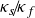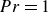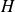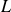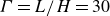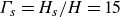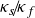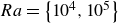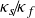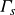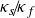Refine search
Actions for selected content:
238502 results in Physics and Astronomy
The higher direct images of locally constant group schemes from the Kummer log flat topology to the classical flat topology
- Part of
-
- Journal:
- Mathematical Proceedings of the Cambridge Philosophical Society / Volume 179 / Issue 3 / November 2025
- Published online by Cambridge University Press:
- 04 August 2025, pp. 667-693
- Print publication:
- November 2025
-
- Article
- Export citation
-
Let S be a fine and saturated (fs) log scheme, and let F be a group scheme over the underlying scheme of S which is étale locally representable by (1) a finite dimensional
 $\mathbb{Q}$-vector space, or (2) a finite rank free abelian group, or (3) a finite abelian group. We give a full description of all the higher direct images of F from the Kummer log flat site to the classical flat site. In particular, we show that: in case (1) the higher direct images of F vanish; and in case (2) the first higher direct image of F vanishes and the nth (
$\mathbb{Q}$-vector space, or (2) a finite rank free abelian group, or (3) a finite abelian group. We give a full description of all the higher direct images of F from the Kummer log flat site to the classical flat site. In particular, we show that: in case (1) the higher direct images of F vanish; and in case (2) the first higher direct image of F vanishes and the nth ( $n\gt 1$) higher direct image of F is isomorphic to the
$n\gt 1$) higher direct image of F is isomorphic to the  $(n-1)$-th higher direct image of
$(n-1)$-th higher direct image of  $F\otimes_{{\mathbb Z}}{\mathbb Q}/{\mathbb Z}$. In the end, we make some computations when the base is a standard henselian log trait or a Dedekind scheme endowed with the log structure associated to a finite set of closed points.
$F\otimes_{{\mathbb Z}}{\mathbb Q}/{\mathbb Z}$. In the end, we make some computations when the base is a standard henselian log trait or a Dedekind scheme endowed with the log structure associated to a finite set of closed points.




The Gardner equation and acoustic solitary waves in plasmas
-
- Journal:
- Journal of Plasma Physics / Volume 91 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 04 August 2025, E117
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Generation of 1-MHz, 64-W, 26-fs green pulses via second-harmonic generation of nonlinearly compressed pulses at 1.03 μm
-
- Journal:
- High Power Laser Science and Engineering / Volume 13 / 2025
- Published online by Cambridge University Press:
- 04 August 2025, e74
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Spectral line identification from a photoionised silicon plasma in emission
-
- Journal:
- Journal of Plasma Physics / Volume 91 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 04 August 2025, E114
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Next-generation X-ray satellite telescopes such as XRISM, NewAthena and Lynx will enable observations of exotic astrophysical sources at unprecedented spectral and spatial resolution. Proper interpretation of these data demands that the accuracy of the models is at least within the uncertainty of the observations. One set of quantities that might not currently meet this requirement is transition energies of various astrophysically relevant ions. Current databases are populated with many untested theoretical calculations. Accurate laboratory benchmarks are required to better understand the coming data. We obtained laboratory spectra of X-ray lines from a silicon plasma at an average spectral resolving power of
 $\sim$7500 with a spherically bent crystal spectrometer on the Z facility at Sandia National Laboratories. Many of the lines in the data are measured here for the first time. We report measurements of 53 transitions originating from the K-shells of He-like to B-like silicon in the energy range between
$\sim$7500 with a spherically bent crystal spectrometer on the Z facility at Sandia National Laboratories. Many of the lines in the data are measured here for the first time. We report measurements of 53 transitions originating from the K-shells of He-like to B-like silicon in the energy range between  $\sim$1795 and 1880 eV (6.6–6.9 Å). The lines were identified by qualitative comparison against a full synthetic spectrum calculated with ATOMIC. The average fractional uncertainty (uncertainty/energy) for all reported lines is
$\sim$1795 and 1880 eV (6.6–6.9 Å). The lines were identified by qualitative comparison against a full synthetic spectrum calculated with ATOMIC. The average fractional uncertainty (uncertainty/energy) for all reported lines is  ${\sim}5.4 \times 10^{-5}$. We compare the measured quantities against transition energies calculated with RATS and FAC as well as those reported in the NIST ASD and XSTAR’s uaDB. Average absolute differences relative to experimentally measured values are 0.20, 0.32, 0.17 and 0.38 eV, respectively. All calculations/databases show good agreement with the experimental values; NIST ASD shows the closest match overall.
${\sim}5.4 \times 10^{-5}$. We compare the measured quantities against transition energies calculated with RATS and FAC as well as those reported in the NIST ASD and XSTAR’s uaDB. Average absolute differences relative to experimentally measured values are 0.20, 0.32, 0.17 and 0.38 eV, respectively. All calculations/databases show good agreement with the experimental values; NIST ASD shows the closest match overall.



