


1. Introduction
Imagine living in a town where most of the residents are your genetic relatives—grandparents, uncles, aunts, siblings, and cousins. Is this high prevalence of kin advantageous or problematic? It depends. For a newborn, it offers great opportunities, as kin are especially likely to provide the resources and protection crucial for survival and growth. In contrast, for a post-puberty young adult, being surrounded by many relatives may be less favorable, as it limits opportunities for mating and friendship outside the family circle. And for parents with young children, the availability of extended family can ease the heavy demands of childcare by offering reliable alloparenting. This example highlights how ecological affordances—the opportunities and threats an environment poses for achieving one’s goals—can differ across life stages as prioritized goals shift with development.
Behavior is shaped by the environments people inhabit. Research from a behavioral ecological perspective reveals that variations in thinking and behavior across societies reflect variations in the physical and social environments that individuals navigate (Nettle, Reference Nettle2009; Nettle et al., Reference Nettle, Gibson, Lawson and Sear2013; Sng et al., Reference Sng, Neuberg, Varnum and Kenrick2018). Yet, environmental influences do not operate in isolation. A core principle in psychology is that behavior emerges from the interaction between the environment and the individual (Lewin, Reference Lewin1951). The opportunities and threats afforded by an environment depend on the individual’s goals. These goals, in turn, drive strategies aimed at maximizing relevant opportunities while minimizing relevant threats. Life-history theory suggests that goal priorities shift systematically across development, implying corresponding changes in how individuals perceive and manage ecological affordances at different life stages. Little is known, however, about how human-environment interactions vary across the lifespan.
To address this gap, we propose a framework that integrates an affordance-management perspective with life-history theory. This conceptual framework provides a foundation for investigating human-environment interactions across development—organizing existing knowledge, generating novel and testable hypotheses, and providing a roadmap for systematic research. It aims to address three critical questions:
-
(1) How do ecological affordances change as an individual’s goal priorities shift across life stages?
-
(2) How do these emergent ecological affordances facilitate or hinder the achievement of stage-specific goals?
-
(3) What strategies do individuals employ to manage ecological affordances in pursuit of their prioritized goals across life stages?
First, we outline the central perspectives underpinning the current framework and define key concepts. Next, we apply this framework to three critical environmental dimensions tied to recurring adaptive challenges critical to survival and reproduction throughout human history: genetic relatedness, physical violence, and sex-age ratio. We examine how the perception and management of these dimensions shift across three life stages characterized by distinct goal priorities: childhood, mating, and parental stages. Finally, we position the framework in relation to existing concepts and theories, highlighting its connections to and departures from traditional perspectives while emphasizing its novel contributions and implications for understanding human-environment interactions across development.
2. A proposed framework of ecological affordances across life stages
2.1. Goals shape ecological affordances
A person’s ability to achieve goals depends on the opportunities and threats afforded by the objects, events, and other people around them. Importantly, these afforded opportunities and threats are not inherent to the properties of those objects, events, and people. Rather, they emerge from the interaction between (a) the objective characteristics and features of these elements and (b) the individual’s needs, desires, and vulnerabilities and the goals generated by them. For example, when a person is tired and motivated to rest, a chair is perceived as affording sitting. However, if a stranger suddenly intrudes into this person’s home and the person is motivated to protect themselves, the same chair may be perceived as affording a barrier for self-defense. Nothing about the physical properties of the chair has changed, yet because the goal of the actor has changed, the perceived affordances of the chair have changed as well. An affordance-management approach emphasizes the role of one’s goals in shaping which potential affordances are perceived and acted upon (Gibson, Reference Gibson1979; McArthur & Baron, Reference McArthur and Baron1983; Neuberg et al., Reference Neuberg, Kenrick and Schaller2010). Although objects, events, and people possess features that potentially imply many opportunities and threats, the affordances perceived and acted upon depend on the perceiver’s goals.
This approach extends to ecology, which examines the interrelationship between (a) environments and (b) the organisms that inhabit them (Begon et al., Reference Begon, Townsend and Harper2012; Davies, Reference Davies2012). Environments contain objective features—measurable aspects that exist independently of perception. Although these features present various potential affordances, they become psychologically meaningful when individuals perceive them in relation to their goals. Ecological affordances, therefore, emerge from the interaction between (a) the objective features of an environment and (b) an individual’s goals. For instance, an environment with a high prevalence of kin offers opportunities for those prioritizing securing resources but poses threats for those seeking mates. Thus, even when the environment includes the same objective features, the psychological salience and functional relevance of specific ecological affordances vary according to an individual’s prioritized goals.
2.2. Goal priorities shift across life stages
Across the lifespan, humans face the challenge of limited time, resources, and energy. Natural selection favors those who efficiently acquire and allocate these resources to maximize reproductive fitness (Del Giudice et al., Reference Del Giudice, Gangestad, Kaplan and Buss2015; Stearns, Reference Stearns1992; Stearns & Rodrigues, Reference Stearns and Rodrigues2020). From a life-history theory perspective, development involves trade-offs in when and how to allocate these scarce resources. As individuals progress through developmental hierarchy, somatic efforts—such as bodily growth and skill acquisition—set the foundation for mating efforts, which, in turn, support parenting efforts (Alexander, Reference Alexander1987; Kenrick et al., Reference Kenrick, Griskevicius, Neuberg and Schaller2010). Importantly, earlier life-history goals are not replaced by later-stage goals but continue to operate alongside them.
These trade-offs drive shifts in prioritization among various life stage goals. During the childhood life stage—from birth to the end of juvenile growth and the onset of sexual maturation—development focuses on somatic maintenance and growth. Children prioritize maintaining health, growing body size, and acquiring the necessary resources and skills that will support later life stages. During the mating life stage—from the onset of sexual maturation to first childbearing—the focus shifts toward mate attraction and reproduction, while earlier goals, such as maintaining health and continuing growth, remain important. Young adults prioritize immediate mate attraction and reproduction while investing in embodied capital (e.g., body size, strength, skill, and knowledge) that will contribute to long-term reproductive success. During the parenting life stage—following childbearing—the focus shifts toward both optimizing quantity and quality of offspring as well as maintaining mate retention. Parents prioritize providing care, protection, and resources for their children while also maintaining stable relationships with mates to support cooperative parenting that ensures their children’s survival and reproduction. Simultaneously, earlier goals, such as personal health and embodied capital, remain critical for successful parenting, as they directly affect the ability to care for children. Life-history theory suggests that each stage involves recalibrating prioritization among various goals to meet the demands of the current life stage, while building on the foundation of previous stages and laying the groundwork for future ones.Footnote 1
Notably, these trade-offs also differ between males and females, particularly after sexual maturation. These sex differences stem from biological reproductive asymmetries and have significant implications for mating and parenting strategies. Females bear higher costs in gestation and lactation but have greater genetic certainty regarding their offspring. Consequently, females tend to be more selective in mate choice and to invest more heavily in parental care compared to males (Clutton-Brock & Parker, Reference Clutton-Brock and Parker1992; Trivers, Reference Trivers1972). This selectivity often leads to intense competition among males for access to mates, resulting in greater male investment in embodied capital prior to mating and riskier strategies that may increase mating opportunities despite elevated mortality risks.
Males, having lower minimum obligatory investment in parenting, are more likely to gain reproductive benefits from short-term, uncommitted sexual relationships—a pattern observed across cultures (Schmitt, Reference Schmitt2005). However, human males are distinctive among mammals, with unusually high parental investment seen in fewer than 5% of mammalian species. This pattern exists because biparental care is crucial for human children’s prolonged period of vulnerability and dependence (Kaplan et al., Reference Kaplan, Hill, Lancaster and Hurtado2000). Consequently, human males are highly motivated to contribute care and resources to their mates and children, and they frequently do so (Geary, Reference Geary2000, Reference Geary2005; Geary & Flinn, Reference Geary and Flinn2001; Ko et al., Reference Ko, Pick, Kwon, Barlev, Krems, Varnum, Neel, Peysha, Boonyasiriwat, Brandstätter, Crispim, Cruz, David, David, de Felipe, Fetvadjiev, Fischer, Galdi, Galindo and Kenrick2020). These between-sex dynamics, combined with within-sex variation in mating and parenting strategies (Kokko & Jennions, Reference Kokko and Jennions2008), also significantly shape goal prioritization throughout the lifespan.
2.3. Ecological affordances shift across life stages
If actors’ goals shape their perceptions of ecological affordances, and if these goals shift across life stages, then perceptions of ecological affordances will also shift across life stages (Figure 1). An environment may be perceived as facilitating the achievement of a particular goal at one life stage, whereas at another stage with different goal priorities, the same environment may be perceived as an impediment or become irrelevant.
An individual’s life stage interacts with sex to shape prioritized life stage goals, which, in turn, shape the ecological affordances—the opportunities and threats the specific features of an environment afford to that individual’s goal achievement.

2.4. Management strategies for shifting ecological affordances across life stages
To manage ecological affordances across life stages, humans may rely both on specialized adaptations for recurrent challenges and flexible recalibration of these mechanisms. First, certain environmental features likely exerted consistent selection pressures, favoring the evolution of specialized psychological adaptations to directly address challenges that posed recurrent adaptive threats or opportunities over evolutionary time. These mechanisms may facilitate the rapid and efficient processing of specific ecological affordances. For instance, detecting the relative ratio of potential mating partners to competitors in a given environment is critical for reproductive success. Indeed, humans appear to possess psychological mechanisms for assessing such features. However, the perceptual sensitivity and attentional allocation of these mechanisms are likely to be shaped by life-stage-specific ecological affordances.
Second, some ecological affordances may be addressed through the functional recalibration or repurposing of pre-existing adaptations. For instance, kin detection mechanisms likely evolved to identify genetic relatives to secure support and reduce the risks of inbreeding. Although these mechanisms are critical regardless of whether there are many or few kin around, they can be recalibrated to address the ecological affordances implied by different prevalence of kin. In environments with a low prevalence of kin, kin detection mechanisms may become more attuned to securing familial support, especially during childhood, when such support is vital for survival and development. In contrast, in environments with a high prevalence of kin, these mechanisms may instead emphasize avoiding inbreeding, particularly during the mating stage, when reproductive decisions are vital to long-term fitness outcomes.
Specifically, we propose that individuals manage shifting ecological affordances across life stages through three strategies:
-
(1) Recalibrating their cognitive and affective attunements by optimizing attention, sensitivity, and memory for ecological affordances relevant to their prioritized life stage goals
-
(2) Recalibrating their psychological and behavioral strategies in response to emergent ecological affordances relevant to their prioritized life stage goals
-
(3) Restructuring their environment to facilitate goal pursuit across various levels by:
-
(a) altering their immediate surroundings
-
(b) relocating within their broader environment
-
(c) migrating to a new environment
-
Examining ecological affordances over the entire lifespan underscores the potential for lifelong flexibility of psychology, as individuals continuously align their perceptions and management strategies with changing life stage goals.
3. Applying the framework to three illustrative environmental dimensions
We apply this framework to three environmental dimensions that contain recurrent adaptive problems critical to survival and reproduction throughout human history: genetic relatedness, physical violence, and sex-age ratio (Table 1). These dimensions create adaptive challenges that have played a crucial role in shaping life-history trade-offs, including resource allocation, cooperation-competition dynamics, reproductive strategies, and parental investment. These dimensions were selected to demonstrate the theoretical value of the framework in illuminating stage-specific variations in affordance perception and management.
