Refine search
Actions for selected content:
11975 results in Plant sciences
Preface
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp ix-x
-
- Chapter
- Export citation
6 - The Evolution of Aquatic Embryophytes: Secondary Colonisers of Aquatic Environments
- from Part I - Origins and Consequences of Early Photosynthetic Organisms
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 96-112
-
- Chapter
- Export citation
5 - The Appearance of Macroalgae: Evolution and Ecological Consequences of Multicellularity
- from Part I - Origins and Consequences of Early Photosynthetic Organisms
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 80-95
-
- Chapter
- Export citation
Part I - Origins and Consequences of Early Photosynthetic Organisms
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 19-112
-
- Chapter
- Export citation
18 - Algae: New Products and Applications
- from Part III - The Future
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 369-386
-
- Chapter
- Export citation
16 - Ultraviolet Radiation Effects under Climate Change
- from Part III - The Future
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 315-340
-
- Chapter
- Export citation
Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 113-292
-
- Chapter
- Export citation
Index
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 387-407
-
- Chapter
- Export citation
3 - …And Nothing Was the Same Anymore: The Rise in O2 and Consequences for Photoautotrophs
- from Part I - Origins and Consequences of Early Photosynthetic Organisms
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 43-64
-
- Chapter
- Export citation
11 - Desiccation
- from Part II - Physiology of Photosynthetic Autotrophs in Present-Day Environments
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 209-226
-
- Chapter
- Export citation
Contents
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp v-vi
-
- Chapter
- Export citation
1 - Environmental Changes Impacting on, and Caused by, the Evolution of Photosynthetic Organisms
-
-
- Book:
- Evolutionary Physiology of Algae and Aquatic Plants