Resilience of DNA chains to molecular fracture after PCR heating cycles and implications on PCR reliability – EXPRESSION OF CONCERN
-
- Journal:
- Quarterly Reviews of Biophysics / Volume 58 / 2025
- Published online by Cambridge University Press:
- 04 August 2025, e16
-
- Article
-
- You have access
- HTML
- Export citation
The Velikhov-ionisation instability revisited: a new opportunity for MHD energy conversion?
-
- Journal:
- Journal of Plasma Physics / Volume 91 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 04 August 2025, E115
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Moduli spaces of rational graphically stable curves
- Part of
-
- Journal:
- Mathematical Proceedings of the Cambridge Philosophical Society / Volume 179 / Issue 3 / November 2025
- Published online by Cambridge University Press:
- 04 August 2025, pp. 649-665
- Print publication:
- November 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Reynolds number effect on the flow statistics and turbulent–non-turbulent interface of a planar jet
-
- Journal:
- Journal of Fluid Mechanics / Volume 1016 / 10 August 2025
- Published online by Cambridge University Press:
- 04 August 2025, A37
-
- Article
- Export citation
-
We investigate the influence of the Reynolds number on the spatial development of an incompressible planar jet. The study relies on direct numerical simulations (DNS) at inlet Reynolds numbers between
 $500 \leqslant Re \leqslant 13\,500$, being the widest range and the largest values considered so far in DNS. At the lowest
$500 \leqslant Re \leqslant 13\,500$, being the widest range and the largest values considered so far in DNS. At the lowest  $Re$, the flow is transitional and characterised by large quasi-two-dimensional vortices; at the largest
$Re$, the flow is transitional and characterised by large quasi-two-dimensional vortices; at the largest  $Re$, the flow reaches a fully turbulent regime with a well-developed self-similar region. We provide a complete description of the flow, from the instabilities in the laminar near-inlet region, to the self-similar regime in the turbulent far field. At the inlet, the leading destabilisation mode is sinusoidal/asymmetric at low Reynolds number and varicose/symmetric at large Reynolds number, with both modes coexisting at intermediate
$Re$, the flow reaches a fully turbulent regime with a well-developed self-similar region. We provide a complete description of the flow, from the instabilities in the laminar near-inlet region, to the self-similar regime in the turbulent far field. At the inlet, the leading destabilisation mode is sinusoidal/asymmetric at low Reynolds number and varicose/symmetric at large Reynolds number, with both modes coexisting at intermediate  $Re$. In the far field, the mean and fluctuating statistics converge to self-similar profiles only for
$Re$. In the far field, the mean and fluctuating statistics converge to self-similar profiles only for  $Re\geqslant 4500$; the flow anisotropy, the budget of the Reynolds stresses and the energy spectra are addressed. The spreading of the jet is quantified via the turbulent–non-turbulent interface (TNTI). We find that the thickness of the turbulent region, and the shape and fractal dimension of the TNTI become
$Re\geqslant 4500$; the flow anisotropy, the budget of the Reynolds stresses and the energy spectra are addressed. The spreading of the jet is quantified via the turbulent–non-turbulent interface (TNTI). We find that the thickness of the turbulent region, and the shape and fractal dimension of the TNTI become  $Re$-independent for
$Re$-independent for  $Re \geqslant 4500$. Comparisons with previous numerical and experimental works are provided whenever available.
$Re \geqslant 4500$. Comparisons with previous numerical and experimental works are provided whenever available.








Dynamics of a liquid film flowing down a granular chain
-
- Journal:
- Journal of Fluid Mechanics / Volume 1016 / 10 August 2025
- Published online by Cambridge University Press:
- 04 August 2025, A36
-
- Article
- Export citation
On the quasi-transitivity degree of branch groups
- Part of
-
- Journal:
- Mathematical Proceedings of the Cambridge Philosophical Society / Volume 179 / Issue 3 / November 2025
- Published online by Cambridge University Press:
- 04 August 2025, pp. 695-715
- Print publication:
- November 2025
-
- Article
- Export citation
-
An action of a group G on a set X is said to be quasi-n-transitive if the diagonal action of G on
 $X^n$ has only finitely many orbits. We show that branch groups, a special class of groups of automorphisms of rooted trees, cannot act quasi-2-transitively on infinite sets.
$X^n$ has only finitely many orbits. We show that branch groups, a special class of groups of automorphisms of rooted trees, cannot act quasi-2-transitively on infinite sets.