Three illustrative environmental dimensions applying the current framework

For each dimension, we first review the potential affordances it may present. We then outline predictions on how the ecological affordances perceived by an individual differ at three life stages—childhood, mating, and parenting—and how these differences vary by sex in some life stages. We explore how individuals at each of these stages manage these ecological affordances.
3.1. Genetic relatedness
Kinship plays a central role in evolutionary explanations of social behavior through the concept of inclusive fitness. Investment in genetic relatives—both offspring and extended kin—would have enhanced reproductive success by promoting the transmission of shared genes to future generations (Hamilton, Reference Hamilton1964; Lee, Reference Lee2003; West & Gardner, Reference West and Gardner2013). Indeed, people across cultures share a strong motivation to foster and maintain family bonds (Ko et al., Reference Ko, Pick, Kwon, Barlev, Krems, Varnum, Neel, Peysha, Boonyasiriwat, Brandstätter, Crispim, Cruz, David, David, de Felipe, Fetvadjiev, Fischer, Galdi, Galindo and Kenrick2020; Pick et al., Reference Pick, Ko, Wormley, Wiezel, Kenrick, Al-Shawaf, Barry, Bereby-Meyer, Boonyasiriwat, Brandstätter, Crispim, Cruz, David, David, Defelipe, Elmas, Espinosa, Fernandez, Fetvadjiev and Varnum2022). Consequently, kinship has long served as a fundamental principle for organizing social life, shaping diverse cultural and psychological patterns (J. F. Schulz et al., Reference Schulz, Bahrami-Rad, Beauchamp and Henrich2019).
Ecological genetic relatedness—the prevalence of one’s kin in one’s environment (Sng et al., Reference Sng, Choi and Ackerman2024)—potentially poses various opportunities and threats.Footnote 2 First, an environment with high genetic relatedness offers opportunities for receiving altruistic support, as providing material and social support to kin can enhance the giver’s indirect reproductive success (Burkart et al., Reference Burkart, Hrdy and Schaik2009; Griffin & West, Reference Griffin and West2003). Evidence consistently shows that altruism is preferentially extended to relatives over non-relatives in both hypothetical and actual life-threatening scenarios (Burnstein et al., Reference Burnstein, Crandall and Kitayama1994; Grayson, Reference Grayson1993; McCullough & Barton, Reference McCullough and Barton1991) and to close relatives over distant ones in both traditional and modern societies (Hooper et al., Reference Hooper, Gurven, Winking and Kaplan2015; Neyer & Lang, Reference Neyer and Lang2003).
Alloparenting—cooperative childrearing by individuals other than parents—exemplifies such altruistic behaviors. Throughout human history, children have primarily received care from relatives (Kramer, Reference Kramer2005; Laughlin, Reference Laughlin2013), especially grandparents and older siblings, who facilitate intergenerational resource transfer (Hooper et al., Reference Hooper, Gurven, Winking and Kaplan2015; Kaplan & Robson, Reference Kaplan and Robson2002). These investment patterns follow genetic certainty: maternal grandmothers invest the most among grandparents, followed by maternal grandfathers and paternal grandmothers, and finally, paternal grandfathers (Kahana & Kahana, Reference Kahana and Kahana1970; Laham et al., Reference Laham, Gonsalkorale and Von Hippel2005). Thus, an environment with high genetic relatedness, especially with maternal kin, offers opportunities for alloparental investment.
In contrast, an environment of high genetic relatedness also poses the threat of inbreeding depression. Reproduction between genetic relatives can compromise offspring fitness because offspring are more likely to inherit two copies of harmful recessive genes from their parents, leading to the increased expression of genetic disorders (Bittles & Neel, Reference Bittles and Neel1994; Charlesworth & Charlesworth, Reference Charlesworth and Charlesworth1999). This genetic homogeneity also elevates susceptibility to infectious diseases (Tooby, Reference Tooby1982) and reduces offspring’s ability to adapt to environmental changes (Keller & Waller, Reference Keller and Waller2002).
In sum, an environment of high genetic relatedness affords both opportunities and threats. It can foster prosocial behavior, intergenerational resource transfer, and alloparenting while also posing a threat of inbreeding depression due to limited genetic diversity. The ecological affordances of genetic relatedness perceived by an individual, however, should depend on life stage. A summary of predictions about ecological affordances of genetic relatedness and their management strategies across life stages is presented in Table 2.
Genetic relatedness: Proposed hypotheses about ecological affordances and management strategies across life stages

3.1.1. Childhood stage
Humans, compared to other primates and mammals, exhibit a uniquely extended period of vulnerability and dependency during childhood, along with delayed maturation and independence from their parents (Kaplan et al., Reference Kaplan, Hill, Lancaster and Hurtado2000). This extended vulnerable life stage necessitates reliance on adults for both material resources of food and shelter and social learning essential for development (Nelson, Reference Nelson2020). Because kin represents the primary source of these critical resources, alloparental care substantially enhances children’s survival, maintenance, and growth (Jaeggi & Gurven, Reference Jaeggi and Gurven2013; Kramer, Reference Kramer2010). Consequently, environments with high genetic relatedness are likely perceived as offering opportunities for reliable alloparental investment (Crittenden & Marlowe, Reference Crittenden and Marlowe2008; Kenkel et al., Reference Kenkel, Perkeybile and Carter2017; Martin et al., Reference Martin, Ringen, Duda and Jaeggi2020), whereas environments lacking kin presence are likely perceived as constraining access to consistent resources and protection.
How do children navigate the ecological affordances of genetic relatedness? We hypothesize that children—mostly unconsciously—leverage evolved mechanisms to optimize care and support in the environment they interact with by (a) detecting kin and showing preferences toward them, (b) calibrating sensitivity and behaviors for monitoring and engaging caregivers, and (c) adjusting attachment strategies.
First, children possess evolved mechanisms for kinship detection that emerge early in development. Newborns demonstrate the ability to recognize their mother’s face within hours after birth (Bushnell, Reference Bushnell2001; Walton & Bower, Reference Walton and Bower1993), and children show sensitivity to reliable kinship cues—including patterns of parental caregiving and shared developmental history (Lieberman et al., Reference Lieberman, Tooby and Cosmides2007; Sznycer et al., Reference Sznycer, De Smet, Billingsley and Lieberman2016). These abilities facilitate identification of potential alloparents, leading children to prefer individuals who resemble their parents or those who have received substantial parental investment, such as their siblings and cousins. Such abilities are likely more important in an environment with low genetic relatedness, where accurately identifying reliable caregivers is essential for meeting developmental goals.
Second, children actively monitor caregiver support and act if care becomes insufficient. Even infants seek attention by establishing eye contact, tracking gazes (Hrdy, Reference Hrdy2009, Reference Hrdy2007), vocalizing (e.g., babbling; Hrdy, Reference Hrdy2007), and displaying emotions (e.g., expressing fear; Grossmann, Reference Grossmann2022). The intensity of this monitoring and engagement is likely calibrated to kin prevalence in the environment. In an environment with high genetic relatedness, where reliable alloparents are abundant, children may not need to constantly seek attention; in an environment with low genetic relatedness, children may intensify their monitoring and engagement to secure adequate care; and in an environment with extremely low genetic relatedness, such as orphanages, children may conserve energy by reducing their efforts to solicit attention, given the low likelihood of success.
Third, children form attachment bonds with multiple individuals while maintaining a primary bond, typically with the mother (Cassidy, Reference Cassidy, Cassidy and Shaver1999). The depth and breadth of these attachments likely calibrate to the kin availability in their environment. Because kin attachments (even if shallow) can buffer against negative life events, such as losing a primary attachment figure (Perry, Reference Perry2021), children in an environment with high genetic relatedness may form multiple attachments with several relatives without heavily relying on any particular kin relationship or non-kin relationships. Conversely, children in an environment with low genetic relatedness may focus on forming stronger and deeper attachments with the limited relatives available or even with non-kin (e.g., godfather, parent’s friends) to secure adequate care and overall investment.
3.1.2. Mating stage
Although perceived as advantageous by children, an environment of high genetic relatedness will likely be perceived as affording both threats and opportunities for individuals in the mating stage. Because young adults face developmental trade-offs between immediate reproduction and long-term reproductive success, their perceived ecological affordances likely depend on their specific reproductive strategies. On the one hand, an environment of high genetic relatedness makes finding genetically diverse mates more challenging, thereby increasing the risk of inbreeding depression. On the other hand, a strong kin network within an environment of high genetic relatedness can provide resources and skills essential for future mating and parenting.
The management strategies to mitigate the threats and exploit the opportunities are likely shaped by individuals’ prioritized mating goals and reproductive strategies. For example, those prioritizing immediate or short-term reproduction may address the challenges of high genetic relatedness by (a) heightening their sensitivity to detect and avoid inbreeding risks, (b) altering their surroundings within the given environment to increase contact with non-kin, and (c) relocating to an environment with a lower kin prevalence to expand mating opportunities.
First, although incest aversion shows a universal pattern—being more disgusted and avoidant for close versus distant relatives due to reproductive fitness costs (Antfolk et al., Reference Antfolk, Lieberman, Harju, Albrecht, Mokros and Santtila2018; Kresanov et al., Reference Kresanov, Kotler, Seto, Lieberman, Santtila and Antfolk2018)—its intensity appears calibrated to environmental threat. Living in environments of high genetic relatedness poses greater risks of inbreeding, requiring heightened sensitivity to detect and avoid such risks. Supporting this, young adults surrounded by many genetic relatives exhibit stronger moral condemnation of incest compared to those in low-relatedness environments (Sng et al., Reference Sng, Choi and Ackerman2024). This calibration of incest aversion is especially relevant during the mating life stage when it has significant implications for reproductive success.
Second, because interactions with non-kin became important for young adults seeking mates, they likely alter their surroundings to reduce kin interactions. Young adults begin spending less time with their parents and more time with friends and peers (Larson & Verma, Reference Larson and Verma1999; Roberts & Dunbar, Reference Roberts and Dunbar2011). Moreover, during the high fertility phase of their ovulatory cycle, women selectively avoid interactions with their fathers (Lieberman et al., Reference Lieberman, Pillsworth and Haselton2011). These behaviors are likely more pronounced in environments with high genetic relatedness.
Third, young adults are likely to relocate to an environment with lower genetic relatedness to expand their mating opportunities. In small hunter-gatherer societies, where susceptibility to low genetic diversity can be high, members of the more prevalent sex often disperse from their natal group to find mates (Kramer et al., Reference Kramer, Schacht and Bell2017). Similarly, in the U.S., young adults are more likely than other age groups to move to large urban areas (Johnson et al., Reference Johnson, Winkler and Rogers2013), where genetic diversity is greater, possibly in search of better mating opportunities, even if it means losing support from kin.
Conversely, those prioritizing future or long-term reproductive potential are likely to maximize their opportunities by delaying both courtship and dispersal from proximity to their local kinship networks. In small-scale hunter-gatherer societies, young adults often delay independence until gaining the necessary skills, physical strength, and social alliances to thrive independently (Kramer, Reference Kramer2011; Natterson-Horowitz & Bowers, Reference Natterson-Horowitz and Bowers2020). Similarly, in modern industrialized societies, many individuals undergo a phase known as ‘emerging adulthood’ (Arnett, Reference Arnett2000, Reference Arnett2007), during which they explore education, relationships, and careers while remaining dependent on their parents. These delaying strategies are likely to be more effective in an environment with high genetic relatedness, particularly where mating competition is intense.
