of Early Photosynthetic Organisms
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 65-79
-
- Chapter
- Export citation
13 - Effects of Pollutants on Microalgae
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 252-271
-
- Chapter
- Export citation
9 - Nutrient Acquisition by Algae and Aquatic Embryophytes
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 151-193
-
- Chapter
- Export citation
14 - Algae in Extreme and Unusual Environments
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 272-292
-
- Chapter
- Export citation
15 - Aquatic Phototrophs and the Greenhouse Effect
- from Part III - The Future
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 295-314
-
- Chapter
- Export citation
12 - Trait Trade-Offs in Mixoplankton: An Analysis
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 227-251
-
- Chapter
- Export citation
2 - Early Photosynthetic Organisms
- from Part I - Origins and Consequences of Early Photosynthetic Organisms
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 21-42
-
- Chapter
- Export citation
Part III - The Future
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 293-386
-
- Chapter
- Export citation
Copyright page
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp iv-iv
-
- Chapter
- Export citation
8 - Temperature: Still an Enigmatic Driver in the Evolution and Physiology of Algae
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 136-150
-
- Chapter
- Export citation
17 - Variation in Nutrient Availability for Aquatic Phototrophs and Its Ecological Consequences
- from Part III - The Future
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 341-368
-
- Chapter
- Export citation