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024, pp 1-18
-
- Chapter
- Export citation

Evolutionary Physiology of Algae and Aquatic Plants
-
- Published online:
- 24 October 2024
- Print publication:
- 07 November 2024
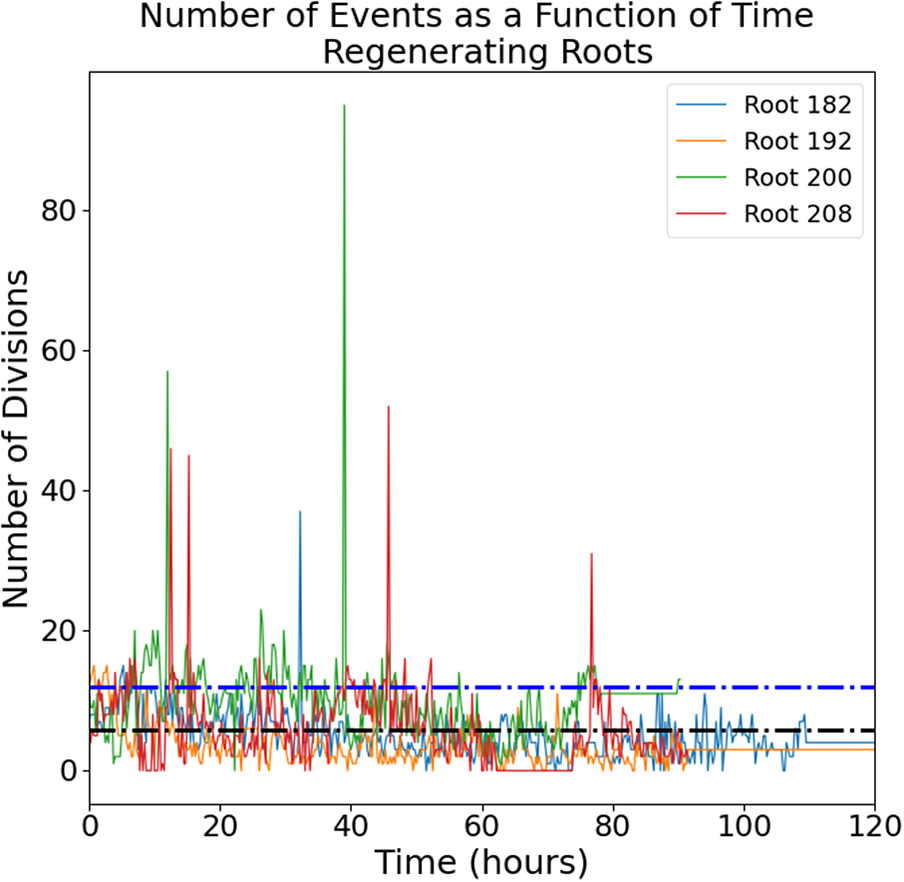
Intermittent cell division dynamics in regenerating Arabidopsis roots reveals complex long-range interactions
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 04 September 2024, e7
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
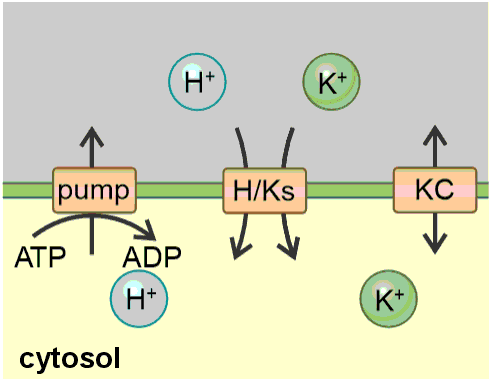
Homeostats: The hidden rulers of ion homeostasis in plants
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 03 September 2024, e8
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
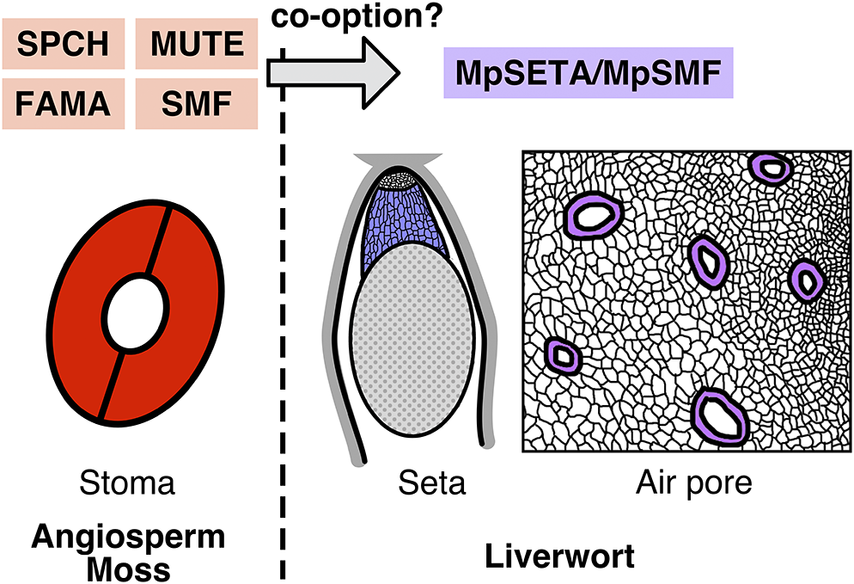
Beyond stomatal development: SMF transcription factors as versatile toolkits for land plant evolution
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 31 May 2024, e6
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
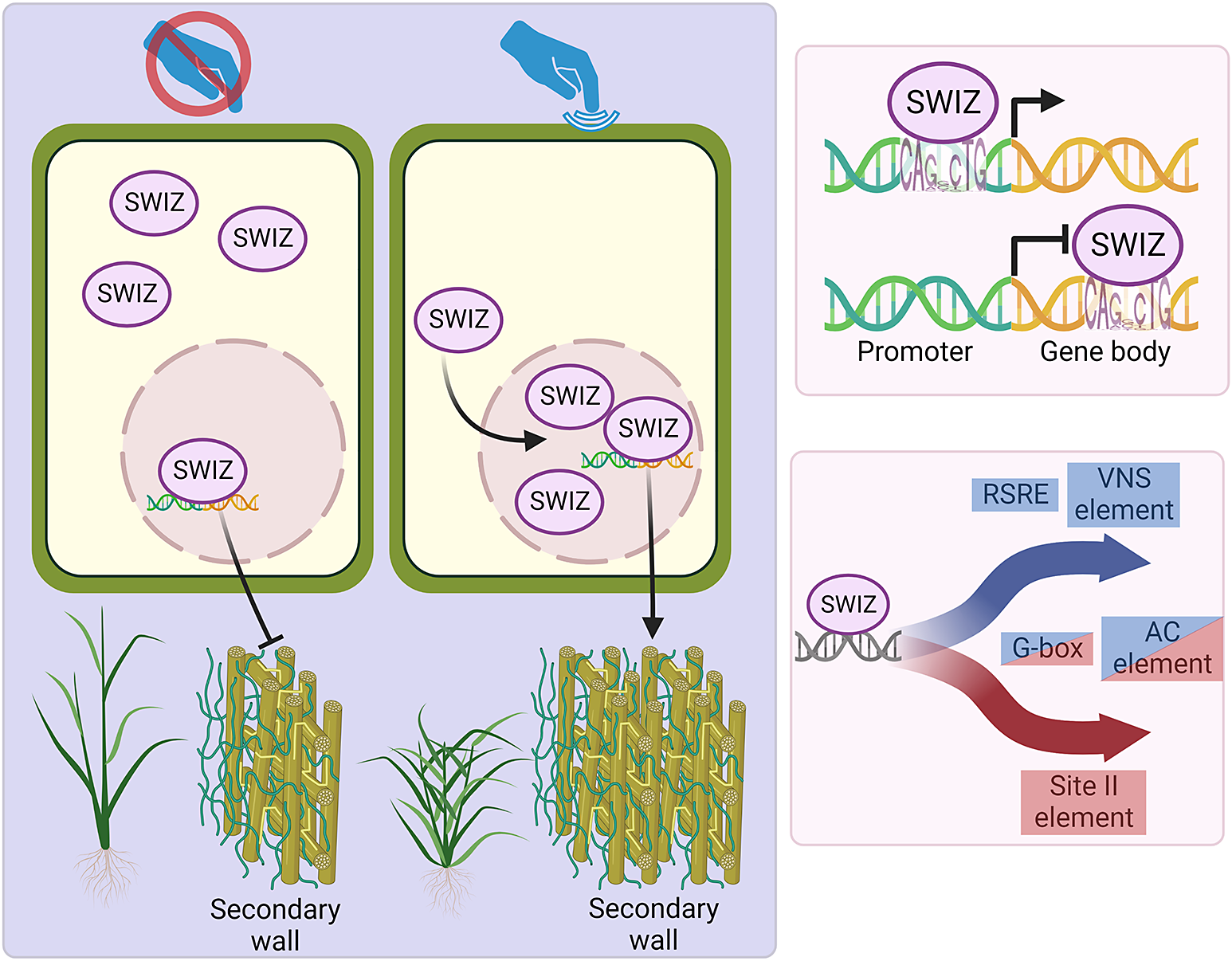