3.1.3. Parenting stage
For parents, especially those with young children, access to alloparents is critical for both parental and offspring success. Maternal commitment—the strongest predictor of infant survival—largely depends on allomaternal support from the community (Hrdy, Reference Hrdy2007, Reference Hrdy2009). Such support also enables mothers to shorten birth intervals (Hrdy, Reference Hrdy2007; Kaplan et al., Reference Kaplan, Hill, Lancaster and Hurtado2000; Richerson & Boyd, Reference Richerson and Boyd2020). Because genetic relatives often serve as alloparents, an environment with high genetic relatedness is likely perceived to afford opportunities for parents to enhance the quality and quantity of their offspring.
How do parents with young children manage the opportunities afforded by environments of high genetic relatedness and the challenges afforded by those of low genetic relatedness? We hypothesize that parents (a) alter their immediate surroundings to facilitate alloparenting by kin or, if necessary, non-kin, depending on the prevalence of kin within their environment, and (b) relocate their family to access kin support.
To maximize the alloparenting opportunities, parents likely alter their immediate social environment by pooling their “energy budget” with available family members (Kramer & Ellison, Reference Kramer and Ellison2010; Reiches et al., Reference Reiches, Ellison, Lipson, Sharrock, Gardiner and Duncan2009). Even after forming new family units through marriage, individuals typically maintain lifelong connections with their birth families (Rodseth et al., Reference Rodseth, Wrangham, Harrigan, Smuts, Dare, Fox, King, Lee, Foley, Muller, Otterbein, Strier, Turke and Wolpoff1991) to preserve access to this shared energy budget. When biological kin are scarce in an environment, people form “fictive kin” networks—family-like relationships with individuals unrelated by blood or marriage. For instance, immigrants who live at great distance from their biological families often create social capital with others from similar backgrounds to simulate and recreate the benefits typically afforded by environments of high genetic relatedness (Ebaugh & Curry, Reference Ebaugh and Curry2000; Regnerus & Burdette, Reference Regnerus and Burdette2006; Taylor et al., Reference Taylor, Chatters, Woodward and Brown2013). Religious groups similarly foster a subjective sense of fictive kin by promoting “brotherhood” and “sisterhood” among members and establishing ritual kin, such as godparenthood (E. B. Hubbard et al., Reference Hubbard, Shannon and Pisor2023), thereby enhancing offspring fitness through shared resources and group support (Moon et al., Reference Moon, Krems, Cohen and Kenrick2019; Weeden & Kurzban, Reference Weeden and Kurzban2013).
As a strategy to maximize the alloparenting opportunities, parents likely relocate to be near genetic relatives, particularly their own parents—children’s grandparents. For instance, foragers exhibit flexible and facultative residence patterns across their life course based on environmental resources, social needs, and kin availability (Marlowe, Reference Marlowe2010). Similar patterns persist in contemporary societies, where parents commonly relocate closer to their own parents for childcare support, or grandparents migrate to assist in childrearing (Hank, Reference Hank2007). This strategy is especially beneficial for resource-constrained families; for example, individuals with lower education and income tend to live closer to their parents than those with higher socioeconomic status (Costa & Kahn, Reference Costa and Kahn1990). The ubiquity of three-generational households reflects, in part, how people alter local genetic relatedness to maximize alloparenting benefits (Hooper et al., Reference Hooper, Gurven, Winking and Kaplan2015).
3.1.4. Summary
Individuals likely perceive ecological affordances of genetic relatedness differently across life stages. In childhood, they may perceive high genetic relatedness as affording opportunities to receive support for survival and development. During the mating stage, they may perceive it as affording both threat and opportunity—limiting immediate reproduction while enhancing future reproductive potential. In parenting, they may perceive it as affording opportunities for alloparenting to improve offspring success. Individuals thus likely calibrate their strategies to optimize these stage-specific affordances of genetic relatedness.
3.2. Physical violence
Historically, physical violence has been a significant cause of human mortality and morbidity (Chagnon, Reference Chagnon1988; Schaller & Neuberg, Reference Schaller and Neuberg2008; Tooby & Cosmides, Reference Tooby and Cosmides1988) and continues today through wars and crime. To mitigate the threat of physical violence, humans have evolved precautionary self-protection systems designed to detect cues that signal intentional harm, triggering a set of adaptive responses to either escape from or removal of the threat (Neuberg et al., Reference Neuberg, Kenrick and Schaller2011). For instance, humans exhibit heightened and more persistent fear responses to evolutionarily recurrent threats, such as male outgroup faces, than to novel or contemporary dangers (e.g., Mallan et al., Reference Mallan, Lipp and Cochrane2013; Navarrete et al., Reference Navarrete, Olsson, Ho, Mendes, Thomsen and Sidanius2009).
Physical violence is a universal threat, but its forms, frequency, context, motivations, and consequences vary by sex and life stage, aligning with different reproductive strategies (Ellis et al., Reference Ellis, Del Giudice, Dishion, Figueredo, Gray, Griskevicius, Hawley, Jacobs, James, Volk and Wilson2012). During childhood, both boys and girls face relatively similar homicide risks, but this changes dramatically after puberty. During adolescence and early adulthood, when competition for resources and mates is most intense, men experience much higher rates of violent victimization and perpetration, particularly in intergroup and intrasexual conflicts (United Nations Office on Drugs and Crime, 2019c). Although dangerous, these high-risk aggressive behaviors have historically offered potential reproductive advantages through resource acquisition and mate competition (Chagnon, Reference Chagnon1988; Palmer & Tilley, Reference Palmer and Tilley1995; Pemberton et al., Reference Pemberton, Insko and Schopler1996). In contrast, women face elevated risks as targets of sexual and intimate partner violence, particularly during their peak reproductive years, driven by male competition for reproductive access and control (Sardinha et al., Reference Sardinha, Yüksel-Kaptanoğlu, Maheu-Giroux and García-Moreno2024; World Health Organization, 2021).
The same environment characterized by varying prevalence of different subtypes of physical violence creates distinct risks for males and females, based on the specific types of aggression each is more vulnerable to. An environment with a high prevalence of intergroup and intrasexual violence is likely perceived as affording greater threat by young men, whereas an environment with a high prevalence of intersexual violence and sexual victimization is likely perceived as affording greater threat by young women. These sex-specific risks vary further across different life stages, leading males and females to perceive distinct ecological affordances over the lifespan, each requiring unique management strategies.
We hypothesize that: (a) males and females display different levels of fear and anger sensitivity at various life stages, with fear driving avoidance and anger prompting confrontation; (b) cooperation strategies shift across life stages to balance the costs and benefits of engaging in violent behavior; (c) these flexibilities influence broader social attitudes, including stigmatization, prejudice, and discrimination toward specific groups; and (d) these factors affect decisions about whether to stay in or relocate to a new environment. A summary of predictions about ecological affordances of physical violence across life stages and the corresponding management strategies is provided in Table 3.
Physical violence: Proposed hypotheses about ecological affordances and their management strategies across life stages

3.2.1. Childhood stage
An environment with a high prevalence of physical violence is likely perceived as affording threats to children by (a) increasing the risk of family violence, (b) limiting caregiving when parents are harmed, and (c) exposing children to community violence. First, child victimization is often linked to several risk factors that are more prevalent in environments with high physical violence, such as social and economic instability (Daly & Wilson, Reference Daly and Wilson1988; Frankenhuis & Amir, Reference Frankenhuis and Amir2022; Pinheiro, Reference Pinheiro2006), parental mental health issues (Kohl et al., Reference Kohl, Jonson-Reid and Drake2011), and domestic violence (Jouriles et al., Reference Jouriles, McDonald, Smith Slep, Heyman and Garrido2008). These factors elevate caregiving stress, leading parents to become perpetrators or victims, ultimately compromising their caregiving abilities (Margolin & Gordis, Reference Margolin and Gordis2004). Second, parental victimization also deprives children of consistent care and access to essential resources, heightening children’s vulnerability and hindering their development (Belsky, Reference Belsky1984; Shonkoff et al., Reference Shonkoff, Garner, Siegel, Dobbins, Earls, Garner, McGuinn, Pascoe and Wood2012). Third, children in such environments often become targets of community violence—whether as direct victims, collateral damage, or through forced recruitment into gangs—perpetuating cycles of harm (Howell & Griffiths, Reference Howell and Griffiths2018).
How do children mitigate the threats of living in an environment with high physical violence? We hypothesize that children (a) calibrate their attention and sensitivity to identifying potential perpetrators and (b) adjust their behavioral strategies according to their own physical formidability.
Children in violent environments are likely to exhibit heightened attention and sensitivity to cues of potential harm. Historically, children have faced significant mortality risks from outgroup members (Hill & Hurtado, Reference Hill and Hurtado2018). As an evolved protective mechanism, infants exhibit greater fear toward male strangers once they begin to crawl. This stranger anxiety is observed across cultures regardless of the primary caregiver’s gender, challenging the idea that this fear stems solely from familiarity with females (Hahn-Holbrook et al., Reference Hahn-Holbrook, Holbrook, Bering, Holbrook and Bering2010). Additionally, children exposed to physical abuse develop greater accuracy, response bias, and sensitivity in detecting anger—an emotion strongly associated with imminent physical harm. Notably, these children do not show the same level of attunement to other negative emotions, such as disgust, fear, or sadness, indicating a selective focus on cues most relevant to their safety (Pollak et al., Reference Pollak, Cicchetti, Hornung and Reed2000, Reference Pollak, Klorman, Thatcher and Cicchetti2001). Children in violent environments are likely to develop increased fear of outgroup males and enhanced sensitivity to cues relevant to harm, thereby enhancing their ability to identify potential perpetrators and mitigate risks.
In addition, children in violent environments are likely to calibrate their behavioral strategies based on their own physical formidability. For physically larger and stronger children, aggression functions as an adaptive strategy to secure resources and deter threats (Dodge & Frame, Reference Dodge and Frame1982; Ellis et al., Reference Ellis, Bianchi, Griskevicius and Frankenhuis2017; Hubbard et al., Reference Hubbard, Dodge, Cillessen, Coie and Schwartz2001; Light & Dishion, Reference Light and Dishion2007). This strategy manifests through (a) bullying—targeted aggression toward more vulnerable peers (Kolbert & Crothers, Reference Kolbert and Crothers2003; Olthof & Goossens, Reference Olthof and Goossens2007; Veenstra et al., Reference Veenstra, Lindenberg, Munniksma and Dijkstra2010; Volk et al., Reference Volk, Camilleri, Dane and Marini2012a, Reference Volk, Camilleri, Dane and Marini2012b), (b) engaging in antisocial behaviors that signal strength and deter potential rivals (Ellis et al., Reference Ellis, Del Giudice, Dishion, Figueredo, Gray, Griskevicius, Hawley, Jacobs, James, Volk and Wilson2012), and (c) affiliating with peer groups characterized by volatility or risk-taking behaviors, which enhances perceived dominance and social capital (e.g., Schwartz & Proctor, Reference Schwartz and Proctor2000). In contrast, children who perceive themselves as physically vulnerable are likely to adopt different strategies, such as (a) forming friendships with older, stronger, or socially influential peers to gain protection and bolster their security or (b) forming alliances with peers who share similar vulnerabilities, thereby fostering mutual support that enhances resilience in a threatening environment.
