Refine search
Actions for selected content:
10186 results in Genetics
Small, inherited differences in blood glucose levels in mice
-
- Journal:
- Genetical Research / Volume 14 / Issue 1 / August 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-7
-
- Article
-
- You have access
- Export citation
Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 36 / Issue 3 / December 1980
- Published online by Cambridge University Press:
- 14 April 2009, pp. 339-341
-
- Article
-
- You have access
- Export citation
Genetic locus determining resistance to phage BF23 and colicins E1, E2 and E3 in Escherichia coli*
-
- Journal:
- Genetical Research / Volume 19 / Issue 3 / June 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 305-312
-
- Article
-
- You have access
- Export citation
The pattern of X-chromosome inactivation in the embryonic and extra-embryonic tissues of post-implantation digynic triploid LT/Sv strain mouse embryos
-
- Journal:
- Genetical Research / Volume 56 / Issue 2-3 / October 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 107-114
-
- Article
-
- You have access
- Export citation
Books Received
-
- Journal:
- Genetical Research / Volume 53 / Issue 2 / April 1989
- Published online by Cambridge University Press:
- 14 April 2009, p. 153
-
- Article
-
- You have access
- Export citation
GRH volume 20 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 20 / Issue 1 / August 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Genetic and environmental control of a continuous trait, Abnormal abdomen, in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 22 / Issue 1 / August 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 37-50
-
- Article
-
- You have access
- Export citation
Nucleolus organizer-suppressed position-effect variegation in Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 57 / Issue 3 / June 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 245-255
-
- Article
-
- You have access
- Export citation
The effects of mutant p-alleles on the reproductive system in mice*
-
- Journal:
- Genetical Research / Volume 23 / Issue 3 / June 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 319-325
-
- Article
-
- You have access
- Export citation
The estimation of genotypic probabilities in an adult population by the analysis of descendants
-
- Journal:
- Genetical Research / Volume 59 / Issue 2 / April 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. 131-137
-
- Article
-
- You have access
- Export citation
Rec-mediated recombinational activity of two adjacent Chi elements in bacteriophage lambda
-
- Journal:
- Genetical Research / Volume 45 / Issue 1 / February 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-8
-
- Article
-
- You have access
- Export citation
The effect of non-random mating within inbred lines on the rate of inbreeding
-
- Journal:
- Genetical Research / Volume 5 / Issue 1 / February 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 164-167
-
- Article
-
- You have access
- Export citation
Genetic variation between mice in their metabolism of coumarin and its derivatives
-
- Journal:
- Genetical Research / Volume 31 / Issue 2 / April 1978
- Published online by Cambridge University Press:
- 14 April 2009, pp. 177-186
-
- Article
-
- You have access
- Export citation
The genetic component in coronary heart disease – a review
-
- Journal:
- Genetical Research / Volume 37 / Issue 1 / February 1981
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-16
-
- Article
-
- You have access
- Export citation
Mapping quantitative trait loci using four-way crosses
-
- Journal:
- Genetical Research / Volume 68 / Issue 2 / October 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 175-181
-
- Article
-
- You have access
- Export citation
The Comparative Method in Evolutionary Biology. By Paul H. Harvey and Mark D. Pagel. Oxford University Press. 1991. 239 pages. Price £12.50. ISBN 0 19 854640 8.
-
- Journal:
- Genetical Research / Volume 62 / Issue 1 / August 1993
- Published online by Cambridge University Press:
- 14 April 2009, pp. 81-82
-
- Article
-
- You have access
- Export citation
GRH volume 55 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 55 / Issue 2 / April 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b2
-
- Article
-
- You have access
- Export citation
Abstracts of papers presented at the Mammalian Genetics Group meeting held in the Linnean Society Rooms, Piccadilly, London, on 28 and 29 November 1985
-
- Journal:
- Genetical Research / Volume 47 / Issue 3 / June 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 217-223
-
- Article
-
- You have access
- Export citation
‘Nude’, a new hairless gene with pleiotropic effects in the mouse
-
- Journal:
- Genetical Research / Volume 8 / Issue 3 / December 1966
- Published online by Cambridge University Press:
- 14 April 2009, pp. 295-309
-
- Article
-
- You have access
- Export citation
-
1. Nude is a new recessive gene causing hairlessness in the mouse. It is linked to rex and trembler in linkage group VII. The order of the three loci and the recombination frequencies are as follows:
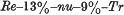
2. In addition to hairlessness the new gene causes reduced body growth rate, very low fertility and a liver disease causing death. Nude mice may be classified at birth by the absence of vibrissae.
3. The hairlessness is due to abnormal keratinization of hair in the follicles. The skin histology resembles that of naked mice. The hair follicles were found to be deficient in free sulphydryl groups.
4. The majority of nude mice die of general body weakness within 2 weeks of birth. The survivors grow slowly and may live for a considerable period. But all nude mice eventually die, usually between 3 and 14 weeks of age.
5. The livers of dead or moribund nude mice are covered with lesions and scars. The defect has been traced histologically to its initial stage, namely, necrosis of small areas of tissue.
6. Attempts to relate the deficiency of sulphydryl groups in the hair follicles to abnormal sulphur metabolism in the liver were unsuccessful.
7. Pseudo-cysts of a parasitic protozoan, Toxoplasma gondii, were identified in the liver and brain of nude mice. In one case the free form of the organism was found.
8. The possible relationship between the liver disease and the pathogenic organism is discussed.
Recombination between two mouse t-haplotypes (tw12tf and tLub-1): segregation of lethal factors relative to centromere and tufted (tf) locus
-
- Journal:
- Genetical Research / Volume 42 / Issue 3 / December 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 335-344
-
- Article
-
- You have access
- Export citation