Enhancement of heat transfer using Faraday instability
-
- Journal:
- Journal of Fluid Mechanics / Volume 1016 / 10 August 2025
- Published online by Cambridge University Press:
- 04 August 2025, A35
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Quantum calculation for two-stream instability and advection test of Vlasov–Maxwell equations: numerical evaluation of Hamiltonian simulation
-
- Journal:
- Journal of Plasma Physics / Volume 91 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 04 August 2025, E116
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The Vlasov–Maxwell equations provide kinetic simulations of collisionless plasmas, but numerically solving them on classical computers is often impractical. This is due to the computational resource constraints imposed by the time evolution in the six-dimensional phase space, which requires broad spatial and temporal scales. The novelty of this study is to implement a quantum–classical hybrid Vlasov–Maxwell solver and the rigorous numerical scheme evaluation by numerical simulations. Specifically, the Vlasov solver implements the Hamiltonian simulation based on quantum singular value transformation, coupled with a classical Maxwell solver. We perform numerical simulation of a one-dimensional advection test and a one-spatial-dimension, one-velocity-dimension two-stream instability test on the Qiskit-Aer-GPU quantum circuit emulator with an A100 GPU. The computational complexity of our quantum algorithm can potentially be reduced from the classical
 $\mathcal{O}(N^6T^2/\epsilon )$ to
$\mathcal{O}(N^6T^2/\epsilon )$ to  $\mathcal{O}\left (\text{poly}(\log {N})\left (NT+T\log \left (T/\epsilon \right )\right )\right )$ for the
$\mathcal{O}\left (\text{poly}(\log {N})\left (NT+T\log \left (T/\epsilon \right )\right )\right )$ for the  $N$ grid system, simulation time
$N$ grid system, simulation time  $T$ and error tolerance
$T$ and error tolerance  $\epsilon$ in the limit where the number of queries is large enough and the error is small enough. Furthermore, the numerical analysis reveals that our quantum algorithm is robust under larger time steps compared with classical algorithms with the constraint of Courant–Friedrichs–Lewy condition.
$\epsilon$ in the limit where the number of queries is large enough and the error is small enough. Furthermore, the numerical analysis reveals that our quantum algorithm is robust under larger time steps compared with classical algorithms with the constraint of Courant–Friedrichs–Lewy condition.





Biosignature preservation in FaRLiP cyanobacteria after prolonged desiccation and its relevance to space missions searching for life
-
- Journal:
- International Journal of Astrobiology / Volume 24 / 2025
- Published online by Cambridge University Press:
- 04 August 2025, e14
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Pulse duration stability in the SG-II picosecond-petawatt laser system
-
- Journal:
- High Power Laser Science and Engineering / Volume 13 / 2025
- Published online by Cambridge University Press:
- 04 August 2025, e75
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Near-axis measures of quasi-isodynamic configurations
-
- Journal:
- Journal of Plasma Physics / Volume 91 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 01 August 2025, E112
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We present a number of measures and techniques to characterise and effectively construct quasi-isodynamic stellarators within the near-axis framework, without the need to resort to the computation of global equilibria. These include measures of the reliability of the model (including aspect-ratio limits and the appearance of ripple wells), quantification of omnigeneity through
 $\epsilon _{\mathrm{eff}}$, measure and construction of MHD-stabilised fields, and the sensitivity of the field to the pressure gradient. The paper presents, discusses and gives examples of all of these, for which expansions to second order are crucial. This opens the door to the exploration of how key underlying choices of the field design govern the interaction of desired properties (‘trade-offs’) and provides a practical toolkit to perform efficient optimisation directly within the space of near-axis quasi-isodynamic configurations.
$\epsilon _{\mathrm{eff}}$, measure and construction of MHD-stabilised fields, and the sensitivity of the field to the pressure gradient. The paper presents, discusses and gives examples of all of these, for which expansions to second order are crucial. This opens the door to the exploration of how key underlying choices of the field design govern the interaction of desired properties (‘trade-offs’) and provides a practical toolkit to perform efficient optimisation directly within the space of near-axis quasi-isodynamic configurations.