3.2.2. Mating stage
After puberty, young males became more effective at employing violence due to their increased strength, muscle mass, and aggressive inclinations. Because males can increase reproductive success by mating with multiple partners, young males face intense intrasexual competition for mates, leading them to pursue high-risk aggressive strategies (Tooby & Cosmides, Reference Tooby and Cosmides1988; Wang et al., Reference Wang, Kruger and Wilke2009). Successful warriors and hunters historically dominated material, social, and reproductive resources (Chagnon, Reference Chagnon1988; Wilson, Reference Wilson1989). As a result, much of male aggression is directed toward other males, manifesting as intergroup conflict, public assaults, and peer aggression, especially during the mating stage (McDonald et al., Reference McDonald, Navarrete and Van Vugt2012). Homicide rates, for instance, peak between the ages of 15 and 29, with over 80% resulting from male-on-male violence (United Nations Office on Drugs and Crime, 2019a; Hirschi & Gottfredson, Reference Hirschi and Gottfredson1983; Moffitt, Reference Moffitt1993).
Young men also often engage in physical violence toward females—particularly those in their reproductive prime—aimed at controlling female sexuality for their reproductive advantage (Smuts, Reference Smuts1992). In the U.S., 90% of rape victims are female, one in four females experience some form of physical and sexual violence by an intimate partner, and half of female murder victims are killed by intimate partners (Ertl et al., Reference Ertl, Sheats, Petrosky, Betz, Yuan and Fowler2019; Smith et al., Reference Smith, Zhang, Basile, Merrick, Wang, Kresnow and Chen2018; Snyder, Reference Snyder2000). Young females in their mating stage are more vulnerable to sexual violence than young men or older women (National Coalition against Domestic Violence, 2020; Smith et al., Reference Smith, Zhang, Basile, Merrick, Wang, Kresnow and Chen2018) as they are perceived as more desirable mates (Buss, Reference Buss1989; Conroy-Beam & Buss, Reference Conroy-Beam and Buss2018; Kenrick & Keefe, Reference Kenrick and Keefe1992).
An environment with a high prevalence of physical violence is likely perceived as affording threats to young men, elevating the risks of mortality and morbidity mainly through male-male aggression. However, for a few dominant, physically formidable, and aggressive men, this environment can be perceived as affording an opportunity to gain access to material, social, and sexual resources (Daly & Wilson, Reference Daly and Wilson1988; Wilson, Reference Wilson1989). In such environments, the immediate benefits for young men able to dominate through violence can outweigh the long-term risks of injury or death. In contrast, for young women, such environments are likely perceived as affording a threat of sexual victimization. War zones illustrate this pattern, where young women are frequently targeted for sexual violence and trafficking (United Nations Office on Drugs and Crime, 2019b; Vikman, Reference Vikman2005).
How do young men and women navigate the ecological affordances of an environment of high physical violence? Given sex differences in vulnerability and the varying costs and benefits of aggression, we hypothesize that people develop sex-specific strategies by (a) adjusting cognitive biases to identify those most capable of causing harm, (b) calibrating attention and sensitivity to detect the types of aggression they are most vulnerable to, and (c) forming strategic alliances and relationships.
First, people in violent environments—both males and females across life stages—are likely to develop heightened fear responses specifically to the anger of young men, who typically possess the greatest ability and inclination to inflict harm. Because those most capable of causing harm are especially likely to signal this via displays of anger (Sell et al., Reference Sell, Tooby and Cosmides2009), missing cues of anger carries high potential costs (Haselton & Buss, Reference Haselton and Buss2000; Nesse, Reference Nesse2005). Thus, people tend to overperceive anger in the faces of young men, even when their expressions are neutral (Maner et al., Reference Maner, Kenrick, Becker, Robertson, Hofer, Neuberg, Delton, Butner and Schaller2005; Neel et al., Reference Neel, Becker, Neuberg and Kenrick2012). Anger displayed by young men is particularly potent in triggering persistent fear conditioning, unlike anger from women (Mazurski et al., Reference Mazurski, Bond, Siddle and Lovibond1996). In an environment of high physical violence, people are likely to develop heightened vigilance and cognitive biases towards aggression cues from young men.
Second, young men in violent environments are likely to develop a heightened sensitivity to detect aggressive intentions from other men, whereas young women are likely to become more attuned to recognizing cues of sexual aggression. Indeed, male biases against outgroup males are frequently driven by concerns over aggression, status competition, and dominance, whereas female biases are often rooted in fear of sexual coercion (Navarrete et al., Reference Navarrete, McDonald, Molina and Sidanius2010). In a violent environment, young men and women—each target of violence, but in different ways—are expected to exhibit pronounced sensitivities and biases toward the threats to which they are most vulnerable.
Third, young men and women in violent environments are likely to adopt distinct social strategies tailored to their vulnerabilities. To minimize the threats and maximize the opportunities, physically formidable, aggressive, and dominant young men are likely to prioritize forming strong same-sex peer groups, such as gangs, to consolidate power, reduce vulnerability, and facilitate resource acquisition in the face of intergroup threats (Balliet et al., Reference Balliet, Li, Macfarlan and Van Vugt2011; Van Vugt et al., Reference Van Vugt, De Cremer and Janssen2007). Conversely, young men vulnerable to aggression are likely to form alliances with stronger or socially influential males for protection or seek support from other males who share similar vulnerabilities, fostering mutual resilience.
In contrast, young women in violent environments are likely to prioritize partnerships with men who can offer protection. Women in violent environments prefer partners who are physically formidable and socially dominant, as these traits provide security (Ryder et al., Reference Ryder, Maltby, Rai, Jones and Flowe2016; Snyder et al., Reference Snyder, Fessler, Tiokhin, Frederick, Woo and David2011). Higher national homicide rates are linked to greater female preferences for masculine features that signal dominance (Brooks et al., Reference Brooks, Scott, Maklakov, Kasumovic, Clark and Penton-Voak2011). However, women face a complex trade-off: Although physically formidable partners may provide external security, they may also pose internal risks of intimate partner violence. Navigating this balance between the protective benefits and potential costs of such partnerships is likely a critical challenge for women in violent environments.
3.2.3. Parenting stage
An environment of high physical violence is likely perceived as affording threats to parents by (a) endangering their own safety and survival, limiting their reproductive potential and reducing overall fitness, (b) impairing their ability to invest in existing offspring due to injury or death, indirectly lowering fitness (Hurtado & Hill, Reference Hurtado and Hill1992; Scelza, Reference Scelza2010), and (c) directly threatening the safety and survival of their offspring, further compromising parental fitness.
How do parents mitigate these threats of living in an environment with a high prevalence of physical violence? We hypothesize that parents (a) exhibit heightened vigilance toward potential threats to their children, (b) calibrate their risk-taking behaviors to balance personal safety with the need to protect their offspring, (c) adopt stricter moral judgments toward transgressors to enforce social order and reduce conflict, and (d) actively seek to modify both physical and social environments to enhance security and communal support.
First, parents are likely to calibrate their vigilance and thresholds for detecting potential threats to their children. Parental precaution systems are particularly attuned to threats to offspring (Hahn-Holbrook et al., Reference Hahn-Holbrook, Holbrook and Haselton2011), functioning similarly to evolved fear responses to predators or heights (Boyer & Bergstrom, Reference Boyer and Bergstrom2011). Throughout human evolutionary history, violence from unrelated males posed significant threats to children (Hahn-Holbrook et al., Reference Hahn-Holbrook, Holbrook, Bering, Holbrook and Bering2010; Hill & Hurtado, Reference Hill and Hurtado2018), perhaps explaining why parents still express heightened concern about harm or abduction by strangers despite the contemporary statistical rarity of such events (Hahn-Holbrook et al., Reference Hahn-Holbrook, Holbrook and Haselton2011). This evolved sensitivity manifests when parental status is activated: parents tend to overestimate both the danger posed by formidable men (Fessler et al., Reference Fessler, Holbrook, Pollack and Hahn-Holbrook2014) and risks in ambiguous situations (Eibach et al., Reference Eibach, Libby and Gilovich2003; Eibach & Mock, Reference Eibach and Mock2011) while becoming more sensitive to cues of outgroup danger (Gilead & Liberman, Reference Gilead and Liberman2014)—all as a part of a conservative error management strategy (Haselton & Buss, Reference Haselton and Buss2000; Nesse, Reference Nesse2005). Such parental precautionary patterns are likely even more pronounced for parents navigating violent environments.
Second, parents are likely to calibrate their risk-taking behaviors to balance personal safety with offspring protection. Parenthood typically reduces risk-taking, particularly when decisions affect their children’s safety and well-being (Görlitz & Tamm, Reference Görlitz and Tamm2020; Wang et al., Reference Wang, Kruger and Wilke2009). Even parents with criminal backgrounds tend to engage in less criminal and violent behavior when they have strong, satisfying relationships with their children, particularly if the child was wanted (Ganem & Agnew, Reference Ganem and Agnew2007; Giordano et al., Reference Giordano, Seffrin, Manning and Longmore2011). However, when offspring face threats, parents’ protective instincts often override their usual risk aversion, leading to increased risk-taking behaviors prioritizing child protection over personal safety (Boyer & Bergstrom, Reference Boyer and Bergstrom2011; Trivers, Reference Trivers1972). Neural evidence supports this flexibility: fathers-to-be show increased activation in motor areas of the brain when imagining threats to their own infants, as compared to unknown infants, suggesting a neural readiness to act in defense of their offspring (Rilling & Mascaro, Reference Rilling and Mascaro2017; van ‘t Veer et al., Reference van ‘t Veer, Thijssen, Witteman, Van IJzendoorn and Bakermans-Kranenburg2019). These risk-taking behaviors prioritizing child protection likely become more pronounced in violent environments where threats to offspring are more frequent and severe.
Third, parents are likely to calibrate their moral judgments to protect their offspring from social threats, with this tendency becoming particularly pronounced in violent environments. Research shows that parenting motivation predicts greater social conservatism and harsher moral evaluations (Kerry & Murray, Reference Kerry and Murray2018). Parents express stronger condemnation of adult transgressions that threaten social order while showing greater leniency toward children’s misbehavior that carries lower social costs (Hofer et al., Reference Hofer, Buckels, White, Beall and Schaller2018). Because maintaining social order and minimizing conflict is crucial for family survival and offspring well-being, parents in violent environments likely develop even more stringent moral judgments toward adult transgressors who pose potential threats to social stability.
Last, parents are likely to alter both their physical and social environments to protect their children. At the physical level, parents likely enhance security through direct modifications such as installing stronger locks or fencing. When possible, relocating to a safer, more stable environment may be likely (Sharkey & Sampson, Reference Sharkey and Sampson2010). At the social level, parents likely create protective buffers for their children by engaging in community advocacy or political action to reduce local violence. Additionally, parents may cultivate strategic networks or fictive kin relationships (e.g., godparents) to strengthen communal bonds that offer mutual support. These networks can offer social buffers, providing emotional coping, practical support (e.g., shared childcare), and a greater sense of community, which helps families navigate the challenges of living in violent neighborhoods (Ceballo & McLoyd, Reference Ceballo and McLoyd2002; Sampson et al., Reference Sampson, Raudenbush and Earls1997). By leveraging both physical and social strategies, parents can better mitigate the threats posed by violent environments and create a more secure space for their families.