Mechanically induced localisation of SECONDARY WALL INTERACTING bZIP is associated with thigmomorphogenic and secondary cell wall gene expression
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 03 May 2024, e5
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A coordinated switch in sucrose and callose metabolism enables enhanced symplastic unloading in potato tubers
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 17 April 2024, e4
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Debunking the idea of biological optimisation: quantitative biology to the rescue
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 03 April 2024, e3
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
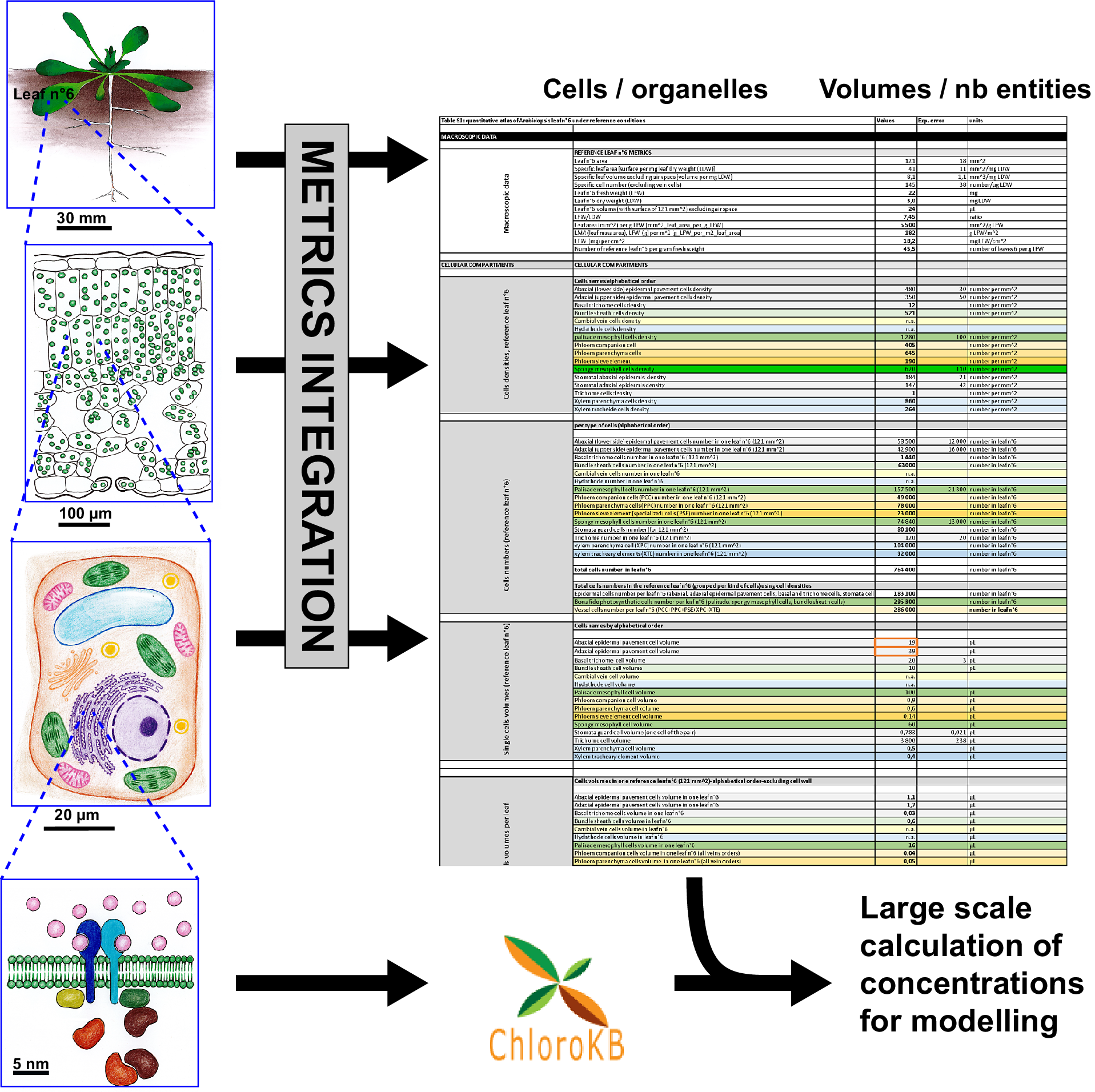
The Arabidopsis leaf quantitative atlas: a cellular and subcellular mapping through unified data integration
-
- Journal:
- Quantitative Plant Biology / Volume 5 / 2024
- Published online by Cambridge University Press:
- 29 February 2024, e2
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