Effects of settling on inertial particle slip velocity statistics in wall-bounded flows
-
- Journal:
- Journal of Fluid Mechanics / Volume 1015 / 25 July 2025
- Published online by Cambridge University Press:
- 01 August 2025, A21
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Developing reduced-order models for the transport of solid particles in turbulence typically requires a statistical description of the particle–turbulence interactions. In this work, we utilize a statistical framework to derive continuum equations for the moments of the slip velocity of inertial, settling Lagrangian particles in a turbulent boundary layer. Using coupled Eulerian–Lagrangian direct numerical simulations, we then identify the dominant mechanisms controlling the slip velocity variance, and find that for a range of Stokes number
 ${S{\kern-0.5pt}t}^+$, Settling number
${S{\kern-0.5pt}t}^+$, Settling number  ${S{\kern-0.5pt}v}^+$ and Reynolds number
${S{\kern-0.5pt}v}^+$ and Reynolds number  $\textit{Re}_\tau$ (based on frictional scales),the slip variance is primarily controlled by local differences between the ‘seen’ variance and the particle velocity variance, while terms appearing due to the inhomogeneity of the turbulence are subleading until
$\textit{Re}_\tau$ (based on frictional scales),the slip variance is primarily controlled by local differences between the ‘seen’ variance and the particle velocity variance, while terms appearing due to the inhomogeneity of the turbulence are subleading until  ${S{\kern-0.5pt}v}^+$ becomes large. We also consider several comparative metrics to assess the relative magnitudes of the fluctuating slip velocity and the mean slip velocity, and we find that the vertical mean slip increases rapidly with
${S{\kern-0.5pt}v}^+$ becomes large. We also consider several comparative metrics to assess the relative magnitudes of the fluctuating slip velocity and the mean slip velocity, and we find that the vertical mean slip increases rapidly with  ${S{\kern-0.5pt}v}^+$, rendering the variance relatively small – an effect found to be most substantial for
${S{\kern-0.5pt}v}^+$, rendering the variance relatively small – an effect found to be most substantial for  ${S{\kern-0.5pt}v}^+\gt 1$. Finally, we compare the results with a model of the acceleration variance (Berk & Coletti 2021 J. Fluid Mech. 917, A47) based the concept of a response function described in Csanady (1963 J. Atmos. Sci. 20, 201–208), highlighting the role of the crossing trajectories mechanism. We find that while there is good agreement for low
${S{\kern-0.5pt}v}^+\gt 1$. Finally, we compare the results with a model of the acceleration variance (Berk & Coletti 2021 J. Fluid Mech. 917, A47) based the concept of a response function described in Csanady (1963 J. Atmos. Sci. 20, 201–208), highlighting the role of the crossing trajectories mechanism. We find that while there is good agreement for low  ${S{\kern-0.5pt}v}^+$, systematic errors remain, possibly due to implicit non-local effects arising from rapid particle settling and inhomogeneous turbulence. We conclude with a discussion of the implications of this work for modelling the transport of coarse dust grains in the atmospheric surface layer.
${S{\kern-0.5pt}v}^+$, systematic errors remain, possibly due to implicit non-local effects arising from rapid particle settling and inhomogeneous turbulence. We conclude with a discussion of the implications of this work for modelling the transport of coarse dust grains in the atmospheric surface layer.







Inverse cascade from helical and non-helical decaying columnar magnetic fields
-
- Journal:
- Journal of Plasma Physics / Volume 91 / Issue 4 / August 2025
- Published online by Cambridge University Press:
- 01 August 2025, E113
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Powerful lasers may be used in the future to produce magnetic fields that would allow us to study turbulent magnetohydrodynamic inverse cascade behaviour. This has so far only been seen in numerical simulations. In the laboratory, however, the produced fields may be highly anisotropic. Here, we present corresponding simulations to show that, during the turbulent decay, such a magnetic field undergoes spontaneous isotropisation. As a consequence, we find the decay dynamics to be similar to that in isotropic turbulence. We also find that an initially pointwise non-helical magnetic field is unstable and develops magnetic helicity fluctuations that can be quantified by the Hosking integral. It is a conserved quantity that characterises magnetic helicity fluctuations and governs the turbulent decay when the mean magnetic helicity vanishes. As in earlier work, the ratio of the magnetic decay time to the Alfvén time is found to be approximately
 $50$ in the helical and non-helical cases. At intermediate times, the ratio can even reach a hundred. This ratio determines the endpoints of cosmological magnetic field evolution.
$50$ in the helical and non-helical cases. At intermediate times, the ratio can even reach a hundred. This ratio determines the endpoints of cosmological magnetic field evolution.