3.2.4. Summary
Consideration of the ecological dimension of physical violence reveals that perceived ecological affordances can differ for males and females, even within the same life stage. In childhood, both boys and girls may perceive such an environment as affording threat via family violence, compromised caregiving, and exposure to community violence. During the mating stage, perceptions become sex-specific: whereas most young men may perceive threats from male-male aggression, physically formidable men may see opportunities for status and resource gain, and women may perceive threat from sexual victimization. In the parenting stage, adults may perceive environments of high physical violence as affording threats to offspring survival and parenting capacity. These life-stage- and sex-specific perceptions likely guide how individuals navigate violent environments.
3.3. Sex-age ratio
The sex ratio—the number of males per 100 females in a population—varies across societies and life stages. In 2014, the U.S. had a sex ratio of 105 for newborns and 75 for 80-year-olds, whereas China’s ratio was 114 for newborns and 84 for 80-year-olds (Population Division, United Nations, 2019). These patterns emerge from multiple influences. Although the sex ratio at conception is balanced, biological factors such as higher female miscarriage rates naturally result in a ratio of approximately 105 boys per 100 girls at birth (Orzack et al., Reference Orzack, Stubblefield, Akmaev, Colls, Munné, Scholl, Steinsaltz and Zuckerman2015). Sociocultural practices, such as son preference in certain Asian countries, can further skew these ratios through sex-selective abortion (Chao et al., Reference Chao, Gerland, Cook and Alkema2019; Gellatly & Petrie, Reference Gellatly and Petrie2017; Jiang et al., Reference Jiang, Yu, Yang and Sánchez-Barricarte2017). As populations age, sex ratios tend to decline due to differential mortality rates stemming from boys’ higher susceptibility to birth complications and infectious diseases (Giefing-Kröll et al., Reference Giefing-Kröll, Berger, Lepperdinger and Grubeck-Loebenstein2015; Sawyer, Reference Sawyer2012), young men’s greater exposure to violence (Cronholm, Reference Cronholm2006), and women’s longer life expectancy (Austad, Reference Austad2006; Cullen et al., Reference Cullen, Baiocchi, Eggleston, Loftus and Fuchs2016; Goldin & Lleras-Muney, Reference Goldin and Lleras-Muney2019).
What implications arise from environmental sex ratios? Understanding the ecological affordances of sex-age ratios requires recognizing how males and females at different life stages present distinct relational opportunities and threats, with these affordances varying based on the perceiver’s own sex and life stage. For example, adult females potentially afford nurturing roles for children, close friendships for other adult females, and romantic or sexual partnerships for heterosexual adult males.
Consequently, the same objective sex-age ratio can create different affordances for males and females across life stage. For instance, the sex ratio of reproductively viable males to females—often referred to as the “operational sex ratio”—potentially affords mating opportunities and threats for intersexual choice and intrasexual competition, particularly for heterosexuals pursuing romantic relationships. When the operational sex ratio is skewed, individuals of the more prevalent sex face greater competition for mates, facing pressure to modify their mating strategies—becoming less selective in mate choice, investing more effort in courtship, and adjusting long-term vs. short-term mating strategies. In contrast, individuals of the less prevalent sex gain increased leverage in mate selection, maintaining high standards for attractiveness and resources while pursuing their preferred mating strategies (Walter et al., Reference Walter, Conroy-Beam, Buss, Asao, Sorokowska, Sorokowski, Aavik, Akello, Alhabahba, Alm, Amjad, Anjum, Atama, Duyar, Ayebare, Batres, Bendixen, Bensafia, Bizumic and Za2021). Due to these facultative calibrations of mating strategies, the sex-age ratio poses different ecological affordances for relationship stability, parental investment, and reproductive success—for actors themselves and their offspring. A summary of predictions about ecological affordances of sex-age ratio and their management strategies across life stages is presented in Table 4.
Sex-age ratio: Proposed hypotheses about ecological affordances and their management strategies across life stages

3.3.1. Childhood stage
Especially in early childhood, the sex ratio of peers has limited relationship affordance implications, given that children’s primary social interactions occur with caregivers rather than peers. Although children begin forming sex-segregated playgroups around age three or four as friendship goals emerge (Maccoby, Reference Maccoby1988; Serbin et al., Reference Serbin, Moller, Gulko, Powlishta and Colburne1994), opposite-sex peers do not yet afford specific opportunities or threats.
In contrast, the sex ratio of genetically related juveniles can affect alloparenting opportunities for young children (Kramer, Reference Kramer2011, Reference Kramer2019). In traditional societies, older siblings are the most common alloparents for infants (Kramer & Veile, Reference Kramer and Veile2018), contributing through various subsistence activities (Bird & Bird, Reference Bird and Bird2002; Crittenden et al., Reference Crittenden, Conklin-Brittain, Zes, Schoeninger and Marlowe2013; Kramer, Reference Kramer2005). The affordance implications of juvenile sex ratio, however, likely depend on ecological factors, local food targets, social organization, and opportunity costs (Bock, Reference Bock2002). For example, in an environment requiring great physical strength for subsistence, male-biased juvenile sex ratios likely afford greater opportunities for younger kin.
Adult sex ratios present complex implications for child development. In environments with female-biased ratios among genetically related adults, children likely benefit from enhanced alloparenting opportunities from their older female relatives. Research consistently shows that maternal grandmothers and older sisters contribute more significantly to child survival than maternal grandfathers and older brothers (Chapman et al., Reference Chapman, Pettay, Lummaa and Lahdenperä2018; Mace & Sear, Reference Mace and Sear2005; Sear & Mace, Reference Sear and Mace2008), reflecting evolved sex differences in nurturant behavior (Geary, Reference Geary2000). Conversely, male-biased sex ratios among unrelated adults likely afford greater paternal investment for children through increased longer-term relationship commitment from fathers (e.g., Uggla & Mace, Reference Uggla and Mace2017). Cooperative biparental investment has played a critical role in enhanceing offspring fitness (Geary, Reference Geary2000, Reference Geary2005; Geary & Flinn, Reference Geary and Flinn2001) in both small-scale (Hurtado & Hill, Reference Hurtado and Hill1992; Kaplan et al., Reference Kaplan, Lancaster, Hurtado, Booth and Crouter1998; Winking et al., Reference Winking, Gurven and Kaplan2011) and modern societies (B. L. Barber & Demo, Reference Barber and Demo2006; N. Barber, Reference Barber2000b).
How do children manage the ecological affordances posed by sex ratios across different life stage groups? We hypothesize children indirectly manage them through preferences for certain caregivers or social settings, potentially influencing caregiving allocation in extended family or communal living situations. From early development, infants show greater proficiency in processing female faces (de Haan et al., Reference de Haan, Johnson, Maurer and Perrett2001; Ramsey et al., Reference Ramsey, Langlois and Marti2005; Walton & Bower, Reference Walton and Bower1993) and display increased sociability with female strangers (Lamb et al., Reference Lamb, Hwang, Frodi and Frodi1982), suggesting an evolved expectation and preference for female caregiving engagement.
Although children can readily distinguish adults by sex (Fagot & Leinbach, Reference Fagot and Leinbach1993; Leinbach & Fagot, Reference Leinbach and Fagot1993), their ability to detect relative sex ratios remains unclear, as this requires complex cognitive mechanisms they may lack. Nevertheless, ecological sex ratios likely have important affordance implications for children through their influence on parental and alloparental investment patterns.
3.3.2. Mating stage
For young adults, the sex ratios of non-reproductively viable groups—such as children or the elderly—present minimal affordances since these groups neither facilitate nor hinder young adults’ primary developmental goals at this life stage.
The operational sex ratio—the ratio of reproductively viable adults—however, creates significant affordances that shape reproductive strategies. These ratios influence both the available pool of potential partners and the intensity of mate competition. In heterosexual populations, members of the less prevalent sex benefit from enhanced mating opportunities and greater leverage in partner selection, enabling them to maintain high selectivity. Conversely, members of the more prevalent sex face intensified intrasexual competition, necessitating strategic adjustments to better align with opposite-sex preferences for successful mate acquisition (Emlen & Oring, Reference Emlen and Oring1977; Maner & Ackerman, Reference Maner and Ackerman2020; Secord, Reference Secord1983).
How do young adults maximize the opportunities and minimize the threats of living in an environment with a biased operational sex ratio? We hypothesize that they (a) increase vigilance toward the operational sex ratio and track changes in it, (b) calibrate their reproductive strategies to align with mating competition, (c) alter their immediate environments to enhance mating opportunities, and (d) relocate to new environments.
First, young adults are likely attuned to the operational sex ratio within mating-relevant subgroups, actively tracking changes in these demographic patterns. Humans can rapidly perceive and process sex ratios, even from minimal cues such as facial photographs (Brandner et al., Reference Brandner, Dillon and Brase2020) and voice patterns (Neuhoff, Reference Neuhoff2017; Neuhoff & Sikich, Reference Neuhoff and Sikich2018). This tracking is likely refined based on both preferred partner characteristics and one’s own mate value. For instance, given that people typically form long-term relationships within their own racial or ethnic group, they may mentally construct operational sex ratios specific to their group rather than to the broader population. Similarly, age-based mating preferences lead younger individuals, such as teens and adolescents, to focus on the ratio within their age cohort to predict future mating prospects (e.g., Barber, Reference Barber2000a, Reference Barber2003). Given that one’s own mate value determines bargaining power in the mating market, individuals with lower mate value may perceive sex ratio imbalances as relatively more unfavorable and adopt cautious strategies in response (e.g., Millar et al., Reference Millar, Westfall and Walsh2019; Uggla & Mace, Reference Uggla and Mace2017).
Second, young adults of the more prevalent sex—who face greater mating competition and fewer opportunities—are likely to calibrate their mating strategies toward those preferred by the less prevalent sex. When reproductive-aged males are abundant, they tend to shift toward female-preferred strategies that favor long-term, committed, monogamous relationships. This facultative calibration often results in higher marriage rates, earlier marriages, and lower out-of-wedlock or teenage birth rates (Barber, Reference Barber2001; South & Trent, Reference South and Trent1988; Stone, Reference Stone2019). In contrast, when reproductive-aged females are abundant, they tend to shift toward male-preferred strategies that favor short-term, uncommitted, casual-sex relationships, resulting in lower marriage rates and higher teen pregnancy rates.
Third, young adults of the more prevalent, unfavored sex are likely to reconstruct their environments to improve their mating prospects, both by selectively occupying specific spaces within their environments and by expanding the environment. They may spend more time in spaces that increase their chances of success (e.g., Matz & Harari, Reference Matz and Harari2020), such as spaces where others are also actively seeking partners (e.g., clubs or bars on Friday nights) or where the sex ratio is more favorable—like coffee shops if seeking women or gyms if seeking men. However, when local options remain limited, young adults may expand their search. For instance, males in male-biased sex ratios show greater interest in and higher engagement with sex tourism (Kock, Reference Kock2021) and in seeking or purchasing brides from abroad to offset local mate shortages (Kaur, Reference Kaur2004; Kawaguchi & Lee, Reference Kawaguchi and Lee2017).
