Refine search
Actions for selected content:
238160 results in Physics and Astronomy
On the role of viscoelasticity in mucociliary clearance: a hydrodynamic continuum approach
-
- Journal:
- Journal of Fluid Mechanics / Volume 971 / 25 September 2023
- Published online by Cambridge University Press:
- 21 September 2023, A33
-
- Article
- Export citation

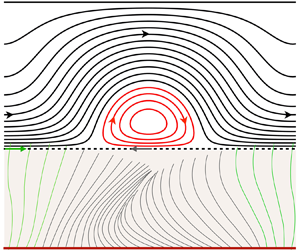
Mode selection in concentric jets: the steady–steady 1 : 2 resonant mode interaction with O(2) symmetry
-
- Journal:
- Journal of Fluid Mechanics / Volume 971 / 25 September 2023
- Published online by Cambridge University Press:
- 21 September 2023, A30
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The linear and nonlinear stability of two concentric jets separated by a duct wall is analysed by means of global linear stability and weakly nonlinear analysis. Three governing parameters are considered, the Reynolds number based on the inner jet, the inner-to-outer jet velocity ratio (
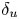 $\delta _u$) and the length of the duct wall (
$\delta _u$) and the length of the duct wall (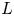 $L$) separating the jet streams. Global linear stability analysis demonstrates the existence of unsteady modes of inherent convective nature, and symmetry-breaking modes that lead to a new non-axisymmetric steady state with a single or double helix. Additionally, we highlight the existence of multiple steady states, as a result of a series of saddle-node bifurcations and its connection to the changes in the topology of the flow. The neutral lines of stability have been computed for inner-to-outer velocity ratios within the range
$L$) separating the jet streams. Global linear stability analysis demonstrates the existence of unsteady modes of inherent convective nature, and symmetry-breaking modes that lead to a new non-axisymmetric steady state with a single or double helix. Additionally, we highlight the existence of multiple steady states, as a result of a series of saddle-node bifurcations and its connection to the changes in the topology of the flow. The neutral lines of stability have been computed for inner-to-outer velocity ratios within the range 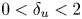 $0 < \delta _u < 2$ and duct wall distances in the interval
$0 < \delta _u < 2$ and duct wall distances in the interval 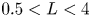 $0.5 < L < 4$. They reveal the existence of hysteresis, and mode switching between two symmetry-breaking modes with azimuthal wavenumbers
$0.5 < L < 4$. They reveal the existence of hysteresis, and mode switching between two symmetry-breaking modes with azimuthal wavenumbers 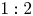 $1:2$. Finally, the mode interaction is analysed, highlighting the presence of travelling waves emerging from the resonant interaction of the two steady states, and the existence of robust heteroclinic cycles that are asymptotically stable.
$1:2$. Finally, the mode interaction is analysed, highlighting the presence of travelling waves emerging from the resonant interaction of the two steady states, and the existence of robust heteroclinic cycles that are asymptotically stable.
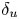
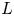
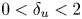
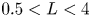
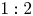
Copyright page
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp iv-iv
-
- Chapter
- Export citation

Spreading of a thin droplet on a soft substrate
-
- Journal:
- Journal of Fluid Mechanics / Volume 971 / 25 September 2023
- Published online by Cambridge University Press:
- 21 September 2023, A32
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Chapter 2 - Solutions of the Schrödinger Equation in One Dimension
-
- Book:
- Quantum Mechanics with Concept Maps
- Published online:
- 07 September 2023
- Print publication:
- 21 September 2023, pp 35-66
-
- Chapter
- Export citation
7 - Possible Conditions on an Exoplanet
- from Part II - The Universe We Live In
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 87-99
-
- Chapter
- Export citation
Preface
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp xvii-xix
-
- Chapter
- Export citation
Bibliography
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 632-660
-
- Chapter
- Export citation
Part IV - The Runaway Brain
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 249-326
-
- Chapter
- Export citation
12 - How Does the Whole Work?
- from Part III - Our Story on Earth
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 164-182
-
- Chapter
- Export citation
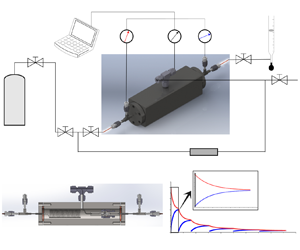
Inertial solution for high-pressure-difference pulse-decay measurement through microporous media
-
- Journal:
- Journal of Fluid Mechanics / Volume 971 / 25 September 2023
- Published online by Cambridge University Press:
- 21 September 2023, R1
-
- Article
- Export citation
-
We present a theoretical asymptotic solution for high-speed transient flow through microporous media in this work by addressing the inertia effect in the high-pressure-difference pulse-decay process. The capillaric model is adopted, in which a bundle of straight circular tubes with a high length–radius ratio is used to represent the internal flow paths of microporous media so that the flow is described by a simplified incompressible Navier–Stokes equation based on the mean density, capturing the major characteristics of mass flow rate. By order-of-magnitude analysis and asymptotic perturbation, the inertial solution with its dimensionless criterion for the high-pressure-difference pulse-decay process is derived. To be compared with experimental data, the theoretical solution involves all three related effects, including the inertia effect, the slippage effect and the compressibility effect. A self-built experimental platform is therefore established to measure the permeability of microporous media by both pulse-decay and steady-state methods to validate the theoretical solution. The results indicate that the relative difference between two methods is less than 30 % even for permeability at as low as
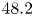 $48.2$ nD
$48.2$ nD 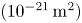 $(10^{-21}\,{\rm m}^2)$, and the present theoretical solution can accurately capture the inertia effect in the high-pressure-difference pulse-decay process, which significantly accelerates the measurements for ultra-low-permeability samples.
$(10^{-21}\,{\rm m}^2)$, and the present theoretical solution can accurately capture the inertia effect in the high-pressure-difference pulse-decay process, which significantly accelerates the measurements for ultra-low-permeability samples.
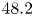
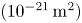
Appendices
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 587-592
-
- Chapter
- Export citation
Chapter 1 - Fundamentals
-
- Book:
- Quantum Mechanics with Concept Maps
- Published online:
- 07 September 2023
- Print publication:
- 21 September 2023, pp 1-34
-
- Chapter
- Export citation
Chapter 6 - Mathematical Background
-
- Book:
- Quantum Mechanics with Concept Maps
- Published online:
- 07 September 2023
- Print publication:
- 21 September 2023, pp 135-169
-
- Chapter
- Export citation
28 - What Might Exolife Be Like?
- from Part VI - Life and Language, Here and Beyond
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 448-471
-
- Chapter
- Export citation
References
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 626-631
-
- Chapter
- Export citation
2 - Looking beyond Earth
- from Part I - Introduction
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 15-30
-
- Chapter
- Export citation
34 - Looking Forward: The Basic Questions Again
- from Part VI - Life and Language, Here and Beyond
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 574-580
-
- Chapter
- Export citation
8 - How and Where to Look for Exolife
- from Part II - The Universe We Live In
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 100-127
-
- Chapter
- Export citation
24 - Language and the Brain
- from Part V - Language, Our Greatest Gift
-
- Book:
- Life and Language Beyond Earth
- Published online:
- 16 September 2023
- Print publication:
- 21 September 2023, pp 399-409
-
- Chapter
- Export citation