Explicit formulas for forced convection in a shrouded longitudinal-fin heat sink with clearance
-
- Journal:
- Journal of Fluid Mechanics / Volume 1016 / 10 August 2025
- Published online by Cambridge University Press:
- 01 August 2025, A32
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Fungus Penicillium rubens isolated from the International Space Station (ISS) shows faster colony growth and better spore resistance to cosmic radiation than type strain
-
- Journal:
- International Journal of Astrobiology / Volume 24 / 2025
- Published online by Cambridge University Press:
- 01 August 2025, e13
-
- Article
-
- You have access
- Open access
- HTML
- Export citation

Effects of conjugate heat transfer on large-scale flow structures in convection
-
- Journal:
- Journal of Fluid Mechanics / Volume 1016 / 10 August 2025
- Published online by Cambridge University Press:
- 01 August 2025, A30
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The constant temperature and constant heat flux thermal boundary conditions, both developing distinct flow patterns, represent limiting cases of ideally conducting and insulating plates in Rayleigh–Bénard convection flows, respectively. This study bridges the gap in between, using a conjugate heat transfer (CHT) set-up and studying finite thermal diffusivity ratios
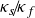 $\kappa _s \! / \! \kappa _f$ to better represent real-life conditions in experiments. A three-dimensional Rayleigh–Bénard convection configuration including two fluid-confining plates is studied via direct numerical simulations given a Prandtl number
$\kappa _s \! / \! \kappa _f$ to better represent real-life conditions in experiments. A three-dimensional Rayleigh–Bénard convection configuration including two fluid-confining plates is studied via direct numerical simulations given a Prandtl number 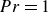 ${Pr}=1$. The fluid layer of height
${Pr}=1$. The fluid layer of height 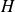 $H$ and horizontal extension
$H$ and horizontal extension 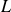 $L$ obeys no-slip boundary conditions at the two solid–fluid interfaces and an aspect ratio of
$L$ obeys no-slip boundary conditions at the two solid–fluid interfaces and an aspect ratio of 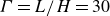 ${\Gamma }=L/H=30$ while the relative thickness of each plate is
${\Gamma }=L/H=30$ while the relative thickness of each plate is 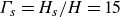 ${\Gamma _s}=H_s/H=15$. The entire domain is laterally periodic. Here, different
${\Gamma _s}=H_s/H=15$. The entire domain is laterally periodic. Here, different 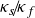 $\kappa _s \! / \! \kappa _f$ are investigated for moderate Rayleigh numbers
$\kappa _s \! / \! \kappa _f$ are investigated for moderate Rayleigh numbers 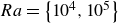 $Ra=\left \{ 10^4, 10^5 \right \}$. We observe a gradual shift of the size of the characteristic flow patterns and their induced heat and mass transfer as
$Ra=\left \{ 10^4, 10^5 \right \}$. We observe a gradual shift of the size of the characteristic flow patterns and their induced heat and mass transfer as 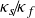 $\kappa _s \! / \! \kappa _f$ is varied, suggesting a relation between the recently studied turbulent superstructures and supergranules for constant temperature and constant heat flux boundary conditions, respectively. Performing a linear stability analysis for this CHT configuration confirms these observations theoretically while extending previous studies by investigating the impact of a varying solid plate thickness
$\kappa _s \! / \! \kappa _f$ is varied, suggesting a relation between the recently studied turbulent superstructures and supergranules for constant temperature and constant heat flux boundary conditions, respectively. Performing a linear stability analysis for this CHT configuration confirms these observations theoretically while extending previous studies by investigating the impact of a varying solid plate thickness 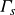 $\Gamma _s$. Moreover, we study the impact of
$\Gamma _s$. Moreover, we study the impact of 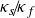 $\kappa _s \! / \! \kappa _f$ on both the thermal and viscous boundary layers. Given the prevalence of finite
$\kappa _s \! / \! \kappa _f$ on both the thermal and viscous boundary layers. Given the prevalence of finite  $\kappa _s \! / \! \kappa _f$ in nature, this work is a starting point to extend our understanding of pattern formation in geo- and astrophysical convection flows.
$\kappa _s \! / \! \kappa _f$ in nature, this work is a starting point to extend our understanding of pattern formation in geo- and astrophysical convection flows.