Last, young adults of the more prevalent, unfavored sex may be more likely to relocate to a new environment. In hunter-gatherer societies, which often have locally constrained mating options, a biased operational sex ratio motivates dispersal outside of the group (Kramer et al., Reference Kramer, Schacht and Bell2017). Similarly, modern young adults often migrate to urban areas, ostensibly for career or educational opportunities that enhance their reproductive potential (Johnson et al., Reference Johnson, Winkler and Rogers2013). Such relocations are likely to be even more pronounced among individuals of the unfavored sex facing heightened local competition.
3.3.3. Parenting stage
An environment with a biased operational sex ratio is likely perceived as affording both opportunities and threats for parental goals depending on the parent’s sex and their offspring’s sex and life stage. A favorable operational sex ratio for parents enables (a) securing partner investment for offspring care or (b) pursuing additional mating opportunities, whereas a favorable operational sex ratio for offspring in the mating stage enhances their offspring’s reproductive success.
First, the ecological affordances of a biased operational sex ratio likely diverge between male and female parents due to the inherent differences in obligatory reproductive investment, whereby females invest more through gestation and early care. Although greater availability of alternative mates is linked to declines in marital stability in both traditional and modern societies (Jones et al., Reference Jones, Marlowe, Hawkes and O’Connell2000; Uggla & Andersson, Reference Uggla and Andersson2018), the opportunities and threats of a male- versus female-biased operational sex ratio vary for each sex. A male-biased operational sex ratio is likely to be perceived as affording opportunities for female parents to enhance their bargaining power, increasing their chances of securing committed partners who can provide resources and care for their offspring. Such environments, however, are likely perceived as threatening by male parents, especially for those with lower mate value, as their female partners would have opportunities to leave the relationship for higher mate value males who can offer better support and investment.
Conversely, a female-biased operational sex ratio is likely perceived as affording increased mating opportunities for male parents. Although human males generally invest in their offspring, such a sex ratio provides an opportunity for male parents to engage in extra-pair mating, enabling them to father additional offspring with relatively lower investment. This strategy is particularly advantageous in environments where external threats reduce the likelihood that heavy investment in a single offspring will guarantee its survival or future success (Kokko & Jennions, Reference Kokko and Jennions2008). Such environments are likely perceived as posing threats to female parents in securing partnerships and paternal investment (Uggla & Mace, Reference Uggla and Mace2017).
Second, the ecological affordances of a biased operational sex ratio may also have implications for parents with offspring in the mating stage. The reproductive success of offspring is crucial to the parent’s long-term fitness; if offspring fail to secure mates and reproduce, parental investment loses value as the flow of genes to future generations is interrupted. These affordances should thus vary based on the sex of the offspring, not the sex of the parent. For example, a male-biased sex ratio is likely to be perceived as affording threat by parents with sons but as affording opportunity for parents with daughters when their children are in the mating stage.
We hypothesize that parents navigate the ecological affordance of a biased operational sex ratio in two ways. First, they are likely to adjust their reproductive strategies by calibrating their own parenting and mating efforts while closely monitoring their partner’s. In a female-biased operational sex ratio, male parents are likely to increase their (extra-pair) mating efforts for additional reproductive opportunities. In response, female parents likely become more vigilant and exhibit heightened jealousy toward their partner’s interactions with other women to protect their relational and paternal investment. Conversely, in a male-biased operational sex ratio, female parents are likely to demand more parenting effort from the father and may pursue additional mates if unsatisfied, prompting father vigilance and jealousy to protect relational and maternal investment.
Second, parents likely calibrate their financial or material investment in offspring based on expected reproductive success in the environment. In China, for example, parents with sons in male-biased regions are more likely to increase their savings (Wei & Zhang, Reference Wei and Zhang2011). They may recognize that boosting their son’s financial resources improves his relative mate value in a competitive marriage market, thereby increasing their own reproductive success.
3.3.4. Summary
Considering the sex-age ratio reveals that perceived ecological affordances vary by the life stages and sexes of both the actor and the target. For children, the sex ratio of their same-age peers is largely perceived as irrelevant, whereas the sex ratio of the adults in their ecology is likely perceived as posing significant affordances for their development. For young adults in the mating stage, the sex ratio of reproductively viable targets is likely perceived as affording significant mating opportunities and threats. For parents, the sex ratio of reproductively viable targets is likely perceived as having significant affordance implications for both their own mate retention prospects and for their offspring’s mate-seeking prospects once they reach the mating stage. This nuanced pattern of ecological affordances of sex-age ratios necessitates equally nuanced psychological and behavioral management strategies.
4. Other relevant concepts and theoretical perspectives
We applied the proposed framework to three ecological dimensions across three developmental life stages, suggesting that environments and life stage goals interact to shape individuals’ perceptions and management strategies of ecological affordances. Here, we examine how this framework aligns with or differs from alternative theories of environments and developmental goals, highlighting the distinctive contributions and novel insights offered by the current framework.
4.1. Lewin’s field theory
Kurt Lewin’s field theory provided a foundational understanding of human behavior as a function of the person and environment (Lewin, Reference Lewin1939, Reference Lewin1951). He argued that an individual’s “field” or psychological environment is inherently subjective and dynamic, emphasizing how individuals interpret their surroundings based on their immediate goals, psychological needs, and perceived environmental constraints and opportunities.
While building on Lewin’s emphasis on subjective human-environment interactions, our approach extends his theory in several ways. First, by incorporating a developmental perspective, we add a temporal dimension. Second, the framework goes beyond describing field dynamics to propose specific strategies individuals employ to manage their ecological affordances, articulating a layer of human agency and adaptation based on the specific goals of life-history theory. This integration reveals the potential for lifelong flexibility in how individuals perceive, interpret, and manage their ecological affordances based on shifting developmental priorities.
4.2. Theories of person × environment interaction in human development
Ecological Systems Theory, later developed into the Bioecological and Process-Person-Context-Time models, posit that developmental outcomes are shaped by four key factors: (a) proximal processes—everyday interactions with the immediate environment, such as parental or peer interactions; (b) person characteristics, including (1) demand characteristics (e.g., age and gender), (2) resource characteristics (e.g., cognitive ability and genetic predispositions), and (3) force characteristics (e.g., motivation and resilience); (c) the context—the broader environmental systems (e.g., family, school, society); and (d) time—the timing and duration of experiences (Bronfenbrenner, Reference Bronfenbrenner1992; Bronfenbrenner & Ceci, Reference Bronfenbrenner and Ceci1994; Bronfenbrenner & Morris, Reference Bronfenbrenner and Morris2007). For instance, a child with high cognitive ability who grows up in a supportive family and regularly interacts with parents and teachers in a stimulating educational setting will likely develop problem-solving skills and confidence over time. In contrast, the same child in an impoverished environment with limited resources and fewer positive interactions over time may not reach their full potential.
Evolutionary-neurodevelopmental theories, such as Biological Sensitivity Context Theory (Boyce & Ellis, Reference Boyce and Ellis2005; Ellis & Boyce, Reference Ellis and Boyce2008) and Differential Susceptibility Theory (Belsky, Reference Belsky1997; Belsky et al., Reference Belsky, Bakermans-Kranenburg and van IJzendoorn2007), also highlight the interplay between individuals and environments in shaping development. These theories posit individuals with heightened sensitivity—due to their temperament, genetic predispositions, or early life experiences—are more responsive to both positive and negative environments. In adverse and stressful environments, such individuals are more vulnerable to poor outcomes, but in supportive and enriching environments, they demonstrate enhanced resilience and thrive. This creates a “for-better-and-for-worse” pattern, where sensitive individuals are more affected by their environment.
Although these theories align with the current framework in emphasizing human-environment interactions in development, our approach offers two advancements. First, integrating the affordance-management framework, the current framework diverges from traditional theories in conceptualizing environmental conditions. Whereas conventional approaches characterize environmental conditions along a linear continuum from negative (stressful, disadvantaged) to positive (supportive, advantaged) (McLaughlin et al., Reference McLaughlin, Sheridan, Humphreys, Belsky and Ellis2021), we posit that the same environment can offer varying affordances depending on the goals prioritized at each life stage. An environment perceived as affording positive support during one life stage might be seen as affording negative obstacles during another.
Second, we extend the developmental analysis beyond early life experiences and childhood, providing a comprehensive lifespan perspective. Although each of the above theories can logically be applied to adulthood, their application across the entire lifespan remains largely unexplored. The current framework raises important questions about how environmental influences change over development—whether individuals become more or less influenced by specific features of the environment as they develop, whether sensitivity to specific features of environment vary across life stages, or whether neurobiological susceptibility remains stable over time.
Rather than viewing environment as a static entity with uniform influence, we conceptualize it as a dynamic force, its impact contingent upon the individual’s current life stage and developmental goals. This approach allows for a more nuanced analysis of human-environment interactions throughout the lifespan development.
4.3. Theories of goal shifts across the lifespan
Motivational Theory of Lifespan Development (Heckhausen et al., Reference Heckhausen, Wrosch and Schulz2010; Heckhausen & Schulz, Reference Heckhausen and Schulz1995) posits that successful development involves pursuing goals that align with life-stage-specific opportunities and challenges (Schulz & Heckhausen, Reference Schulz and Heckhausen1996). In youth and early adulthood, individuals have greater opportunities to explore and shape their environment. Thus, they use “primary control” to shape the environment to fit their goals through education and skill acquisition. In middle and later adulthood, as external opportunities decrease, they use “secondary control” to adjust their goals to fit existing environmental constraints, such as letting go of unattainable goals or focusing on smaller, more achievable ones.
Socioemotional Selectivity Theory (Carstensen et al., Reference Carstensen, Isaacowitz and Charles1999; Charles & Carstensen, Reference Charles and Carstensen2010; Giasson et al., Reference Giasson, Liao and Carstensen2018) focuses on how time perception shapes social goals across the lifespan. As young individuals perceive their future as open-ended, they prioritize knowledge acquisition and novel experiences. As perceived time becomes limited with age, the focus shifts to emotionally meaningful goals, such as deepening existing relationships and maximizing emotional well-being. People become more selective in social interactions, favoring emotional satisfaction over long-term, knowledge-seeking purposes.
The current framework aligns with these theories in sharing some common principles: (a) stage-specific developmental goals, (b) shifting goal prioritization across the lifespan, (c) flexible goal pursuit strategies, and (d) the active role individuals play in navigating life stage affordances. It differs in important ways, though.
First, from a life-history perspective, the current framework integrates reproductive goals alongside growth goals, revealing stage-specific strategies that diverge between males and females. Although growth goals remain important throughout development, reproductive goals emerge and shift across the lifespan following key biological transitions such as puberty. For example, mate-seeking behaviors become salient only after sexual maturation, while parenting emerges upon having offspring. Additionally, reproductive strategies diverge between males and females even within the same life stage due to sex-specific constraints and opportunities in reproduction. For instance, females face higher obligatory parental investment (e.g., pregnancy) and limited reproductive capacity (e.g., menopause), whereas males contend with higher mate competition pressures and reproductive success variance. These sex-specific reproductive goals and strategies interact with developmental stage to shape distinct adaptive affordances and corresponding management strategies.
Second, whereas Socioemotional Selectivity Theory focuses on how chronological age shapes time perception and subsequent goal prioritization, the current framework emphasizes life stage transitions that can occur independent of age. Unlike age-based models, life stages follow both biological imperatives and social choices – parenting, for example, can begin at various ages or may never occur. This distinction captures both inevitable biological transitions (like puberty) and deliberate social choices (like deciding to have children), providing a more nuanced understanding of human development.
By integrating life-history theory, the current framework extends beyond traditional developmental models to examine how males and females navigate their environments for reproductive success across life stages. This approach enriches existing theories while generating novel predictions about sex-specific ecological navigation.
5. Future directions and implications
5.1. Identifying ecological affordances
How do individuals identify ecological affordances? Research has documented the downstream effects of environments on beliefs, emotions, behavior, and culture. However, the cognitive mechanisms underlying the processing of environmental information remain less understood. Consider the identification of sex ratios: individuals must process multiple cues including age, gender, and perceived mate value—and then integrate this information to compute average frequencies (Brandner et al., Reference Brandner, Dillon and Brase2020). This cognitive task is complicated by incomplete and imperfect information, varying spatial scales (e.g., from home to nation), context-specific variations (e.g., military base, coffee shop), and temporal dynamics (e.g., from hours to a lifetime).
Moreover, it remains unclear whether humans possess specialized cognitive mechanisms for detecting and tracking ecological affordances across life stages, allowing for continuous reevaluation of environmental features as developmental goals shift. If such mechanisms exist, an important question is whether and how the processing of spatial and temporal information for ecological affordances expands with development. For example, newborns focus on the immediate family environment, whereas adults engage with broader geographical and temporal scales, including historical and anticipated future contexts. The apparent automatic way in which these reevaluation and expansion processes come online suggests sophisticated cognitive mechanisms operating at multiple levels: attention systems that prioritize developmentally relevant features, memory systems that store environmental statistics, computational processes that integrate historical and current information, and decision-making systems that adjust behavioral strategies accordingly.
Three key questions emerge about identifying emerging ecological affordances: First, how do biological transitions prepare cognitive systems for detecting ecological affordances? For instance, pubertal mechanisms may attune attention and sensitivities to specific environmental features relevant for emerging developmental tasks. Second, what individual differences exist in the identification of emerging ecological affordances? That is, which traits, experiences, and cognitive patterns enhance sensitivity to reevaluating environmental opportunities and threats as life stage goals shift? Third, how do individuals perceive ecological trade-offs when environments simultaneously present both opportunities and threats. Understanding how people differentially detect such trade-offs and employ strategies to navigate them would shed light on why some individuals navigate complex ecological affordances more successfully than others.
5.2. Other environmental dimensions across life stages
The proposed framework can be extended to other environmental dimensions tied to recurrent adaptive challenges. For instance, an environment of high pathogen prevalence poses significant threats to individuals at all life stages. However, the risk of encountering pathogens, the types of pathogens prevalent, the severity and fatality of pathogen infections, and the trade-offs of avoidance behaviors varies across life stages. For example, mortality from infection is highest in infancy, declines through childhood, and rises in late adulthood (GBD 2013 Mortality and Causes of Death Collaborators, 2015; Glynn & Moss, Reference Glynn and Moss2020), and such differences likely lead to variations in pathogen defenses and management strategies across life stages.
Furthermore, one might plausibly extend this framework beyond those environmental dimensions long recurring ancestrally to those more temporally novel. For example, although toxic waste dumps are a modern phenomenon with no direct evolutionary precedent, humans may leverage and repurpose existing disgust mechanisms—which evolved to elicit aversions to foul smells, unusual textures, and visually unappealing substances—to recognize and manage contamination risks. Although evolved mechanisms alone are likely insufficient for addressing the complex risks associated with novel environmental features, they may provide a foundation for adaptive responses to novel threats that resemble ancestral challenges.
5.3. Other life stages for shaping ecological affordances
This framework can be expanded to address (a) transitions between stages, (b) overlaps among multiple stages, and (c) life stages extending beyond parenting. Each area presents unique opportunities for further exploration.
First, adolescence, bridging childhood and mating stages, presents unique life stage challenges. Although physically capable of mating and independence, adolescents often delay these milestones to accumulate resources and skills (Natterson-Horowitz & Bowers, Reference Natterson-Horowitz and Bowers2020). This stage is characterized by continuing to seek parental investment (childhood stage goals) while beginning to explore independence and mating opportunities (mating stage goals). To navigate these life stage challenges, the calibration of pubertal timing in response to ecological affordances (Ellis, Reference Ellis2004) may play a role in managing the transition.
Second, stages can overlap, as when pregnant women may lack secure partner investment or when parents of young children are separated or divorced. Such circumstances often involve balancing parenting responsibilities with the pursuit of new mating opportunities, necessitating complex trade-offs (Beall & Schaller, Reference Beall and Schaller2017). Individuals in this overlapping stage likely face distinct ecological affordances compared to those focused on a single goal, underscoring the need for flexible adaptive strategies.
Third, from middle age, adults experience significant life transitions in marriages, careers, parenthood, and grandparenthood (Infurna et al., Reference Infurna, Gerstorf and Lachman2020). Humans are unique among primates in having a prolonged post-reproductive lifespan. This suggests an evolutionary adaptation whereby grandparents, especially grandmothers, enhance offspring success by providing care and resources to grandchildren (Hawkes, Reference Hawkes2004; Hawkes et al., Reference Hawkes, O’Connell, Jones, Alvarez and Charnov2002; Lahdenperä et al., Reference Lahdenperä, Lummaa, Helle, Tremblay and Russell2004). In the grandparenting stage, individuals are likely to support grandchild care and development and thus perceive ecological affordances similarly to parents of young children. This cognitive, emotional, and behavioral calibration is likely to be especially pronounced in maternal grandmothers, who tend to invest more in grandchild care (Perry & Daly, Reference Perry and Daly2017; Sear & Mace, Reference Sear and Mace2008; Strassmann & Garrard, Reference Strassmann and Garrard2011).
Exploring ecological affordances beyond the main life stages of childhood, mating, and parenting will likely pave the way for novel research directions that consider the full complexity of human development across the lifespan.
5.4. Lifelong flexibility in managing ecological affordance changes
Research on critical periods has focused on early life years, particularly infancy through preschool, examining how adverse childhood environments shape development through family environment and parenting quality (Gabard-Durnam & McLaughlin, Reference Gabard-Durnam and McLaughlin2020; Hartley & Frankenhuis, Reference Hartley and Frankenhuis2020). However, for long-lived species like humans, early life environments rarely predict future conditions reliably (Lea & Rosebaum, Reference Lea and Rosebaum2020). Given the variability in ecological conditions and affordances across the lifespan, natural selection has likely favored a degree of lifelong flexibility to modify traits and strategies (Del Giudice & Belsky, Reference Del Giudice and Belsky2010; Dufty Jr et al., Reference Dufty, Clobert and Møller2002; Forsman, Reference Forsman2015; Jones, Reference Jones2005; Ratikainen & Kokko, Reference Ratikainen and Kokko2019; Rickard & Lummaa, Reference Rickard and Lummaa2007; Wells, Reference Wells2007).
Evidence supports this perspective, showing that sensitive periods occur across ontogeny—at onset, midway through, and toward its end (Walasek et al., Reference Walasek, Frankenhuis and Panchanathan2022). Adolescence, for example, represents another critical period during which adverse environments moderate the effects of early childhood adversity (Andersen et al., Reference Andersen, Steinberg and Belsky2021). Our framework offers insights into how humans adapt to spatial environmental changes (e.g., geographic relocations), temporal environmental changes (e.g., pandemics), and shifting ecological affordances across life stages without requiring unique mechanisms for each scenario.
Do humans calibrate the onset, termination, and/or lengths of life stages at biological, psychological, behavioral, and societal levels in response to ecological affordances? For instance, environments with low kin prevalence may promote earlier sexual maturation and earlier transitions to independence. Research shows that girls with absent fathers experience accelerated pubertal development and earlier sexual activity (Coley et al., Reference Coley, Votruba-Drzal and Schindler2009; Draper & Harpending, Reference Draper and Harpending1982; Ellis, Reference Ellis2004; Ellis & Essex, Reference Ellis and Essex2007). Conversely, in environments with high kin prevalence, puberty onset and pursuit of independence may be delayed to avoid inbreeding while maximizing support from kin.
Important questions emerge about lifelong flexibility: (a) What are the critical sensitive periods for lifelong flexibility? (b) What are the implications of having multiple sensitive periods throughout life? (c) What are the costs and benefits associated with the ability to remain flexible across the lifespan? (d) What happens to adaptations once individuals surpass the life stages for which they were optimal? Do they linger with reduced accessibility and diminished function, or are they completely discarded and abandoned, similar to how sea squirts consume their brains upon settling permanently (Cosmides & Tooby, Reference Cosmides and Tooby1997; Glenberg et al., Reference Glenberg, Jaworski, Rischal and Levin2007)? Understanding these dynamics will enhance our understanding of how humans optimize resources in response to changing ecological affordances across the lifespan.
6. Conclusion
The proposed framework, integrating affordance-management and life-history perspectives, offers a unique theoretical approach to explore (a) how ecological affordances change as people’s prioritized goals shift across life-history stages, (b) how these emergent ecological affordances facilitate or hinder the achievement of stage-specific developmental goals, and (c) how people manage their ecological affordances in pursuit of their prioritized goals across life stages.
By bridging social, developmental, and cognitive psychology with behavioral ecology and evolutionary biology, this framework opens novel theoretical and empirical avenues for a comprehensive view of human-environment interactions. First, it challenges the assumption that environmental effects are static, revealing instead how environmental features gain or lose psychological salience and functional relevance as developmental priorities shift. Second, it generates testable hypotheses about psychological and behavioral patterns across the lifespan, enabling systematic and holistic investigation. Third, it highlights the potential for lifelong flexibility in ecological navigation.
We hope the current framework sets the stage for investigating the complexities of ecological affordances in diverse environments and cultures, advancing our understanding of the dynamic interplay between human development and ecological navigation.
Acknowledgements
We thank Oliver Sng, Douglas T. Kenrick, and Frank J. Infurna for their valuable comments on earlier versions of this article.
Financial support
The writing of this manuscript was supported by funds granted to Steven L. Neuberg by the Arizona State University Foundation for a New American University.
Competing interests
The authors declared no potential conflict of interest with respect to the research, authorship, and/or publication of this article.
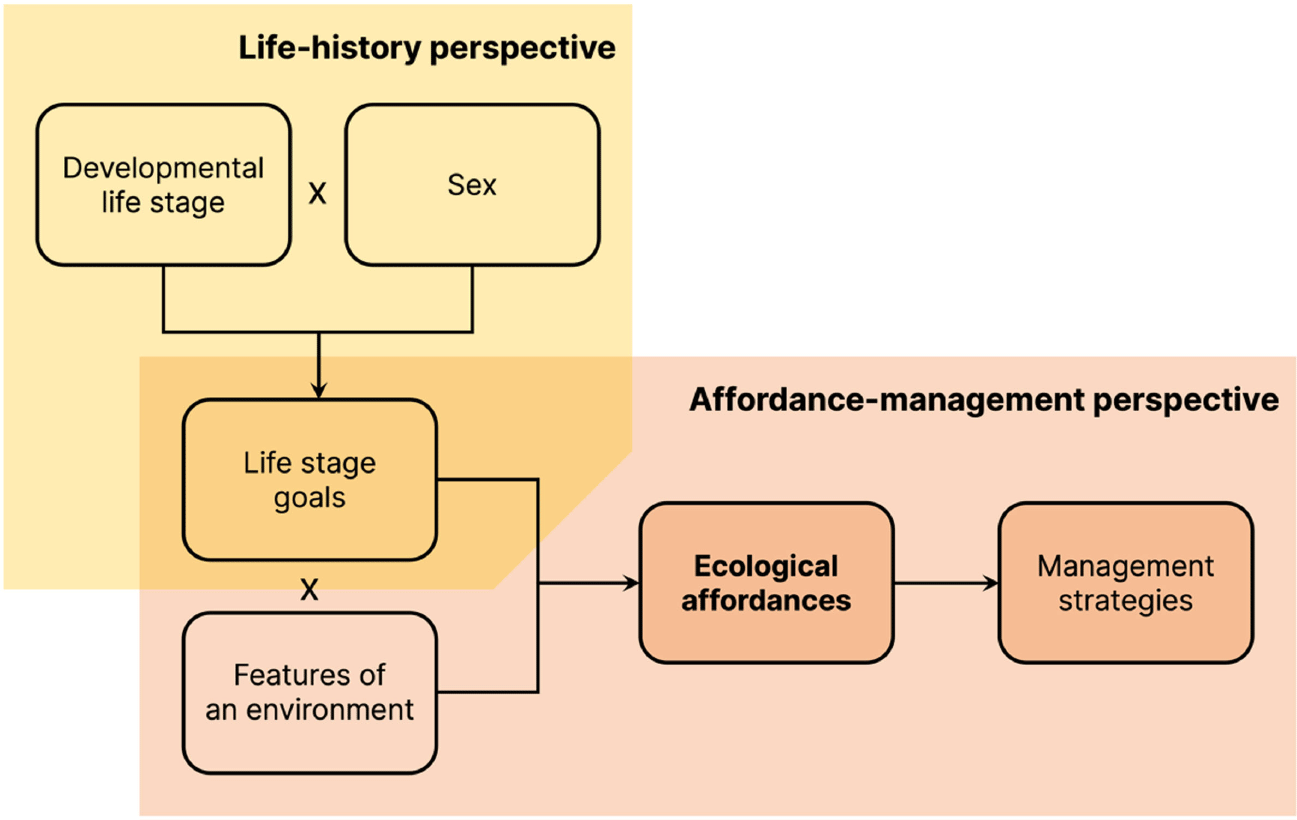
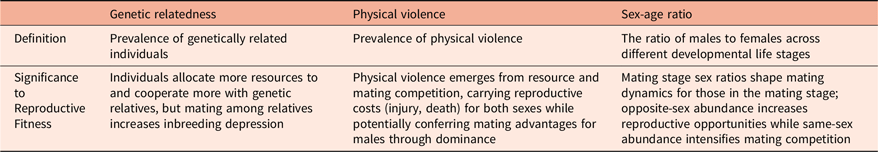
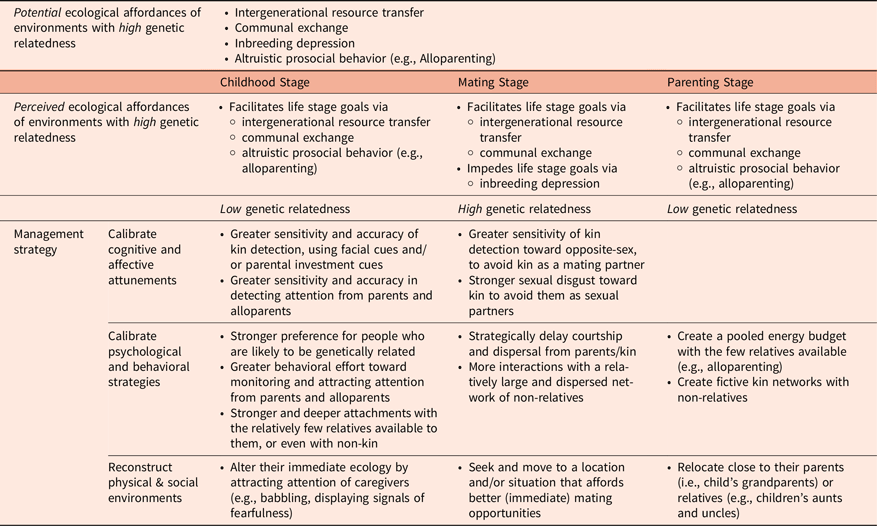
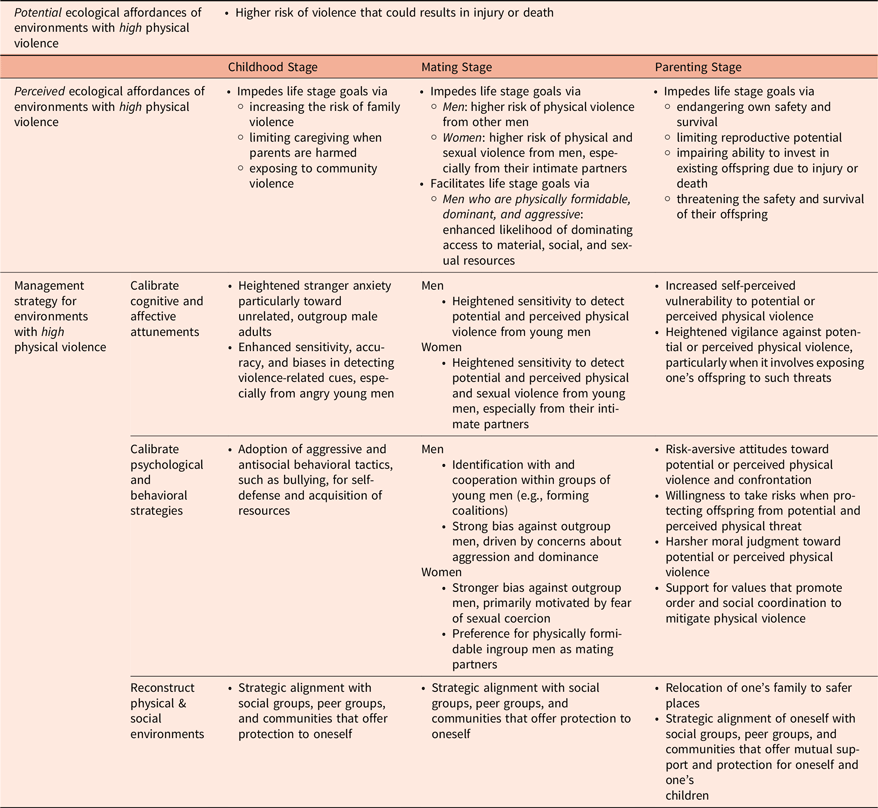
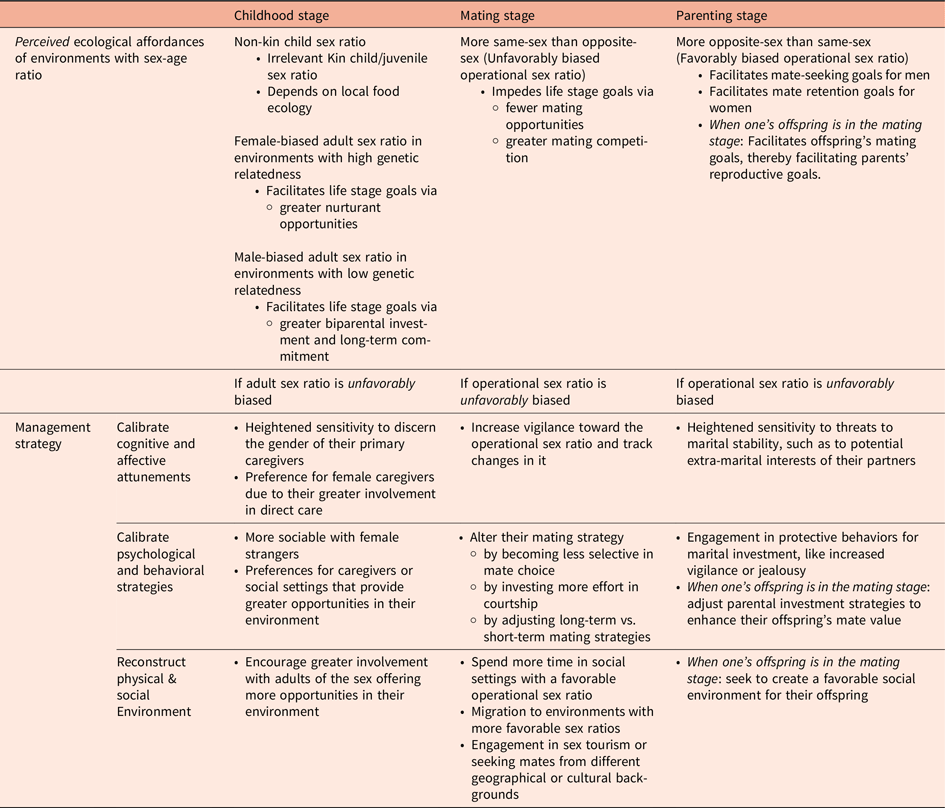

Target article
Ecological affordances across life stages: An affordance-management framework
Related commentaries (29)
A constraint-based approach to ecological affordances
A contingency management framework of ecological affordances across the lifespan
A psychology of ecology or a psychology of groups?
Affordance management’s cultural-ecological extensions outside of the West
Affordances and development: Distinguishing ecological and standard evolutionary psychology approaches
Affordances and life history complexity
Affordances are for life (and not just for maximizing reproductive fitness)
Affordances in social ecologies: A biological markets extension
Affordances, culture, and consciousness
An evolutionary model of human development based on reproduction goals is neither evolutionary nor developmental
Digital environments reshaping children’s ecological affordance management: Hybrid ecologies, parallel strategies, and environmental shaping
Distinguishing regulatory variables and ecological affordances: Prioritizing goals versus implementing action
Ecological affordances across life stages and evolutionary mismatch: hormonal contraceptives as an illustrative example
Ecological affordances as stages for trait expression
Ecological affordances change with the seasons
Extending affordance management to late adulthood: a critical gap target
Extending ecological affordance theory to late adulthood
From life stages to micro-events: Ecological psychology and affordance management
From screens to green: Digital and natural environments as ecological affordances across childhood, mating, and parenting stages
Managing affordances in a complex world depends on social and cultural factors
Neuroplasticity in changing ecologies
Personality adaptively mediates ecological affordances and behavior across the lifespan
Rethinking cognitive recalibration: Integrating AI into the management of affordances across the life-stages
The mind’s compass: semantic control as the guiding mechanism of affordance management
The missing ecology in affordance management
Tracking affordances requires a sophisticated model of life stages, phases, and transitions
Two ecological approaches in social psychology
Violence is not a monolith: Ecologies of threat and their psychological signatures
What do we perceive when we think about perception?
Author response
Ecological affordances across the lifespan: Refinements and Extensions through interdisciplinary perspectives